การบ้าน. รองเท้าแตะ Infusoria
เมื่อเปรียบเทียบกับโปรโตซัวกลุ่มอื่น ciliates มีโครงสร้างที่ซับซ้อนที่สุด ซึ่งสัมพันธ์กับความหลากหลายและความซับซ้อนของหน้าที่ของพวกมัน
ชื่อ "รองเท้าแตะ infusoria" มาจากไหน? คุณจะไม่แปลกใจเลยถ้าคุณดูกล้องจุลทรรศน์ที่มี ciliate ที่มีชีวิต หรือแม้แต่รูปของมัน (รูปที่ 85)
อันที่จริง รูปร่างของ ciliate นี้คล้ายกับรองเท้าสตรีที่สง่างาม
รองเท้า Infusoria มีการเคลื่อนไหวค่อนข้างเร็วอย่างต่อเนื่อง ความเร็ว (ที่อุณหภูมิห้อง) ประมาณ 2.0-2.5 มม./วินาที นั่นเป็นความเร็วมากสำหรับสัตว์ตัวเล็ก ๆ ตัวนี้! ท้ายที่สุด นี่หมายความว่าในไม่กี่วินาทีรองเท้าจะวิ่งระยะทางเกินความยาวของลำตัว 10-15 เท่า วิถีของรองเท้าค่อนข้างซับซ้อน เธอเคลื่อนส่วนหน้าตรงไปข้างหน้า
รองเท้า INFUSORIUM (PARAMECIUM CAUDATUM)
เพื่อทำความคุ้นเคยกับโครงสร้างและวิถีชีวิตของสิ่งมีชีวิตเซลล์เดียวที่น่าสนใจเหล่านี้ ก่อนอื่นเรามาดูตัวอย่างลักษณะเฉพาะ ให้เรานำ ciliates รองเท้า (สายพันธุ์ Paramecium) ซึ่งแพร่หลายในอ่างเก็บน้ำน้ำจืดตื้น ciliates เหล่านี้ง่ายมากที่จะผสมพันธุ์ในตู้ปลาขนาดเล็กถ้าคุณเติมหญ้าแห้งในบ่อด้วยหญ้าแห้งธรรมดา ในทิงเจอร์ดังกล่าว โปรโตซัวหลายประเภทพัฒนา และ ciliates รองเท้ามักจะพัฒนา ด้วยความช่วยเหลือของกล้องจุลทรรศน์เพื่อการศึกษาทั่วไป คุณสามารถเห็นสิ่งมากมายที่จะกล่าวถึงต่อไป
ในบรรดาที่ง่ายที่สุด รองเท้า ciliatesเป็นสิ่งมีชีวิตที่ค่อนข้างใหญ่ ความยาวลำตัวประมาณ 1/6-1/3 มม. และหมุนไปทางขวาตามแกนตามยาวของร่างกาย
การเคลื่อนไหวของรองเท้านั้นขึ้นอยู่กับการทำงานของอวัยวะที่คล้ายผมจำนวนมาก - cilia ที่ปกคลุมทั้งตัวของ ciliate จำนวน cilia ในหนึ่ง ciliates รองเท้าคือ 10-15,000!
ซีลีเนียมแต่ละตัวทำการเคลื่อนไหวเหมือนไม้พายบ่อยครั้งมาก - ที่อุณหภูมิห้องสูงถึง 30 ครั้งต่อวินาที ในระหว่างการเป่ากลับ ซีลีเนียมจะอยู่ในตำแหน่งที่เหยียดตรง เมื่อกลับสู่ตำแหน่งเดิม (เมื่อเลื่อนลง) จะเคลื่อนที่ช้าลง 3-5 เท่าและอธิบายรูปครึ่งวงกลม
เมื่อรองเท้าแหวกว่าย จะมีการสรุปการเคลื่อนไหวของขนตาจำนวนมากที่ปกคลุมร่างกายของมัน การทำงานของตาแต่ละตาได้รับการประสานกันทำให้เกิดการสั่นสะเทือนเหมือนคลื่นที่ถูกต้องของทุกตา คลื่นสั่นเริ่มต้นที่ส่วนหน้าของร่างกายและแพร่กระจายไปข้างหลัง ในขณะเดียวกัน คลื่นของการหดตัว 2-3 คลื่นจะเคลื่อนไปตามลำตัวของรองเท้า ดังนั้นเครื่องมือปรับเลนส์ทั้งหมดของ ciliate จึงเป็นเสมือนการทำงานทางสรีรวิทยาทั้งหมดซึ่งเป็นการกระทำของหน่วยโครงสร้างแต่ละหน่วยซึ่ง (cilia) มีการเชื่อมต่ออย่างใกล้ชิด (ประสานงาน) ซึ่งกันและกัน
โครงสร้างของซีเลียมรองเท้าแต่ละอัน ดังที่แสดงโดยการศึกษาด้วยกล้องจุลทรรศน์อิเล็กตรอน มีความซับซ้อนมาก
ทิศทางและความเร็วของการเคลื่อนที่ของรองเท้าไม่คงที่และไม่เปลี่ยนแปลง รองเท้าก็เหมือนกับสิ่งมีชีวิตทั้งหมด (เราได้เห็นสิ่งนี้กับตัวอย่างของอะมีบาแล้ว) ตอบสนองต่อการเปลี่ยนแปลง สภาพแวดล้อมภายนอกเปลี่ยนทิศทางของการเคลื่อนไหว
การเปลี่ยนแปลงทิศทางการเคลื่อนที่ของโปรโตซัวภายใต้อิทธิพลของสิ่งเร้าต่างๆ เรียกว่า แท็กซี่ ใน ciliates มันง่ายที่จะสังเกตแท็กซี่ต่างๆ หากตกหล่นที่รองเท้าลอย ให้วางสารที่ส่งผลเสียต่อรองเท้า (เช่น คริสตัล เกลือแกง) จากนั้นรองเท้าก็ลอยออกไป (ราวกับวิ่งหนี) จากปัจจัยนี้ที่ไม่เอื้ออำนวยต่อพวกเขา (รูปที่ 86)

ก่อนที่เราจะเป็นตัวอย่างของรถแท็กซี่เชิงลบเกี่ยวกับผลกระทบทางเคมี (คีโมแทกซิสเชิงลบ) คุณสามารถสังเกตรองเท้าและเคมีบำบัดเชิงบวก ตัวอย่างเช่น หากหยดน้ำที่ ciliates ว่ายน้ำถูกปกคลุมด้วยกระจกและปล่อยฟองคาร์บอนไดออกไซด์ (CO2) ไว้ใต้มัน ciliates ส่วนใหญ่จะไปที่ฟองนี้และปักหลักอยู่ใน แหวน.
ปรากฏการณ์ของแท็กซี่ปรากฏชัดมากในรองเท้าภายใต้อิทธิพลของกระแสไฟฟ้า หากกระแสไฟฟ้าอ่อนไหลผ่านของเหลวที่รองเท้าลอยอยู่ สามารถสังเกตภาพต่อไปนี้: ciliates ทั้งหมดปรับแนวแกนตามยาวของพวกเขาขนานกับเส้นปัจจุบัน และจากนั้น ราวกับว่าได้รับคำสั่ง ให้เคลื่อนไปทางแคโทด ในพื้นที่ซึ่งก่อตัวเป็นกระจุกหนาแน่น การเคลื่อนที่ของ ciliates ที่กำหนดโดยทิศทางของกระแสไฟฟ้าเรียกว่า galvanotaxis สามารถตรวจจับรถแท็กซี่ต่างๆ ใน ciliates ได้ภายใต้อิทธิพลของปัจจัยแวดล้อมที่หลากหลาย
โครงร่างไซโตพลาสซึมทั้งหมดของ ciliate นั้นแบ่งออกเป็น 2 ชั้นอย่างชัดเจน: ชั้นนอกเบากว่า (ectoplasm) และชั้นในนั้นมีสีเข้มกว่าและเป็นเม็ดเล็ก ๆ (เอนโดพลาสซึม) มีบทบาทสำคัญในการรักษาความคงตัวของรูปร่างของ ciliates
ในชั้นนอก (ใน ectoplasm) ของร่างกายของรองเท้าที่มีชีวิตสามารถมองเห็นแท่งสั้นจำนวนมากได้อย่างชัดเจนซึ่งตั้งฉากกับพื้นผิว (รูปที่ 85, 7) การก่อตัวเหล่านี้เรียกว่าไตรโคซิสต์ ฟังก์ชั่นของพวกเขาน่าสนใจมากและเชื่อมโยงกับการป้องกันที่ง่ายที่สุด ด้วยแรงสั่นสะเทือนทางกล สารเคมี หรือการระคายเคืองที่รุนแรงอื่นๆ ไทรโคซิสต์จะถูกเหวี่ยงออกไปด้วยแรง กลายเป็นเส้นยาวบางๆ ที่จะไปโดนนักล่าที่จู่โจมรองเท้า Trichocysts เป็นการป้องกันที่มีประสิทธิภาพ พวกมันถูกจัดเรียงอย่างสม่ำเสมอระหว่าง cilia เพื่อให้จำนวนไตรโคซิสต์ใกล้เคียงกับจำนวน cilia โดยประมาณ แทนที่จะใช้ไตรโคซิสต์ (“ช็อต”) ไทรโคซิสต์ชนิดใหม่จะพัฒนาในชั้นนอกสุดของรองเท้า
ด้านหนึ่งประมาณกึ่งกลางลำตัว (รูปที่ 85, 5) รองเท้ามีความหดหู่ค่อนข้างลึก นี่คือช่องปากหรือเพอริสโตม Cilia ตั้งอยู่ตามผนังของเพอริสโตมเช่นเดียวกับตามพื้นผิวของร่างกาย พวกมันได้รับการพัฒนาที่นี่อย่างมีพลังมากกว่าส่วนอื่นๆ ของร่างกาย cilia ที่มีระยะห่างอย่างใกล้ชิดเหล่านี้จัดเป็นสองกลุ่ม หน้าที่ของ cilia ที่มีความแตกต่างสูงเหล่านี้ไม่ได้เกี่ยวข้องกับการเคลื่อนไหว แต่เกี่ยวข้องกับโภชนาการ (รูปที่ 87)

รองเท้ากินอย่างไรและย่อยอย่างไร?
รองเท้าอยู่ในกลุ่ม ciliates ซึ่งเป็นอาหารหลักที่เป็นแบคทีเรีย นอกจากแบคทีเรียแล้ว พวกมันยังสามารถกลืนอนุภาคอื่นๆ ที่ลอยอยู่ในน้ำได้ โดยไม่คำนึงถึงคุณค่าทางโภชนาการของพวกมัน ตาในช่องปากสร้างการไหลของน้ำอย่างต่อเนื่องโดยมีอนุภาคแขวนลอยไปในทิศทางของช่องปากซึ่งอยู่ลึกลงไปในเพอริสโตม เศษอาหารขนาดเล็ก (ส่วนใหญ่มักเป็นแบคทีเรีย) จะแทรกซึมเข้าไปในคอหอยหลอดเล็ก ๆ และสะสมที่ด้านล่างของมัน ที่ขอบของเอ็นโดพลาสซึม การเปิดปากเปิดอยู่เสมอ บางทีมันอาจจะไม่ผิดที่จะบอกว่ารองเท้า ciliate เป็นหนึ่งในสัตว์ที่โลภมากที่สุด: มันกินอย่างต่อเนื่อง กระบวนการนี้ถูกขัดจังหวะในบางช่วงเวลาของชีวิตที่เกี่ยวข้องกับการสืบพันธุ์และกระบวนการทางเพศเท่านั้น
ก้อนอาหารที่สะสมอยู่ที่ด้านล่างของคอหอยจะแตกออกจากด้านล่างของคอหอยและเมื่อรวมกับของเหลวจำนวนเล็กน้อยจะเข้าสู่เอนโดพลาสซึม ก่อตัวเป็นแวคิวโอลย่อยอาหาร หลังไม่ได้อยู่ที่ไซต์ของการก่อตัวของมัน แต่ตกลงไปในกระแสของเอนโดพลาสซึมทำให้เส้นทางที่ค่อนข้างซับซ้อนและสม่ำเสมอในร่างกายของรองเท้าเรียกว่า cyclosis ของ vacuole ย่อยอาหาร (รูปที่ 88) ในระหว่างการเดินทางที่ค่อนข้างยาว (ที่อุณหภูมิห้อง ใช้เวลาประมาณหนึ่งชั่วโมง) ของแวคิวโอลย่อยอาหาร การเปลี่ยนแปลงหลายอย่างเกิดขึ้นภายในแวคิวโอลที่เกี่ยวข้องกับการย่อยอาหารในนั้น
ในที่นี้เช่นเดียวกับในอะมีบาและแฟลเจลเลตบางชนิด การย่อยภายในเซลล์โดยทั่วไปจะเกิดขึ้น จากเอ็นโดพลาสซึมที่อยู่รอบๆ แวคิวโอลย่อยอาหาร เอ็นไซม์ย่อยอาหารเข้าไปซึ่งทำหน้าที่เกี่ยวกับอนุภาคอาหาร ผลิตภัณฑ์ย่อยอาหารจะถูกดูดซึมผ่านแวคิวโอลย่อยอาหารเข้าสู่เอนโดพลาสซึม
ในระหว่างการ cyclosis ของ vacuole ย่อยอาหารจะมีการแทนที่หลายขั้นตอนของการย่อยอาหาร ในช่วงแรกหลังจากการก่อตัวของแวคิวโอล การเติมของเหลวจะแตกต่างจากของเหลวในสิ่งแวดล้อมเพียงเล็กน้อย ในไม่ช้าเอ็นไซม์ย่อยอาหารก็เริ่มเข้าสู่ vacuole จากเอนโดพลาสซึมและปฏิกิริยาของสิ่งแวดล้อมภายในนั้นจะกลายเป็นกรดอย่างรวดเร็ว การตรวจจับนี้ทำได้ง่ายโดยการเพิ่มตัวบ่งชี้บางอย่างลงในอาหาร ซึ่งสีของอาหารจะเปลี่ยนไปตามปฏิกิริยา (ที่เป็นกรด เป็นกลาง หรือเป็นด่าง) ของสิ่งแวดล้อม ในสภาพแวดล้อมที่เป็นกรดนี้ ระยะแรกของการย่อยอาหารจะเกิดขึ้น จากนั้นภาพจะเปลี่ยนไปและปฏิกิริยาภายในแวคิวโอลย่อยอาหารจะกลายเป็นด่างเล็กน้อย ภายใต้เงื่อนไขเหล่านี้มี ขั้นตอนถัดไปการย่อยภายในเซลล์ เฟสกรดมักจะสั้นกว่าเฟสอัลคาไล มันกินเวลาประมาณ 1/6-1/4 ของระยะเวลาทั้งหมดของแวคิวโอลย่อยอาหารในร่างกายของ ciliate อย่างไรก็ตาม อัตราส่วนของเฟสที่เป็นกรดและด่างอาจแตกต่างกันภายในขอบเขตที่ค่อนข้างกว้าง ขึ้นอยู่กับลักษณะของอาหาร
เส้นทางของ vacuole ย่อยอาหารในเอนโดพลาสซึมจบลงด้วยความจริงที่ว่ามันเข้าใกล้พื้นผิวของร่างกายและผ่าน pellicle เนื้อหาซึ่งประกอบด้วยของเหลวและเศษอาหารที่ไม่ได้แยกแยะถูกโยนออกไป - การถ่ายอุจจาระเกิดขึ้น กระบวนการนี้ตรงกันข้ามกับอะมีบาซึ่งการถ่ายอุจจาระสามารถเกิดขึ้นได้ทุกที่ในรองเท้าเช่นเดียวกับใน ciliates อื่น ๆ ถูกกักขังไว้เฉพาะบริเวณเฉพาะของร่างกายที่อยู่ด้านข้างหน้าท้อง (ตามอัตภาพเรียกว่าพื้นผิวของ สัตว์ที่วางช่อง perioral ) ประมาณกึ่งกลางระหว่างเพอริสโตมและส่วนหลังของร่างกาย
ดังนั้นการย่อยภายในเซลล์จึงเป็นกระบวนการที่ซับซ้อนซึ่งประกอบด้วยหลายขั้นตอนแทนที่กันอย่างต่อเนื่อง

การคำนวณแสดงให้เห็นว่าในเวลาประมาณ 30-45 นาที ปริมาตรของของไหลเท่ากับปริมาตรของตัว ciliate จะถูกขับออกจากรองเท้าผ่าน vacuoles ที่หดตัว ดังนั้นเนื่องจากกิจกรรมของ vacuoles ที่หดตัวจึงมีการไหลของน้ำอย่างต่อเนื่องผ่านร่างกายของ ciliate ซึ่งเข้ามาจากภายนอกทางปาก (พร้อมกับ vacuoles ย่อยอาหาร) เช่นเดียวกับ osmotically โดยตรงผ่าน pellicle แวคิวโอลที่หดตัวมีบทบาทสำคัญในการควบคุมการไหลของน้ำที่ไหลผ่านร่างกายของซิลิเอตและในการควบคุมแรงดันออสโมติก กระบวนการนี้ดำเนินไปตามหลักการในลักษณะเดียวกับในอะมีบา มีเพียงโครงสร้างของแวคิวโอลที่หดตัวเท่านั้นที่มีความซับซ้อนมากขึ้น
หลายปีที่ผ่านมา ในหมู่นักวิทยาศาสตร์ที่ศึกษาโปรโตซัว มีการโต้เถียงกันเกี่ยวกับคำถามที่ว่า มีโครงสร้างใดๆ ในไซโตพลาสซึมที่เกี่ยวข้องกับการปรากฏตัวของแวคิวโอลที่หดตัวหรือไม่ หรือเกิดขึ้นใหม่ทุกครั้งหรือไม่ บน ciliate ที่มีชีวิตไม่สามารถสังเกตโครงสร้างพิเศษใด ๆ ที่จะเกิดขึ้นก่อนการก่อตัวของมัน หลังจากการหดตัวของ vacuole - systole - เกิดขึ้นไม่มีโครงสร้างใดที่มองเห็นได้ในไซโตพลาสซึมที่ไซต์ของ vacuole เดิม จากนั้นฟองโปร่งใสหรือช่อง adductor ก็ปรากฏขึ้นอีกครั้งซึ่งเริ่มมีขนาดเพิ่มขึ้น อย่างไรก็ตาม ไม่พบความเชื่อมโยงระหว่างแวคิวโอลที่เพิ่งเกิดใหม่กับแวคิวโอลที่มีอยู่ก่อนหน้านี้ ดูเหมือนว่าไม่มีความต่อเนื่องกันระหว่างวัฏจักรที่ต่อเนื่องกันของแวคิวโอลที่หดตัวและแวคิวโอลที่หดตัวใหม่ใดๆ จะก่อตัวขึ้นใหม่ในไซโตพลาสซึม อย่างไรก็ตาม วิธีการวิจัยพิเศษพบว่าไม่เป็นเช่นนั้นจริง การใช้กล้องจุลทรรศน์อิเล็กตรอนซึ่งให้กำลังขยายสูงมาก (มากถึง 100,000 เท่า) แสดงให้เห็นอย่างน่าเชื่อถือว่า ciliate มีไซโตพลาสซึมที่แตกต่างกันโดยเฉพาะในบริเวณที่เกิดแวคิวโอลหดตัวซึ่งประกอบด้วยหลอดที่บางที่สุดผสมเข้าด้วยกัน ดังนั้นจึงกลายเป็นว่า vacuole หดตัวไม่เกิดขึ้นในไซโตพลาสซึมบน " ที่ว่างเปล่า” แต่บนพื้นฐานของออร์แกนอยด์เซลล์พิเศษก่อนหน้าซึ่งหน้าที่ของมันคือการก่อตัวของแวคิวโอลที่หดตัว
เช่นเดียวกับโปรโตซัว ciliates มีนิวเคลียสของเซลล์ อย่างไรก็ตาม ในแง่ของโครงสร้างของอุปกรณ์นิวเคลียร์ ciliates แตกต่างอย่างมากจากโปรโตซัวกลุ่มอื่นทั้งหมด
เครื่องมือนิวเคลียร์ของ ciliates มีลักษณะเป็นคู่ของมัน ซึ่งหมายความว่า ciliates มีนิวเคลียสที่แตกต่างกันสองประเภท - นิวเคลียสขนาดใหญ่หรือมาโครนิวเคลียสและนิวเคลียสขนาดเล็กหรือไมโครนิวเคลียส เรามาดูกันว่าโครงสร้างของอุปกรณ์นิวเคลียร์มีอยู่ใน ciliates ของรองเท้าอย่างไร (รูปที่ 85)
ในใจกลางของร่างกายของ ciliate (ที่ระดับของ peristome) จะมีการวางนิวเคลียสขนาดใหญ่ที่มีรูปร่างเป็นรูปไข่หรือถั่ว นี่คือแมโครนิวเคลียส ในบริเวณใกล้เคียงกัน จะมีนิวเคลียสที่สองที่เล็กกว่าหลายเท่า มักจะอยู่ติดกับมาโครนิวเคลียสค่อนข้างมาก นี่คือไมโครนิวเคลียส ความแตกต่างระหว่างนิวเคลียสทั้งสองนี้ไม่เพียงแต่มีขนาดเท่านั้น แต่ยังมีความสำคัญมากกว่าและส่งผลต่อโครงสร้างของพวกมันอย่างลึกซึ้ง
มาโครนิวเคลียสเมื่อเปรียบเทียบกับไมโครนิวเคลียสนั้นเข้มข้นกว่ามากในสารนิวเคลียร์พิเศษ (โครมาตินหรือกรดดีออกซีไรโบนิวคลีอิกหรือ DNA แบบย่อที่แม่นยำกว่า) ซึ่งเป็นส่วนหนึ่งของโครโมโซม
การศึกษาล่าสุดได้แสดงให้เห็นว่าแมโครนิวเคลียสมีหลายสิบ (และในบาง ciliates และหลายร้อย) ครั้ง จำนวนมากโครโมโซมมากกว่าไมโครนิวเคลียส มาโครนิวเคลียสเป็นนิวเคลียส multichromosomal (polyploid) ชนิดพิเศษ ดังนั้นความแตกต่างระหว่างไมโครและมาโครนิวเคลียสจึงส่งผลต่อองค์ประกอบโครโมโซมซึ่งกำหนดความสมบูรณ์ของสารนิวเคลียร์ - โครมาติน
ใน ciliates ชนิดหนึ่งที่พบบ่อยที่สุด - รองเท้า(Paramecium caudatum) - มีมาโครนิวเคลียสหนึ่งตัว (ย่อมาจาก Ma) และไมโครนิวเคลียสหนึ่งตัว (ย่อมาจาก Mi) โครงสร้างของอุปกรณ์นิวเคลียร์นี้เป็นลักษณะของ ciliates จำนวนมาก คนอื่นอาจมี Ma และ Mi หลายตัว แต่ลักษณะเฉพาะของ ciliates ทั้งหมดคือการสร้างความแตกต่างของนิวเคลียสออกเป็นสองกลุ่มที่แตกต่างกันในเชิงคุณภาพเป็น Ma และ Mi หรือกล่าวอีกนัยหนึ่งคือปรากฏการณ์ของการเกิดคู่นิวเคลียส
ciliates สืบพันธุ์อย่างไร? ให้เรากลับมาเป็นตัวอย่างอีกครั้งสำหรับรองเท้า infusoria หากคุณปลูกรองเท้าเพียงสำเนาเดียวในภาชนะขนาดเล็ก (ไมโครอควาเรียม) จากนั้นในหนึ่งวันจะมีสอง ciliates และมักจะสี่ สิ่งนี้เกิดขึ้นได้อย่างไร? หลังจากว่ายน้ำและให้อาหารเป็นระยะเวลาหนึ่ง ciliate จะยืดออกบ้าง จากนั้นตรงกึ่งกลางของร่างกายมีการหดตัวตามขวางที่ลึกขึ้นเรื่อย ๆ (รูปที่ 90) ในท้ายที่สุด ciliates อย่างที่เคยเป็นมานั้นถูกมัดไว้ครึ่งหนึ่งและได้บุคคลสองคนจากบุคคลหนึ่งคนในขั้นต้นค่อนข้างเล็กกว่าผู้ปกครอง กระบวนการฟิชชันทั้งหมดใช้เวลาประมาณหนึ่งชั่วโมงที่อุณหภูมิห้อง การศึกษากระบวนการภายในแสดงให้เห็นว่าก่อนที่การหดตัวตามขวางจะปรากฏขึ้น กระบวนการฟิชชันของอุปกรณ์นิวเคลียร์ก็เริ่มต้นขึ้น Mi ถูกแชร์ก่อนและต่อจากนี้คือ Ma เราจะไม่หยุดที่นี่ การพิจารณาอย่างละเอียดกระบวนการแตกตัวของนิวเคลียร์และเราทราบเพียงว่า Mi ถูกแบ่งโดยไมโทซีสในขณะที่การแบ่งตัวของ Ma นั้นมีลักษณะคล้ายกับการแยกตัวของนิวเคลียร์โดยตรง - อะมิโทซิส กระบวนการสืบพันธุ์แบบไม่อาศัยเพศของ ciliates รองเท้า ดังที่เราเห็น คล้ายกับการสืบพันธุ์แบบไม่อาศัยเพศของอะมีบาและแฟลเจลเลต ในทางตรงกันข้าม ciliates ในกระบวนการของการสืบพันธุ์แบบไม่อาศัยเพศมักจะถูกแบ่งออก ในขณะที่ในแฟลเจลเลต ระนาบการแบ่งจะขนานกับแกนตามยาวของร่างกาย
ในระหว่างการแบ่งตัว การปรับโครงสร้างภายในลึกของร่างกาย ciliate เกิดขึ้น เพอริสโตมใหม่ 2 อัน คอหอย 2 อัน และปากเปิด 2 อัน ในเวลาเดียวกันการแบ่งนิวเคลียสพื้นฐานของ cilia จะถูกกำหนดเวลาเนื่องจากมีการสร้าง cilia ใหม่ หากจำนวนของ cilia ไม่เพิ่มขึ้นในระหว่างการสืบพันธุ์ ผลของการแบ่งแต่ละส่วน ลูกสาวแต่ละคนจะได้รับจำนวน cilia ของแม่ประมาณครึ่งหนึ่ง ซึ่งจะทำให้ "ศีรษะล้าน" ของ ciliates สมบูรณ์ อันที่จริงสิ่งนี้ไม่ได้เกิดขึ้น

ในบางครั้ง ciliates ส่วนใหญ่รวมทั้งรองเท้ามีรูปแบบที่พิเศษและแปลกประหลาดอย่างยิ่งของกระบวนการทางเพศซึ่งเรียกว่าการผันคำกริยา เราจะไม่วิเคราะห์รายละเอียดการเปลี่ยนแปลงทางนิวเคลียร์ที่ซับซ้อนทั้งหมดที่มาพร้อมกับกระบวนการนี้โดยละเอียด แต่ให้สังเกตเฉพาะสิ่งที่สำคัญที่สุดเท่านั้น การผันคำกริยาดำเนินการดังนี้ (รูปที่ 91) สอง ciliates เข้าหากันถูกนำไปใช้อย่างใกล้ชิดโดยด้านข้างหน้าท้องและในรูปแบบนี้พวกเขาจะว่ายน้ำค่อนข้าง เวลานานกัน (ที่รองเท้าประมาณ 12 ชั่วโมงที่อุณหภูมิห้อง) คอนจูกันต์ก็แยกจากกัน เกิดอะไรขึ้นในร่างกายของ ciliates ระหว่างการผันคำกริยา? สาระสำคัญของกระบวนการเหล่านี้มีดังนี้ (รูปที่ 91) นิวเคลียสขนาดใหญ่ (มาโครนิวเคลียส) จะยุบตัวและค่อยๆ ละลายในไซโตพลาสซึม การแบ่งตัวครั้งแรกของไมโครนิวเคลียส นิวเคลียสบางส่วนที่เกิดขึ้นจากการแยกตัวถูกทำลาย (ดูรูปที่ 91) คอนจูกันต์แต่ละตัวยังคงมีนิวเคลียสอยู่สองนิวเคลียส หนึ่งในนิวเคลียสเหล่านี้ยังคงอยู่ในบุคคลที่มันถูกสร้างขึ้น (นิวเคลียสนิ่ง) ในขณะที่อีกนิวเคลียสเคลื่อนเข้าสู่คู่ของการผันคำกริยา (นิวเคลียสการโยกย้าย) และรวมเข้ากับนิวเคลียสที่อยู่กับที่ ดังนั้นในแต่ละคอนจูกันต์ในขั้นตอนนี้จะมีนิวเคลียสหนึ่งอันเกิดขึ้นจากการหลอมรวมของนิวเคลียสที่อยู่กับที่และนิวเคลียสที่เคลื่อนที่ นิวเคลียสที่ซับซ้อนนี้เรียกว่าซินคาเรียน การก่อตัวของ synkaryon ไม่มีอะไรมากไปกว่ากระบวนการปฏิสนธิ และในสิ่งมีชีวิตหลายเซลล์ ช่วงเวลาสำคัญของการปฏิสนธิคือการรวมตัวของนิวเคลียสของเซลล์สืบพันธุ์ ใน ciliates เซลล์สืบพันธุ์ไม่ได้เกิดขึ้นมีเพียงนิวเคลียสเพศที่ผสานเข้าด้วยกัน ดังนั้นการปฏิสนธิระหว่างกันจึงเกิดขึ้น
ไม่นานหลังจากการก่อตัวของ synkaryon คอนจูกันก็แยกจากกัน ตามโครงสร้างของเครื่องมือนิวเคลียร์ของพวกเขา ในขั้นตอนนี้พวกเขายังมีความแตกต่างอย่างมากจาก ciliates ที่เป็นกลาง (ไม่คอนจูเกต) ตามปกติ เนื่องจากมีนิวเคลียสเพียงอันเดียว ในอนาคตเนื่องจากการซิงคาเรียน อุปกรณ์นิวเคลียร์แบบปกติจึงได้รับการฟื้นฟู synkaryon ถูกแบ่งออก (หนึ่งครั้งหรือมากกว่า) ส่วนหนึ่งของผลิตภัณฑ์ของแผนกนี้ ผ่านการเปลี่ยนแปลงที่ซับซ้อนที่เกี่ยวข้องกับการเพิ่มจำนวนโครโมโซมและการเสริมคุณค่าในโครมาติน จะกลายเป็นมาโครนิวเคลียส อื่น ๆ ยังคงลักษณะโครงสร้างของไมโครนิวเคลียส ด้วยวิธีนี้ คุณลักษณะของอุปกรณ์นิวเคลียร์และแบบฉบับของ ciliates ได้รับการฟื้นฟู หลังจากนั้น ciliates จะเริ่มการสืบพันธุ์แบบไม่อาศัยเพศโดยการแยกตัวออกจากกัน
ดังนั้น กระบวนการคอนจูเกตจึงรวมช่วงเวลาทางชีววิทยาที่สำคัญสองช่วงเวลา: การปฏิสนธิและการฟื้นฟูมาโครนิวเคลียสอันเนื่องมาจากซินคาเรียน
ความหมายทางชีวภาพของการผันคำกริยาคืออะไร มีบทบาทอย่างไรในชีวิตของ ciliates? เราไม่สามารถเรียกมันว่าการสืบพันธุ์ได้เพราะไม่มีการเพิ่มจำนวนบุคคล คำถามข้างต้นถูกใช้เป็นสื่อสำหรับการศึกษาทดลองจำนวนมากที่ดำเนินการในหลายประเทศ ผลลัพธ์หลักของการศึกษาเหล่านี้มีดังนี้ ประการแรกการผันคำกริยาเช่นเดียวกับกระบวนการทางเพศอื่น ๆ ซึ่งหลักการทางพันธุกรรมสองประการ (บิดาและมารดา) รวมกันเป็นหนึ่งสิ่งมีชีวิตนำไปสู่การเพิ่มขึ้นของความแปรปรวนทางพันธุกรรมความหลากหลายทางพันธุกรรม ความแปรปรวนทางพันธุกรรมที่เพิ่มขึ้นจะเพิ่มความสามารถในการปรับตัวของสิ่งมีชีวิตให้เข้ากับสภาพแวดล้อม ด้านที่สองที่สำคัญทางชีวภาพของการผันคำกริยาคือการพัฒนามาโครนิวเคลียสอันเนื่องมาจากผลิตภัณฑ์ฟิชชันของซินคาเรียนและในเวลาเดียวกันการทำลายของเก่า ข้อมูลการทดลองแสดงให้เห็นว่าเป็นมาโครนิวเคลียสที่มีบทบาทสำคัญในชีวิตของ ciliates มันควบคุมกระบวนการชีวิตหลักทั้งหมดและกำหนดกระบวนการที่สำคัญที่สุดของพวกเขา - การก่อตัว (การสังเคราะห์) ของโปรตีนที่ถือเป็นส่วนหลักของโปรโตพลาสซึมของเซลล์ที่มีชีวิต ด้วยการสืบพันธุ์แบบไม่อาศัยเพศที่ยืดเยื้อโดยการแบ่งแยก กระบวนการพิเศษของ "ความชรา" ของมาโครนิวเคลียส และในเวลาเดียวกันของเซลล์ทั้งหมด เกิดขึ้น: กิจกรรมของกระบวนการเผาผลาญอาหารลดลง อัตราการแบ่งตัวลดลง หลังจากการผันคำกริยา (ในขณะที่เราเห็นมาโครนิวเคลียสเก่าถูกทำลาย) อัตราการเผาผลาญและอัตราการแบ่งตัวจะกลับคืนมา เนื่องจากกระบวนการปฏิสนธิเกิดขึ้นระหว่างการผันคำกริยาซึ่งในสิ่งมีชีวิตอื่น ๆ ส่วนใหญ่เกี่ยวข้องกับการสืบพันธุ์และการปรากฏตัวของคนรุ่นใหม่ใน ciliates บุคคลที่เกิดขึ้นหลังจากการผันคำกริยาก็ถือได้ว่าเป็นรุ่นทางเพศใหม่ที่เกิดขึ้นที่นี่เช่นกัน เกิดจากการ “ฟื้นฟู” ของเก่า
ในตัวอย่างของรองเท้า ciliates เราได้พบกับตัวแทนทั่วไปของกลุ่ม ciliates ที่กว้างขวาง อย่างไรก็ตาม คลาสนี้มีเอกลักษณ์เฉพาะด้วยความหลากหลายของสายพันธุ์ทั้งในด้านโครงสร้างและการใช้ชีวิต มาดูรูปแบบที่มีลักษณะเฉพาะและน่าสนใจกันดีกว่า
ใน ciliates cilia ของรองเท้าจะครอบคลุมพื้นผิวทั้งหมดของร่างกายอย่างสม่ำเสมอ นี่คือลักษณะเฉพาะของโครงสร้าง (Holotricha) ciliates จำนวนมากมีลักษณะที่แตกต่างกันของการพัฒนาฝาครอบเลนส์ปรับเลนส์ ความจริงก็คือว่า cilia ของ ciliates มีความสามารถเมื่อรวมกันเพื่อสร้างคอมเพล็กซ์ที่ซับซ้อนมากขึ้น ตัวอย่างเช่น มักสังเกตว่า cilia อยู่ในแถวหนึ่งหรือสองแถวใกล้กัน (ติดกัน) เข้าด้วยกัน ก่อตัวเป็นจาน ซึ่งเหมือนกับ cilia สามารถตีได้ การก่อตัวที่หดตัวของแผ่นลามิเนตดังกล่าวเรียกว่าเมมเบรนลา (ถ้าสั้น) หรือเมมเบรน (ถ้ายาว) ในกรณีอื่น cilia จะรวมกันเป็นมัดแน่น การก่อตัวเหล่านี้ - cirres - คล้ายกับแปรงซึ่งมีขนแต่ละเส้นติดกัน การก่อตัวของเลนส์ปรับเลนส์ที่ซับซ้อนต่าง ๆ เป็นลักษณะของ ciliates จำนวนมาก บ่อยครั้งที่ฝาครอบเลนส์ปรับเลนส์ไม่พัฒนาเท่า ๆ กัน แต่เฉพาะในบางส่วนของร่างกายเท่านั้น
เป่าแตร INFUSORIA (STENTOR POLYMORPHIC)
ในน้ำจืดสายพันธุ์ ciliates ที่สวยงามขนาดใหญ่ที่เป็นของ ชนิดของนักเป่าแตร(สเตนเตอร์). ชื่อนี้ค่อนข้างสอดคล้องกับรูปร่างของสัตว์เหล่านี้ซึ่งคล้ายกับท่อ (รูปที่ 92) ซึ่งเปิดกว้างที่ปลายด้านหนึ่ง เมื่อรู้จักกับนักเป่าแตรสดครั้งแรก เราจะสังเกตเห็นคุณลักษณะหนึ่งที่ไม่ใช่ลักษณะของรองเท้า เมื่อเกิดการระคายเคืองเพียงเล็กน้อย รวมถึงกลไก (เช่น ใช้ดินสอเคาะกระจกที่มีหยดน้ำมีแตร) ร่างกายจะหดตัวอย่างรวดเร็วและเร็วมาก (ในเสี้ยววินาที) โดยใช้เวลาเกือบปกติ ทรงกลม จากนั้นค่อนข้างช้า (วัดเวลาเป็นวินาที) นักเป่าแตรจะยืดออกโดยใช้รูปร่างลักษณะเฉพาะของเขา ความสามารถของนักเป่าแตรในการหดตัวอย่างรวดเร็วนี้เกิดจากการมีเส้นใยกล้ามเนื้อพิเศษอยู่ตามร่างกายและในชั้นนอกสุด ดังนั้นระบบกล้ามเนื้อสามารถพัฒนาในสิ่งมีชีวิตที่มีเซลล์เดียวได้

มีสปีชีส์ในสกุลทรัมเป็ตซึ่งบางชนิดมีลักษณะเป็นสีที่ค่อนข้างสว่าง พบมากในน้ำจืด เป่าแตรสีน้ำเงิน(Stentor coeruleus) ซึ่งจะมีสีฟ้าสดใส สีของแตรเป่าแตรนี้เกิดจากการที่เม็ดสีน้ำเงินที่เล็กที่สุดอยู่ในชั้นนอกสุดของมัน
แตรทรัมเป็ตอีกชนิดหนึ่ง (Stentor polymorphus) มักมีสีเขียว สาเหตุของสีนี้ค่อนข้างแตกต่าง สีเขียวเนื่องจากสาหร่ายสีเขียวที่มีเซลล์เดียวขนาดเล็กอาศัยอยู่และทวีคูณในเอนโดพลาสซึมของ ciliate ซึ่งทำให้ร่างกายของนักเป่าแตรมีสีเฉพาะ Stentor polymorphus เป็นตัวอย่างทั่วไปของการอยู่ร่วมกันที่เป็นประโยชน์ร่วมกัน - symbiosis นักเป่าแตรและสาหร่ายมีความสัมพันธ์ซึ่งกันและกัน: นักเป่าแตรปกป้องสาหร่ายที่อาศัยอยู่ในร่างกายและจัดหาคาร์บอนไดออกไซด์ที่เกิดจากการหายใจ ในส่วนของสาหร่ายนั้นสาหร่ายจะให้ออกซิเจนแก่นักเป่าแตรซึ่งปล่อยออกมาในกระบวนการสังเคราะห์ด้วยแสง เห็นได้ชัดว่าสาหร่ายบางส่วนถูกย่อยโดย ciliates ซึ่งเป็นอาหารสำหรับนักเป่าแตร
นักเป่าแตรว่ายช้าๆ ในน้ำโดยให้ปลายกว้างไปข้างหน้า แต่พวกมันยังสามารถยึดติดกับพื้นผิวได้ชั่วคราวโดยส่วนท้ายที่แคบด้านหลังของร่างกายซึ่งมีตัวดูดขนาดเล็กเกิดขึ้น
ในร่างกายของนักเป่าแตร เราสามารถแยกความแตกต่างของบริเวณลำตัวที่ขยายจากด้านหลังไปด้านหน้าและช่องรอบวงกว้าง (รอบนอก) ซึ่งตั้งเกือบตั้งฉากกับมัน ฟิลด์นี้คล้ายกับกรวยแบนแบบอสมมาตร โดยที่ขอบด้านหนึ่งมีช่อง - คอหอยที่นำไปสู่เอนโดพลาสซึมของ ciliates ลำตัวของคนเป่าแตรถูกปกคลุมด้วยขนสั้นตามยาวเป็นแถว บริเวณใกล้ปาก (adoral) ของเมมเบรนลาที่พัฒนาขึ้นอย่างมีประสิทธิภาพตามแนวขอบของสนาม peristomal ตั้งอยู่ในวงกลม (รูปที่ 92) โซนนี้ประกอบด้วยแผ่น ciliated จำนวนมากซึ่งแต่ละแผ่นจะประกอบด้วย cilia จำนวนมากติดกันซึ่งอยู่ในสองแถวที่มีระยะห่างกันอย่างใกล้ชิด
ในบริเวณช่องเปิดช่องปากเยื่อหุ้มในช่องปากจะพันไปทางคอหอยทำให้เกิดเกลียวซ้าย การไหลของน้ำที่เกิดจากการสั่นของเยื่อหุ้มช่องท้องจะพุ่งไปที่ช่องเปิดของปาก (เข้าไปในความลึกของช่องทางที่เกิดจากส่วนหน้าของร่างกาย) นอกจากน้ำแล้ว เศษอาหารที่ลอยอยู่ในน้ำยังเข้าสู่คอหอยอีกด้วย วัตถุที่เป็นอาหารของนักเป่าแตรมีความหลากหลายมากกว่ารองเท้า นอกจากแบคทีเรียแล้ว มันกินโปรโตซัวขนาดเล็ก (เช่น แฟลเจลเลต) สาหร่ายที่มีเซลล์เดียว ฯลฯ
คนเป่าแตรมีแวคิวโอลหดตัวที่พัฒนามาอย่างดีซึ่งมีโครงสร้างเฉพาะ อ่างเก็บน้ำกลางตั้งอยู่ในส่วนหน้าที่สามของร่างกาย อยู่ใต้ช่องเปิดของปากเล็กน้อย สองช่อง adducting ยาวออกจากมัน หนึ่งในนั้นไปจากอ่างเก็บน้ำไปยังส่วนท้ายของร่างกายส่วนที่สองตั้งอยู่ในพื้นที่ของสนาม peristomal ขนานกับโซน perioral ของเยื่อหุ้มเซลล์
เป่าแตร infusoria เป็นวัตถุที่ชื่นชอบสำหรับการวิจัยเชิงทดลองเกี่ยวกับการฟื้นฟู การทดลองหลายครั้งได้พิสูจน์แล้วว่าความสามารถในการสร้างใหม่ของแตร ciliate กับมีดผ่าตัดบาง ๆ สามารถตัดเป็นหลายส่วน และแต่ละส่วนผ่าน เวลาอันสั้น(หลายชั่วโมง วันละครั้งหรือนานกว่านั้น) จะกลายเป็นนักเป่าแตรตัวเล็กที่สร้างขึ้นตามสัดส่วน ซึ่งเป็นผลมาจากการให้อาหารอย่างกระฉับกระเฉง ถึงขนาดปกติสำหรับสายพันธุ์นี้ เพื่อให้กระบวนการกู้คืนสมบูรณ์ ชิ้นส่วนที่สร้างใหม่ต้องมีมาโครนิวเคลียสที่มีลูกปัดอย่างน้อยหนึ่งส่วน
ดังที่เราเห็นแล้ว นักเป่าแตรมีตาที่แตกต่างกัน: ในอีกด้านหนึ่งพวกมันสั้นครอบคลุมทั้งร่างกายและอีกด้านหนึ่งมีเยื่อหุ้มเซลล์ใกล้ปาก ตามลักษณะเฉพาะของโครงสร้างนี้ การปลด ciliates ซึ่งเป็นของนักเป่าแตร ได้รับการตั้งชื่อ ciliated ciliates(เฮเทอโรทริชา).
อินฟูโซเรีย บูร์ซาเรีย (BURSARIA TRUNCATELLA)
ตัวแทนที่น่าสนใจอันดับสองของ ciliary ciliates มักพบในน้ำจืด bursaria(Bursaria truncatella, รูปที่ 93). นี่คือยักษ์ในหมู่ ciliates: ขนาดของมันสามารถสูงถึง 2 มม. ขนาดทั่วไปที่สุดคือ 0.5-1.0 มม. Bursaria มองเห็นได้ชัดเจนด้วยตาเปล่า ตามชื่อของมัน บูร์ซาเรียมีรูปร่างเหมือนกระเป๋า โดยเปิดกว้างที่ส่วนหน้า (บูร์ซาเป็นคำภาษาละติน แปลเป็นภาษารัสเซียแปลว่า "กระเป๋าเงิน", "กระเป๋า") และขยายออกเล็กน้อยที่ปลายด้านหลัง ร่างกายของ ciliate ทั้งหมดถูกปกคลุมด้วยแถวตามยาวของ cilia สั้น การตีของพวกมันทำให้สัตว์เคลื่อนที่ไปข้างหน้าค่อนข้างช้า Bursaria แหวกว่ายราวกับว่า "พลิกตัว" จากทางด้านข้าง

จากส่วนหน้าสู่ส่วนลึกของร่างกาย (ประมาณ 2/3 ของความยาว) ความกดอากาศในช่องท้องจะยื่นออกมา - เพอริสโตม ทางหน้าท้องจะสื่อสารกับสภาพแวดล้อมภายนอกผ่านช่องแคบ ๆ ที่ด้านหลังช่องเพอริสโตมไม่สื่อสารกับสภาพแวดล้อมภายนอก หากคุณดูที่ส่วนตัดขวางของส่วนบนที่สามของร่างกายของ bursaria (รูปที่ 93, B) คุณจะเห็นว่าช่องเพอริสโตมครอบครองส่วนใหญ่ของร่างกายในขณะที่ไซโตพลาสซึมล้อมรอบมันในรูปแบบของขอบ
ที่ส่วนหน้าของร่างกายทางด้านซ้าย โซน perioral (adoral) membranella (รูปที่ 93, 4) ได้รับการพัฒนาอย่างทรงพลังมากมีต้นกำเนิดในเบอร์ซาเรีย มันลงไปในความลึกของโพรงเพอริสโตม เลี้ยวซ้าย บริเวณความรักขยายไปถึงส่วนที่ลึกที่สุดของเพอริสโตม ไม่มีการก่อตัวของเลนส์ปรับเลนส์อื่น ๆ ในช่อง peristomal นอกเหนือจากเยื่อหุ้มในช่องท้อง ยกเว้นแถบปรับเลนส์ที่วิ่งไปตามด้านท้องของช่องเพอริสโตม (รูปที่ 93, 10) ข้างใน ผนังด้านหลังช่อง peristomal มีช่องแคบตลอดความยาวเกือบทั้งหมด (รูปที่ 93, 7) ซึ่งขอบมักจะติดกันอย่างใกล้ชิด นี่คือการเปิดปาก ขอบของมันเคลื่อนออกจากกันในเวลาที่กินเท่านั้น
Bursaria ไม่มีความเชี่ยวชาญด้านอาหารอย่างแคบ แต่ส่วนใหญ่เป็นสัตว์กินเนื้อ เมื่อก้าวไปข้างหน้าก็จะพบกับสัตว์เล็กต่างๆ ต้องขอบคุณการทำงานของเยื่อหุ้มเซลล์ของบริเวณใกล้ปาก เหยื่อจึงถูกดึงเข้าไปในโพรง peristomal อันกว้างใหญ่ ซึ่งมันไม่สามารถว่ายออกไปได้อีกต่อไป วัตถุที่เป็นอาหารถูกกดลงที่ผนังด้านหลังของโพรง peristomal และเจาะเข้าไปในเอนโดพลาสซึมผ่านรอยแยกในช่องปากที่ขยายออก บูร์ซาเรียนั้นตะกละตะกลามมาก พวกมันสามารถกลืนสิ่งของที่ค่อนข้างใหญ่ได้ ตัวอย่างเช่น อาหารโปรดของพวกมันคือ ciliates รองเท้า บูร์ซาเรียสามารถกลืนรองเท้าได้ 6-7 ตัวติดต่อกัน เป็นผลให้เกิด vacuoles ย่อยอาหารขนาดใหญ่มากในเอนโดพลาสซึมของเบอร์ซาเรีย
เครื่องมือนิวเคลียร์ของ bursaria ค่อนข้างซับซ้อน พวกมันมีมาโครนิวเคลียสรูปไส้กรอกยาวหนึ่งอันและมีไมโครนิวเคลียสขนาดเล็กจำนวนมาก (มากถึงประมาณ 30) จำนวนที่กระจัดกระจายแบบสุ่มในเอนโดพลาสซึมของซิลิเอต
Bursaria เป็นหนึ่งในไม่กี่สายพันธุ์ของ ciliates น้ำจืดที่ไม่มี vacuole ที่หดตัว วิธีการดูดซึมใน ciliate ขนาดใหญ่นี้ยังไม่ชัดเจนนัก ภายใต้ ectoplasm ของ bursaria ในส่วนต่าง ๆ ของร่างกายเราสามารถสังเกตรูปร่างและขนาดต่าง ๆ ของฟองของเหลว - vacuoles ซึ่งเปลี่ยนปริมาตร เห็นได้ชัดว่าสิ่งเหล่านี้ รูปร่างผิดปกติแวคิวโอลและสอดคล้องกับหน้าที่ของแวคิวโอลที่หดตัวของซิลิเอตอื่นๆ

เป็นเรื่องที่น่าสนใจที่จะสังเกตขั้นตอนต่อเนื่องของการสืบพันธุ์แบบไม่อาศัยเพศของ Bursaria ในระยะเริ่มต้นของการแบ่งตัว มีการลดลงอย่างสมบูรณ์ของช่องทั้งหมดของเพอริสโตมและบริเวณใกล้ปากของเยื่อหุ้มเซลล์ (รูปที่ 94) รักษาเฉพาะซีเลียมชั้นนอกเท่านั้น Infusoria อยู่ในรูปของไข่ หลังจากนั้นร่างกายจะเย็บด้วยร่องตามขวางเป็นสองส่วน ในแต่ละลูกสาวที่เป็นผลลัพธ์ ciliates ผ่านการเปลี่ยนแปลงที่ค่อนข้างซับซ้อน peristome และ perioral zone ของ membranella พัฒนาโดยผ่านการเปลี่ยนแปลงที่ค่อนข้างซับซ้อน ขั้นตอนการแบ่งเบอร์ซาเรียทั้งหมดดำเนินไปอย่างรวดเร็วและใช้เวลามากกว่าหนึ่งชั่วโมงเล็กน้อย
มันง่ายมากที่จะสังเกตกระบวนการชีวิตที่สำคัญอีกอย่างหนึ่งใน bursaria ซึ่งการเริ่มต้นนั้นสัมพันธ์กับสภาวะที่ไม่เอื้ออำนวยต่อ ciliates กระบวนการของการสร้างซีสต์ (encystation) ปรากฏการณ์นี้เป็นลักษณะเฉพาะของอะมีบา แต่ปรากฎว่าแม้แต่โปรโตซัวที่จัดอย่างซับซ้อนเช่น ciliates ก็สามารถผ่านเข้าสู่สภาวะที่ไม่ใช้งาน หากวัฒนธรรมที่ bursaria อาศัยอยู่ไม่ได้รับอาหารในเวลาหรือเย็นลงจากนั้นภายในไม่กี่ชั่วโมงก็จะเริ่มสร้างมวลรวม กระบวนการนี้ดำเนินการดังนี้ Bursarids เช่นเดียวกับก่อนการแบ่งจะสูญเสีย peristome และ perioral zone ของ membranells จากนั้นพวกเขาก็กลายเป็นทรงกลมอย่างสมบูรณ์หลังจากนั้นพวกเขาก็แยกแยะเปลือกสองชั้นที่มีรูปร่างลักษณะเฉพาะ (รูปที่ 94, D)
ในสถานะของซีสต์ bursaria อาจอยู่ได้นานหลายเดือน เมื่อเงื่อนไขที่เอื้ออำนวยเกิดขึ้นเปลือกของถุงน้ำจะแตกออก bursaria จะออกมาจากมัน พัฒนาเพอริสโตมและก้าวไปสู่ชีวิตที่กระฉับกระเฉง
สไตโลนิเชีย มิทิลุส
เครื่องมือปรับเลนส์ที่ซับซ้อนและแตกต่างกันอย่างมากมีเลนส์ ciliates ที่เกี่ยวข้องกับ การหลุดออกของทางเดินอาหาร(Hypotricha) หลายชนิดซึ่งอาศัยทั้งในน้ำจืดและน้ำทะเล หนึ่งในตัวแทนที่พบบ่อยที่สุดและพบบ่อยของสิ่งนี้ กลุ่มที่น่าสนใจเรียกได้ว่า stilonychia(สไตโลนิเชีย มิติลุส). นี่คือ ciliate ที่ค่อนข้างใหญ่ (ยาวไม่เกิน 0.3 มม.) อาศัยอยู่ที่ด้านล่างของอ่างเก็บน้ำน้ำจืด on พืชน้ำ(รูปที่ 95). ต่างจากรองเท้าแตะ เป่าแตร และเบอร์ซาเรีย สไตโลนีเชียขาดเลนส์ปรับเลนส์ต่อเนื่อง และเครื่องมือปรับเลนส์ทั้งหมดจะแสดงด้วยการสร้างเลนส์ปรับเลนส์จำนวนจำกัด

ร่างกายของ Stilonychia (เช่นเดียวกับ ciliates หน้าท้องอื่น ๆ ส่วนใหญ่) ถูกแบนอย่างมากในทิศทางของลำตัว - ช่องท้อง และด้านหลังและหน้าท้องส่วนหน้าและส่วนหลังมีความชัดเจน ด้านหน้าลำตัวค่อนข้างกว้าง ด้านหลังแคบลง เมื่อตรวจสอบสัตว์จากด้านท้องจะเห็นได้ชัดว่าในส่วนหน้าที่สามทางด้านซ้ายมีพินเนทและช่องปากที่จัดเรียงอย่างซับซ้อน
ที่ด้านหลัง cilia นั้นค่อนข้างหายากซึ่งไม่สามารถตีได้ พวกเขาสามารถเรียกได้ว่าเป็นขนแปรงยืดหยุ่นบาง ๆ พวกมันไม่มีการเคลื่อนไหวและไม่เกี่ยวอะไรกับการทำงานของการเคลื่อนไหว ตาเหล่านี้มักจะถูกกำหนดให้เป็นหน้าที่ที่สัมผัสได้และละเอียดอ่อน
การสร้างเลนส์ปรับเลนส์ทั้งหมดที่เกี่ยวข้องกับการเคลื่อนไหวและการจับอาหารจะกระจุกตัวอยู่ที่หน้าท้องของสัตว์ (รูปที่ 95) มีการก่อตัวคล้ายนิ้วหนาจำนวนเล็กน้อยซึ่งอยู่ในหลายกลุ่ม เหล่านี้เป็นตับแข็งในช่องท้อง แต่ละอันเป็นรูปแบบของเลนส์ปรับเลนส์ที่ซับซ้อนซึ่งเป็นผลมาจากการเชื่อมต่ออย่างใกล้ชิด (ติดกัน) ของตาแต่ละตัวหลายสิบตัว ขนแต่ละเส้นมารวมกันอย่างใกล้ชิดและเชื่อมต่อถึงกัน
ด้วยความช่วยเหลือของโรคตับแข็งสัตว์จะเคลื่อนที่อย่างรวดเร็ว "วิ่ง" ไปตามพื้นผิว นอกเหนือจาก "การคลาน" และ "การวิ่ง" บนพื้นผิวแล้ว stilonychia ยังสามารถกระโดดได้ค่อนข้างคมและแข็งแรงและแตกออกจากพื้นผิวทันที การเคลื่อนไหวที่เฉียบคมเหล่านี้ดำเนินการโดยใช้หางหางอันทรงพลังสองตัว (รูปที่ 95) ซึ่งไม่ได้มีส่วนร่วมในการ "คลาน" ตามปกติ
ตับแข็งชายขอบ (ขอบ) สองแถวตั้งอยู่ตามขอบของร่างกายทางด้านขวาและซ้าย จากขอบด้านขวาของสัตว์ พวกมันวิ่งไปตามร่างกายทั้งหมด ในขณะที่จากขอบด้านซ้ายจะไปถึงเฉพาะบริเวณเพอริสโตม การก่อตัวของเลนส์ปรับเลนส์เหล่านี้ใช้เพื่อขับเคลื่อนสัตว์เมื่อแยกออกจากพื้นผิวและแหวกว่ายอย่างอิสระในน้ำ
ดังนั้น เราจึงเห็นว่าเครื่องมือปรับเลนส์เลนส์เฉพาะทางของสไตโลนีเชียช่วยให้เคลื่อนไหวได้หลากหลายมาก ตรงกันข้ามกับการสไลด์ในน้ำธรรมดาๆ เช่น รองเท้าหรือนักเป่าแตร
เครื่องมือปรับเลนส์ที่เกี่ยวข้องกับการทำงานของโภชนาการก็ซับซ้อนเช่นกัน เราได้เห็นแล้วว่าช่องใกล้ปาก (เพอริสโตม) ซึ่งอยู่ด้านล่างของช่องเปิดปากที่นำไปสู่คอหอย ตั้งอยู่ครึ่งหน้าของสัตว์ทางด้านซ้าย ตามแนวขอบด้านซ้าย เริ่มจากส่วนหน้าสุดของร่างกาย มีเยื่อหุ้มเยื่อบุช่องท้อง (adoral) ที่พัฒนาขึ้นอย่างมาก ด้วยการตีพวกมันจะควบคุมการไหลของน้ำไปทางปาก นอกจากนี้ในพื้นที่ของช่อง peristomal มีเยื่อหุ้มหด (เมมเบรน) อีกสามแผ่นซึ่งขยายเข้าไปในคอหอยที่มีปลายด้านในและ cilia perioral พิเศษจำนวนหนึ่ง (รูปที่ 95) เครื่องมือที่ซับซ้อนทั้งหมดนี้ทำหน้าที่จับและนำอาหารเข้าปาก
Stilonychia เป็นโปรโตซัวชนิดหนึ่งที่มีวัตถุอาหารหลากหลายมาก สามารถเรียกได้ว่าเป็นสัตว์กินพืชทุกชนิด เธอกินได้เหมือนรองเท้า แบคทีเรีย ในบรรดาวัตถุที่เป็นอาหารของมันคือแฟลเจลเลต สาหร่ายเซลล์เดียว (มักเป็นไดอะตอม) ในที่สุด Stilonychia ยังสามารถเป็นนักล่า โจมตี ciliates อื่น ๆ ที่มีขนาดเล็กกว่าและกินพวกมัน
Stilonychia มีแวคิวโอลที่หดตัว ประกอบด้วยอ่างเก็บน้ำกลางซึ่งอยู่ที่ปลายด้านหลังด้านซ้ายของเพอริสโตม และคลองแอดดักเตอร์หนึ่งช่องหันกลับ
เครื่องมือนิวเคลียร์เช่นเคยใน ciliates ประกอบด้วยมาโครนิวเคลียสและไมโครนิวเคลียส
มาโครนิวเคลียสประกอบด้วยสองส่วนที่เชื่อมต่อกันด้วยการหดตัวบางๆ มีไมโครนิวเคลียสอยู่ 2 นิวเคลียส ตั้งอยู่ใกล้ทั้งสองซีกของหม่า
Stilonychia, bursaria บางส่วน, เป่าแตร - ทั้งหมดนี้เป็น ciliates ที่มีรายการอาหารหลากหลาย ความสามารถในการดูดซับอาหารต่าง ๆ เป็นลักษณะของ ciliates ส่วนใหญ่ อย่างไรก็ตาม ในหมู่พวกเขา เรายังสามารถพบสปีชีส์ดังกล่าวที่มีความเชี่ยวชาญอย่างเคร่งครัดเกี่ยวกับธรรมชาติของอาหาร
ผู้ล่าเหยื่อ
ในบรรดา ciliates มีนักล่าที่ "จู้จี้จุกจิก" มากเกี่ยวกับเหยื่อของพวกมัน ตัวอย่างที่ดีคือ infusoria Didynia(ดิดิเนียม นาสุทุม). ดิดิเนียมเป็นซิลิเอตที่ค่อนข้างเล็ก โดยมีความยาวเฉลี่ย 0.1-0.15 มม. ส่วนหน้าจะยาวในรูปแบบของงวงที่ส่วนท้ายของช่องเปิดปาก เครื่องมือปรับเลนส์นั้นแสดงด้วย cilia สองกลีบ (รูปที่ 96) Didinius แหวกว่ายอย่างรวดเร็วในน้ำ มักจะเปลี่ยนทิศทาง อาหารยอดนิยมของ Didinia คือรองเท้า ciliates ในกรณีนี้ ผู้ล่ามีขนาดเล็กกว่าเหยื่อ Didinius เจาะเหยื่อด้วยลำตัวแล้วค่อยๆอ้าปากกว้างขึ้นกลืนรองเท้าทั้งหมด! ในงวงมีเครื่องมือพิเศษที่เรียกว่าร็อด ประกอบด้วยแท่งที่แข็งแรงยืดหยุ่นจำนวนหนึ่งซึ่งอยู่ในไซโตพลาสซึมตามขอบงวง เป็นที่เชื่อกันว่าอุปกรณ์นี้เพิ่มความแข็งแรงของผนังงวงซึ่งไม่แตกเมื่อกลืนเหยื่อขนาดใหญ่เช่นนี้เมื่อเทียบกับดิดิเนียมเช่นรองเท้า Didinius - ตัวอย่าง วิธีสุดท้ายการปล้นสะดมระหว่างโปรโตซัว หากเราเปรียบเทียบดิดิเนียมที่กลืนเหยื่อ - รองเท้า - กับการปล้นสะดมในสัตว์ที่สูงกว่า เป็นการยากที่จะหาตัวอย่างที่คล้ายกัน

Didinius กลืน paramecia แน่นอนบวมมาก กระบวนการย่อยอาหารทำได้เร็วมาก ที่อุณหภูมิห้องใช้เวลาประมาณสองชั่วโมงเท่านั้น จากนั้นซากที่ไม่ได้แยกแยะก็จะถูกโยนทิ้ง และไดดิเนียมก็เริ่มออกล่าเหยื่ออีกราย การศึกษาพิเศษพบว่า "อาหาร" ประจำวันของ Didinia คือ 12 รองเท้า - ความอยากอาหารมหาศาลอย่างแท้จริง! ต้องระลึกไว้เสมอว่าบางครั้งการแบ่ง "การล่า" ครั้งต่อไปของ Didinia เนื่องจากขาดอาหาร Didinia จึงซึมซับได้ง่ายและหลุดออกจากซีสต์ได้ง่ายเช่นเดียวกัน
ciliates กินพืชเป็นอาหาร
พบน้อยกว่าการปล้นสะดมมาก พบในกลุ่ม ciliates "การกินเจล้วนๆ" - กินเฉพาะ อาหารจากพืช. หนึ่งในตัวอย่างบางส่วนของ "มังสวิรัติ" ciliates สามารถเป็นตัวแทนได้ สกุล passula(นัสซูล่า). วัตถุที่เป็นอาหารของมันคือสาหร่ายสีเขียวแกมน้ำเงิน (รูปที่ 97)

พวกมันเจาะเข้าไปในเอ็นโดพลาสซึมทางปากซึ่งอยู่ด้านข้างแล้วบิด ciliates ให้เป็นเกลียวแน่นซึ่งจะถูกย่อยทีละน้อย เม็ดสีสาหร่ายเข้าสู่ไซโตพลาสซึมของ ciliates บางส่วนและย้อมด้วยสีเขียวเข้มสดใส
ซูโวอิก้า (VORTICELLA NEBULIFERA)
ciliates กลุ่มที่น่าสนใจและค่อนข้างใหญ่ในแง่ของจำนวนสายพันธุ์คือรูปแบบนั่งติดกับพื้นผิวก่อตัว ตากลม(เพอริทริชา). สมาชิกที่พบมากที่สุดในกลุ่มนี้คือ suvoyki(สายพันธุ์ของสกุล Vorticella).
ซูวอยกีมีลักษณะเป็นดอกไม้งามสง่าเหมือนระฆังหรือดอกลิลลี่แห่งหุบเขานั่งบนก้านยาวซึ่งติดอยู่กับพื้นผิวด้วยปลาย ซูวอยก้าใช้ชีวิตส่วนใหญ่ในสภาพที่ติดอยู่กับพื้นผิว

พิจารณาโครงสร้างร่างกายของ ciliates ในสปีชีส์ต่างๆ ขนาดของพวกมันจะแตกต่างกันไปตามช่วงที่ค่อนข้างกว้าง (มากถึงประมาณ 150 ไมครอน) แผ่นช่องปาก (รูปที่ 98) ตั้งอยู่บนส่วนหน้าของร่างกายที่ขยายออกไปซึ่งไม่มี cilia อย่างสมบูรณ์ อุปกรณ์ปรับเลนส์ตั้งอยู่ตามขอบของดิสก์ในช่องปาก (peristomal) (รูปที่ 98) ในร่องพิเศษซึ่งด้านนอกจะมีลูกกลิ้ง (peristomal lip) เยื่อ ciliated สามแผ่นวิ่งไปตามขอบของลูกกลิ้ง ซึ่งสองแผ่นอยู่ในแนวตั้ง หนึ่ง (ด้านนอก) เป็นแนวนอน พวกมันก่อตัวเป็นเกลียวเต็มมากกว่าหนึ่งรอบ เยื่อเหล่านี้มีการเคลื่อนไหวที่กะพริบตลอดเวลา โดยกำหนดทิศทางการไหลของน้ำไปยังช่องเปิดปาก เครื่องมือในช่องปากเริ่มต้นค่อนข้างลึกด้วยช่องทางที่ขอบของสนาม peristomal (รูปที่ 98) ในความลึกซึ่งมีช่องปากที่นำไปสู่คอหอยสั้น Suvoys เช่นรองเท้ากินแบคทีเรีย ปากเปิดตลอดเวลาและมีน้ำไหลเข้าปากอย่างต่อเนื่อง
แวคิวโอลที่หดตัวหนึ่งอันที่ไม่มีคลองแอดดักเตอร์ตั้งอยู่ใกล้กับช่องเปิดปาก มาโครนิวเคลียสมีรูปร่างคล้ายริบบิ้นหรือคล้ายไส้กรอก ติดกับไมโครนิวเคลียสขนาดเล็กเพียงตัวเดียว
Suvoyka สามารถย่อก้านให้สั้นลงอย่างรวดเร็วซึ่งในเสี้ยววินาทีนั้นบิดเกลียวด้วยเหล็กไขจุก ในเวลาเดียวกัน ร่างกายของ ciliate ก็หดตัวเช่นกัน: แผ่น peristomal และเยื่อหุ้มเซลล์ถูกดึงเข้าด้านในและส่วนหน้าทั้งหมดจะปิดลง

คำถามเกิดขึ้นตามธรรมชาติ: เนื่องจากรถซูวอยติดอยู่กับพื้นผิว การตั้งถิ่นฐานในอ่างเก็บน้ำดำเนินการในลักษณะใด? สิ่งนี้เกิดขึ้นโดยการก่อตัวของเวทีลอยอิสระคนจรจัด ที่ส่วนท้ายของ ciliate body กลีบของ cilia จะปรากฏขึ้น (รูปที่ 99) ในเวลาเดียวกัน แผ่น peristomal จะหดกลับเข้าด้านใน และ ciliate ก็แยกออกจากก้าน คนจรจัดที่สามารถว่ายน้ำได้หลายชั่วโมง เหตุการณ์จะถูกเล่นออกมาใน กลับคำสั่ง: ciliate ยึดติดกับพื้นผิวด้วยส่วนหลังของมัน, ก้านเติบโต, กลีบหลังของ cilia ลดลง, แผ่น peristomal ยืดตรงที่ส่วนหน้า, และเยื่อ adoral เริ่มทำงาน การก่อตัวของคนเร่ร่อนในซูวอยก้ามักเกี่ยวข้องกับกระบวนการสืบพันธุ์แบบไม่อาศัยเพศ Infusoria บนก้านแบ่งและลูกสาวคนหนึ่ง (และบางครั้งทั้งคู่) กลายเป็นคนจรจัดและแหวกว่าย
ซูว็อกหลายประเภทสามารถยับยั้งได้ภายใต้สภาวะที่ไม่เอื้ออำนวย
ในบรรดา ciliates ที่อาศัยอยู่ซึ่งเป็นของกลุ่ม ciliated กลมมีเพียงไม่กี่ชนิดเท่านั้นเช่น suwoks ที่กล่าวถึงข้างต้นเป็นรูปแบบชีวิตที่โดดเดี่ยว สปีชีส์ส่วนใหญ่ที่นี่คือสิ่งมีชีวิตในยุคอาณานิคม
โดยปกติการตกเป็นอาณานิคมเป็นผลมาจากการสืบพันธุ์แบบไม่อาศัยเพศหรือการสืบพันธุ์อย่างไม่สมบูรณ์ ปัจเจกบุคคลซึ่งเกิดขึ้นจากการสืบพันธุ์จะมากหรือน้อยก็รักษาความสัมพันธ์ซึ่งกันและกันและรวมกันเป็นปัจเจกบุคคลที่มีลำดับชั้นสูง รวมกันเป็นหนึ่ง ปริมาณมากปัจเจกบุคคลซึ่งได้รับชื่อของอาณานิคม (เราได้พบกับตัวอย่างของสิ่งมีชีวิตในยุคอาณานิคมใน .แล้ว คลาสของแฟลเจลเลต.

อาณานิคมของ ciliates กลมนั้นเกิดขึ้นจากความจริงที่ว่าบุคคลที่แยกจากกันไม่ได้กลายเป็นคนจรจัด แต่ให้ติดต่อกันด้วยความช่วยเหลือของก้าน (รูปที่ 100) ในกรณีนี้ ต้นกำเนิดหลักของอาณานิคมเช่นเดียวกับกิ่งแรกไม่สามารถนำมาประกอบกับบุคคลใด ๆ ได้ แต่เป็นของทั้งอาณานิคม บางครั้งอาณานิคมประกอบด้วยบุคคลเพียงไม่กี่คนในขณะที่ ciliates สายพันธุ์อื่นจำนวนบุคคลในอาณานิคมสามารถเข้าถึงได้หลายร้อยคน อย่างไรก็ตาม การเติบโตของอาณานิคมไม่มีจำกัด เมื่อถึงขนาดของลักษณะเฉพาะของสปีชีส์นี้ อาณานิคมก็หยุดเพิ่มขึ้น และปัจเจกที่เกิดจากการแบ่งแยกพัฒนากลีบของตา กลายเป็นคนเร่ร่อนและว่ายออกไป ทำให้เกิดอาณานิคมใหม่
อาณานิคมของ ciliates กลมเป็นสองประเภท ในบางส่วน ก้านของอาณานิคมจะลดน้อยลง: เมื่อระคายเคือง เฉพาะบุคคลในสัญญาอาณานิคม วาดในพินเนท แต่ทั้งอาณานิคมทั้งหมดไม่ได้รับการเปลี่ยนแปลง (อาณานิคมประเภทนี้รวมถึงจำพวก สกุล Epistylis, Opercularia). ในส่วนอื่นๆ (เช่น สกุล Carchesium) ก้านของโคโลนีทั้งหมดสามารถหดตัวได้ เนื่องจากไซโตพลาสซึมผ่านกิ่งก้านสาขาทั้งหมดและเชื่อมโยงบุคคลในอาณานิคมเข้าด้วยกัน เมื่ออาณานิคมดังกล่าวระคายเคือง พวกมันจะหดตัวลงอย่างสิ้นเชิง อาณานิคมทั้งหมดในกรณีนี้มีปฏิกิริยาโดยรวมในฐานะปัจเจกบุคคลแบบอินทรีย์
ในบรรดา ciliates กลมๆ ที่เป็นอาณานิคมทั้งหมด สิ่งที่น่าสนใจเป็นพิเศษคือ บางที สวนสัตว์(ซูธัมเนียม อาร์บัสคูลา). อาณานิคมของ ciliate นี้มีความโดดเด่นด้วยความสม่ำเสมอเป็นพิเศษของโครงสร้าง นอกจากนี้ยังมีการสรุปปรากฏการณ์ทางชีววิทยาที่น่าสนใจของความหลากหลายในอาณานิคมด้วย

อาณานิคมโซตัมเนียดูเหมือนร่ม หนึ่ง หลัก ก้านของอาณานิคมเป็นกิ่งรอง (รูปที่ 101) ขนาดของอาณานิคมสำหรับผู้ใหญ่คือ 2-3 มม. จึงมองเห็นได้ชัดเจนด้วยตาเปล่า Zootamnii อาศัยอยู่ใน สระน้ำขนาดเล็กกับ น้ำสะอาด. อาณานิคมของพวกมันมักพบในพืชใต้น้ำ ส่วนใหญ่มักพบในเอโลเดีย (โรคระบาดในน้ำ)
ก้านของอาณานิคม Zootamnia นั้นหดตัวเนื่องจากไซโตพลาสซึมที่หดตัวผ่านกิ่งก้านทั้งหมดของอาณานิคมยกเว้นส่วนฐานของก้านหลัก ด้วยการลดลงที่เกิดขึ้นอย่างรวดเร็วและฉับพลัน อาณานิคมทั้งหมดรวมกันเป็นก้อน

Zootamnia โดดเด่นด้วยการจัดเรียงกิ่งก้านอย่างสม่ำเสมอ หนึ่งลำต้นหลักติดอยู่กับพื้นผิว กิ่งก้านหลักของอาณานิคมเก้ากิ่งแยกออกจากส่วนบนในระนาบตั้งฉากกับก้านซึ่งอยู่อย่างสม่ำเสมอโดยสัมพันธ์กันอย่างเคร่งครัด (รูปที่ 102, 6) กิ่งก้านรองขยายจากกิ่งเหล่านี้ซึ่งบุคคลในอาณานิคมนั่ง สาขารองแต่ละสาขาสามารถมีได้มากถึง 50 ciliates จำนวนบุคคลในอาณานิคมมีจำนวนถึง 2-3 พันคน
บุคคลส่วนใหญ่ของอาณานิคมในโครงสร้างของพวกเขามีลักษณะคล้ายกับรถ Suvoik เดี่ยวขนาดเล็กขนาด 40-60 ไมครอน แต่นอกเหนือจากบุคคลขนาดเล็กซึ่งเรียกว่า microzoids ในอาณานิคมของผู้ใหญ่ซึ่งอยู่ตรงกลางของกิ่งก้านหลักโดยเฉลี่ยแล้วบุคคลที่มีประเภทและขนาดแตกต่างไปจากเดิมอย่างสิ้นเชิง (รูปที่ 102, 5) เหล่านี้เป็นบุคคลทรงกลมขนาดใหญ่ที่มีเส้นผ่านศูนย์กลาง 200-250 ไมครอนซึ่งเกินปริมาตรของ microzoid มากกว่าร้อยเท่าหรือมากกว่าในมวล บุคคลขนาดใหญ่เรียกว่า macrozoids
ในโครงสร้างของพวกเขา พวกเขาแตกต่างจากบุคคลขนาดเล็กในอาณานิคมอย่างมีนัยสำคัญ เพอริสโตมของพวกมันไม่ได้แสดงออก: มันถูกดึงเข้าด้านในและไม่ทำงาน ตั้งแต่เริ่มต้นการพัฒนาจากไมโครซออยด์ มาโครโซออนก็หยุดกินอาหารเอง มันขาด vacuoles ย่อยอาหาร เห็นได้ชัดว่าการเติบโตของ macrozoid เกิดขึ้นจากค่าใช้จ่ายของสารที่เข้าสู่สะพานไซโตพลาสซึมที่เชื่อมต่อบุคคลทั้งหมดในอาณานิคมเข้าด้วยกัน ในส่วนของร่างกายของมาโครโซอันซึ่งติดอยู่กับก้านนั้นมีเมล็ดธัญพืชพิเศษ (แกรนูล) สะสมซึ่งดังที่เราจะได้เห็นกันว่าจะมีบทบาทสำคัญในชะตากรรมต่อไป macrozoids ทรงกลมขนาดใหญ่เหล่านี้คืออะไร บทบาททางชีวภาพของพวกมันในชีวิตของอาณานิคม Zootamnia คืออะไร? การสังเกตแสดงให้เห็นว่า macrozoids เป็นผู้เร่ร่อนในอนาคตซึ่งอาณานิคมใหม่จะพัฒนา ถึง ขนาดจำกัด, macrozoid พัฒนากลีบของ cilia แยกออกจากอาณานิคมและแหวกว่ายออกไป ในขณะเดียวกัน รูปร่างของมันก็เปลี่ยนไปบ้าง จากทรงกลมจะกลายเป็นรูปกรวย หลังจากผ่านไประยะหนึ่ง คนจรจัดจะยึดติดกับพื้นผิวเสมอโดยด้านที่มีความละเอียดอยู่ การก่อตัวและการเจริญเติบโตของก้านเริ่มขึ้นทันทีและมีการใช้เม็ดเล็ก ๆ ในการสร้างก้านซึ่งมีการแปลเป็นภาษาท้องถิ่นที่ปลายด้านหลังของคนจรจัด เมื่อลำต้นโตขึ้น ความหยาบจะหายไป หลังจากที่ก้านถึงลักษณะความยาวขั้นสุดท้ายของซูแทมเนีย การแบ่งส่วนต่อเนื่องกันอย่างรวดเร็วก็เริ่มต้นขึ้น ซึ่งนำไปสู่การก่อตัวของอาณานิคม การแบ่งส่วนเหล่านี้ทำขึ้นตามลำดับที่กำหนดไว้อย่างเคร่งครัด (รูปที่ 102)
เราจะไม่อาศัยรายละเอียดของกระบวนการนี้ ขอให้เราใส่ใจกับปรากฏการณ์ที่น่าสนใจต่อไปนี้เท่านั้น ในช่วงแรกของสัตว์จรจัด Zootamnia ในระหว่างการพัฒนาอาณานิคมในบุคคลที่ก่อตัวขึ้น pinnate และปากจะไม่ทำงาน การให้อาหารเริ่มขึ้นในภายหลังเมื่ออาณานิคมเล็กประกอบด้วยบุคคล 12-16 คนแล้ว ดังนั้นทุกขั้นตอนแรกของการพัฒนาอาณานิคมจึงดำเนินการโดยเฉพาะโดยเสียค่าใช้จ่ายสำรองที่เกิดขึ้นในร่างกายของ macrozoid ในระหว่างการเจริญเติบโตและการพัฒนาในอาณานิคมของแม่ มีความคล้ายคลึงกันอย่างปฏิเสธไม่ได้ระหว่างการพัฒนาของโซแทมเนียเร่ร่อนและการพัฒนาของไข่ในสัตว์หลายเซลล์ ความคล้ายคลึงกันนี้แสดงออกในความจริงที่ว่าการพัฒนาในทั้งสองกรณีดำเนินการโดยใช้เงินสำรองที่สะสมไว้ก่อนหน้านี้โดยไม่ต้องรับรู้อาหารจากสภาพแวดล้อมภายนอก
เมื่อศึกษา ciliates รอบ ciliates นั่ง คำถามเกิดขึ้น: รูปแบบของกระบวนการทางเพศลักษณะเฉพาะของ ciliates ดำเนินการอย่างไร - conjugation? ปรากฎว่าเมื่อเชื่อมต่อกับวิถีชีวิตประจำที่มีการเปลี่ยนแปลงที่สำคัญบางอย่าง ในช่วงเริ่มต้นของกระบวนการทางเพศ จะเกิดคนจรจัดพิเศษขนาดเล็กมากก่อตัวขึ้นในอาณานิคม เคลื่อนไหวอย่างแข็งขันด้วยความช่วยเหลือของกลีบตาพวกมันคลานไปตามอาณานิคมเป็นระยะเวลาหนึ่งจากนั้นจึงรวมเข้ากับบุคคลที่อยู่ประจำปกติขนาดใหญ่ของอาณานิคม ดังนั้น การแยกความแตกต่างของคอนจูกันต์ออกเป็นสองกลุ่มของบุคคลจึงเกิดขึ้นที่นี่: ขนาดเล็ก เคลื่อนที่ได้ (จุลภาค) และขนาดใหญ่กว่า ไม่เคลื่อนที่ (มาโครคอนจูแกนต์) การแยกผันของ conjugants ออกเป็นสองประเภท โดยหนึ่ง (microconjugants) เป็นแบบเคลื่อนที่ เป็นการปรับตัวที่จำเป็นให้เข้ากับวิถีชีวิตที่อยู่ประจำ หากปราศจากสิ่งนี้ กระบวนการทางเพศปกติ (conjugation) ก็ไม่สามารถรับรองได้
ดูดบุหรี่ (SUCTORIA)
กลุ่มที่แปลกประหลาดมากในแง่ของวิธีที่พวกเขากินเป็นตัวแทนของ ดูด ciliates(ซัคโทเรีย). สิ่งมีชีวิตเหล่านี้ เช่น suvoyka และ ciliated ciliates อื่นๆ เป็นสิ่งมีชีวิต จำนวนสปีชีส์ที่อยู่ในลำดับนี้วัดได้หลายโหล รูปร่างของ ciliates ที่ดูดนั้นมีความหลากหลายมาก ลักษณะเด่นบางชนิดแสดงในรูปที่ 103 บางชนิดนั่งบนลำต้นบนก้านที่ยาวมากหรือน้อย บางชนิดไม่มีก้าน ในบางกิ่งก้านค่อนข้างแข็งแรง ฯลฯ อย่างไรก็ตาม แม้ว่าจะมีรูปแบบที่หลากหลาย แต่ ciliates ดูดทั้งหมด มีลักษณะเด่น ๒ ประการ คือ
1) การขาดอย่างสมบูรณ์ (ในรูปแบบผู้ใหญ่) ของอุปกรณ์ปรับเลนส์
2) การปรากฏตัวของอวัยวะพิเศษ - หนวดที่ทำหน้าที่ดูดเหยื่อ

ในการดูด ciliates ประเภทต่างๆ จำนวนหนวดไม่เท่ากัน มักจะรวมตัวกันเป็นกลุ่ม ด้วยกล้องจุลทรรศน์กำลังขยายสูง จะเห็นได้ว่าที่ปลายหนวดมีสารเพิ่มความหนารูปไม้กระบอง
หนวดทำงานอย่างไร? ไม่ยากเลยที่จะตอบคำถามนี้โดยสังเกตการดูด ciliates เป็นระยะเวลาหนึ่ง หากโปรโตซัวตัวเล็ก (แฟลเจลเลท, อินฟิวโซเรีย) สัมผัสหนวดของซัคทอเรีย มันก็จะเกาะติดกับมันทันที ความพยายามทั้งหมดของเหยื่อที่จะแยกตัวออกไปมักจะไร้ประโยชน์ หากคุณยังคงสังเกตเหยื่อติดอยู่กับหนวด คุณจะเห็นว่ามันค่อยๆ เริ่มลดขนาดลง เนื้อหาจะค่อยๆ "สูบ" ผ่านหนวดเข้าไปในเอ็นโดพลาสซึมของซิลิเอตที่ดูดจนเหลือเพียงเม็ดเดียวจากเหยื่อซึ่งถูกทิ้งไป ดังนั้นหนวดของ ciliates ที่ดูดจึงมีความโดดเด่นอย่างสมบูรณ์ไม่มีที่ใดในโลกของสัตว์ที่เป็นอวัยวะสำหรับดักจับและในขณะเดียวกันก็ดูดอาหาร (รูปที่ 103)
การดูด ciliates เป็นสัตว์นักล่าที่ไม่เคลื่อนไหวซึ่งไม่ไล่ตามเหยื่อ แต่จับได้ทันทีหากเหยื่อประมาทสัมผัสตัวมันเอง

ทำไมเราถึงอ้างถึงสิ่งมีชีวิตที่แปลกประหลาดเหล่านี้กับคลาสของ ciliates? เมื่อมองแวบแรกพวกเขาไม่มีส่วนเกี่ยวข้องกับพวกเขา ข้อเท็จจริงต่อไปนี้พูดถึงการเป็นของ suctoria กับ ciliates ประการแรก พวกมันมีอุปกรณ์นิวเคลียร์ตามแบบฉบับของ ciliates ซึ่งประกอบด้วยมาโครนิวเคลียสและไมโครนิวเคลียส ประการที่สองในระหว่างการสืบพันธุ์พวกเขาพัฒนา cilia ที่ไม่มีอยู่ในบุคคลที่ "เป็นผู้ใหญ่" การสืบพันธุ์แบบไม่อาศัยเพศและในขณะเดียวกันการตั้งถิ่นฐานใหม่ของ ciliates ที่ดูดนั้นดำเนินการโดยการก่อตัวของคนจรจัดพร้อมกับกลีบเลี้ยงวงแหวนหลายซี่ การก่อตัวของคนจรจัดใน suctoria สามารถเกิดขึ้นได้หลายวิธี บางครั้งพวกมันเกิดขึ้นจากการแบ่งตัวที่ไม่สม่ำเสมอ (แตกหน่อ) ซึ่งไตแต่ละข้างที่แยกออกด้านนอกจะได้รับส่วนมาโครนิวเคลียสและหนึ่งไมโครนิวเคลียส (รูปที่ 104, L) ในมารดาคนหนึ่ง ตาของลูกสาวหลายคนสามารถเกิดขึ้นได้ในคราวเดียว (รูปที่ 104, 5) ในสายพันธุ์อื่น (รูปที่ 104, D, E) มีการสังเกตวิธีการ "แตกหน่อภายใน" ที่แปลกประหลาดมาก ในเวลาเดียวกันจะเกิดโพรงขึ้นภายในร่างกายของตัวดูดนมซึ่งมีการสร้างไตจรจัด มันออกมาทางรูพิเศษซึ่งมัน "บีบ" ด้วยความยากลำบากที่ทราบ
พัฒนาการของเอ็มบริโอภายในร่างกายของมารดา และจากนั้นการคลอดบุตร เป็นการเปรียบเทียบที่น่าสนใจของสิ่งที่ง่ายที่สุดกับสิ่งที่เกิดขึ้นในสิ่งมีชีวิตหลายเซลล์ที่สูงกว่า
ในหน้าที่แล้ว มีการพิจารณาตัวแทนที่อาศัยอยู่อย่างอิสระหลายคนของคลาส ciliates ซึ่งปรับให้เข้ากับสภาพแวดล้อมที่แตกต่างกัน เป็นเรื่องที่น่าสนใจที่จะเข้าหาประเด็นของการปรับ ciliates ให้เข้ากับสภาพความเป็นอยู่และในทางกลับกันเพื่อดูว่ามีลักษณะอย่างไร คุณสมบัติทั่วไป ciliates อาศัยอยู่ในสภาพแวดล้อมที่กำหนดไว้อย่างชัดเจน
ตัวอย่างเช่น ลองมาดูที่อยู่อาศัยที่แตกต่างกันอย่างมากสองแห่ง: ชีวิตในองค์ประกอบของแพลงก์ตอนและชีวิตที่อยู่ด้านล่างในความหนาของทราย
อาหารเสริมแพลงค์โทนิค
พบชนิด ciliate จำนวนมากในแพลงก์ตอนในทะเลและน้ำจืด
คุณสมบัติของการปรับตัวให้เข้ากับชีวิตในคอลัมน์น้ำนั้นเด่นชัดเป็นพิเศษในเรดิโอลาเรียน แนวหลักของการปรับตัวให้เข้ากับวิถีชีวิตของแพลงก์ตอนจะลดลงไปสู่การพัฒนาลักษณะโครงสร้างดังกล่าวที่นำไปสู่การทะยานของสิ่งมีชีวิตในคอลัมน์น้ำ

แพลงก์โทนิกทั่วไป ยิ่งกว่านั้น ตระกูล ciliates ในทะเลเกือบทั้งหมดคือ ตินตินนิดส์(Tintinnidae, รูปที่ 105, 5). จำนวนสปีชีส์ของทินทินนิดที่รู้จักกันทั้งหมดมีประมาณ 300 ชนิด สิ่งเหล่านี้เป็นรูปแบบขนาดเล็ก โดดเด่นด้วยความจริงที่ว่าร่างกายโปรโตพลาสซึมของ ciliate นั้นถูกวางไว้ในที่โล่งโปร่งแสงและในเวลาเดียวกันที่แข็งแรงประกอบด้วยอินทรียวัตถุ แผ่นดิสก์ยื่นออกมาจากบ้านโดยมีกลีบตาซึ่งเคลื่อนไหวกะพริบตลอดเวลา ในสถานะของการลอยตัว infusoria ในคอลัมน์น้ำนั้นส่วนใหญ่ได้รับการสนับสนุนโดยการทำงานอย่างต่อเนื่องของอุปกรณ์ปรับเลนส์ เห็นได้ชัดว่าบ้านทำหน้าที่ปกป้องร่างกายส่วนล่างของ ciliate ตินตินนิดเพียง 2 สายพันธุ์เท่านั้นที่อาศัยอยู่ในน้ำจืด (ไม่นับ 7 สายพันธุ์ที่มีลักษณะเฉพาะของทะเลสาบไบคาลเท่านั้น)
ciliates น้ำจืดมีการปรับตัวอื่น ๆ ให้กับชีวิตในแพลงก์ตอน ในหลาย ๆ ไซโตพลาสซึมนั้นถูกทำให้ร้อนมาก (Loxodes, Condylostoma, Trachelius) เพื่อให้ดูเหมือนโฟม สิ่งนี้นำไปสู่การลดความถ่วงจำเพาะอย่างมีนัยสำคัญ นอกจากนี้ ciliates ที่ระบุไว้ทั้งหมดยังมี ciliary cover ซึ่งร่างกายของ ciliate ในแง่ของความถ่วงจำเพาะมีขนาดใหญ่กว่าเพียงเล็กน้อยเท่านั้น แรงดึงดูดเฉพาะน้ำบำรุงรักษาง่ายในสภาวะ "ทะยาน" ในบางสปีชีส์ รูปร่างของร่างกายมีส่วนทำให้พื้นที่ผิวจำเพาะเพิ่มขึ้นและช่วยให้ลอยตัวในน้ำได้ง่ายขึ้น ตัวอย่างเช่น แพลงก์ตอน ciliates บางชนิดของทะเลสาบไบคาลมีลักษณะคล้ายร่มหรือร่มชูชีพ (Liliomorpha, รูปที่ 105, 2) มีแพลงก์โทนิกดูด ciliate หนึ่งตัวในทะเลสาบไบคาล (Mucophrya pelagica, รูปที่ 105, 4) ซึ่งแตกต่างอย่างมากจากญาตินั่งของมัน สายพันธุ์นี้ไม่มีลำต้น ตัวโปรโตพลาสซึมของมันถูกล้อมรอบด้วยปลอกเมือกกว้าง การปรับตัวที่นำไปสู่การลดน้ำหนัก หนวดเส้นบางยาวยื่นออกมา ซึ่งควบคู่ไปกับหน้าที่ตรงของพวกมัน อาจทำหน้าที่อีกอย่างหนึ่งด้วย - การเพิ่มพื้นที่ผิวจำเพาะซึ่งก่อให้เกิดการลอยตัวในน้ำ
สุดท้ายนี้ จำเป็นต้องพูดถึงอีกรูปแบบหนึ่ง กล่าวคือ รูปแบบทางอ้อมของการปรับตัวของ ciliates ให้เข้ากับชีวิตในแพลงก์ตอน นี่คือสิ่งที่แนบมาของ ciliates ขนาดเล็กกับสิ่งมีชีวิตอื่น ๆ ที่นำไปสู่วิถีชีวิตของแพลงก์ตอน ใช่ ท่ามกลาง ciliates ขลิบ(Peritricha) มีสปีชีส์ค่อนข้างมากที่เกาะติดกับโคปพอดของแพลงก์โทนิก นี่เป็นวิถีชีวิตปกติและปกติของ ciliates ประเภทนี้
พร้อมกับ ciliates circumciliary และในหมู่ ดูด(Suctoria) มีสปีชีส์ที่อาศัยอยู่บนแพลงตอน
พลเมืองที่อาศัยอยู่ในSand
หาดทรายและสันดอนเป็นที่อยู่อาศัยที่แปลกประหลาดอย่างยิ่ง ตามแนวชายฝั่งทะเล พวกเขาครอบครองพื้นที่กว้างใหญ่และมีลักษณะเป็นสัตว์ที่แปลกประหลาด
ดำเนินการเพื่อ ปีที่แล้วการศึกษาจำนวนมากในประเทศต่างๆ แสดงให้เห็นว่าทรายทะเลจำนวนมากอุดมไปด้วยสัตว์ที่มีขนาดเล็กมากหรือขนาดเล็กมาก ระหว่างอนุภาคทรายมีช่องว่างขนาดเล็กและเล็ก ๆ มากมายที่เต็มไปด้วยน้ำ ปรากฎว่าพื้นที่เหล่านี้เต็มไปด้วยสิ่งมีชีวิตที่เป็นของกลุ่มสัตว์โลกที่หลากหลายที่สุด ครัสเตเชียหลายสิบสายพันธุ์อาศัยอยู่ที่นี่ annelids, พยาธิตัวกลม, โดยเฉพาะหนอนตัวแบนจำนวนมาก, หอยบางตัว, coelenterates ในจำนวนมากยังมีโปรโตซัวซึ่งส่วนใหญ่เป็น ciliates ตามข้อมูลสมัยใหม่ สัตว์ ciliates ที่อาศัยอยู่ในความหนาของทรายทะเลมีประมาณ 250-300 สายพันธุ์ หากเราระลึกไว้เสมอว่าไม่เพียงแค่ ciliates เท่านั้น แต่ยังรวมถึงสิ่งมีชีวิตกลุ่มอื่น ๆ ที่อาศัยอยู่ในความหนาของทรายด้วยจำนวนรวมของสายพันธุ์ของพวกมันจะมีมาก สัตว์ทั้งชุดที่อาศัยอยู่ในความหนาของทราย อาศัยอยู่ในช่องว่างที่เล็กที่สุดระหว่างเม็ดทราย เรียกว่าสัตว์จำพวกสะเทินน้ำสะเทินบก
ความสมบูรณ์และองค์ประกอบของสปีชีส์ของสัตว์จำพวก psammophilic นั้นพิจารณาจากปัจจัยหลายประการ ในหมู่พวกเขาโดยเฉพาะ ความสำคัญมีขนาดเท่าเม็ดทราย ทรายหยาบมีสัตว์ที่น่าสงสาร บรรดาสัตว์ที่มีทรายปนทรายละเอียดมาก (ที่มีเส้นผ่านศูนย์กลางอนุภาคน้อยกว่า 0.1 มม.) ก็ไม่ดีเช่นกัน ซึ่งเห็นได้ชัดว่าช่องว่างระหว่างอนุภาคนั้นเล็กเกินไปสำหรับสัตว์ที่จะอาศัยอยู่ในนั้น ทรายที่ร่ำรวยที่สุดในชีวิตคือเม็ดทรายปานกลางและเม็ดละเอียด
ปัจจัยที่สองที่มีบทบาทสำคัญในการพัฒนาของสัตว์จำพวก psammophilic คือความสมบูรณ์ของทรายในซากอินทรีย์และการสลายตัวของสารอินทรีย์ (ระดับที่เรียกว่า saprobity) ทรายที่ปราศจากอินทรียวัตถุเป็นสิ่งที่น่าสงสารในชีวิต ในทางกลับกัน ทรายเกือบจะไร้ชีวิตชีวาและอุดมไปด้วยอินทรียวัตถุ เนื่องจากการสลายตัวของอินทรียวัตถุนำไปสู่การสูญเสียออกซิเจน บ่อยครั้งที่การหมักไฮโดรเจนซัลไฟด์แบบไม่ใช้ออกซิเจนจะถูกเพิ่มเข้าไป
การปรากฏตัวของไฮโดรเจนซัลไฟด์อิสระเป็นปัจจัยลบอย่างยิ่งที่ส่งผลต่อการพัฒนาของสัตว์
พืชที่มีสาหร่ายเซลล์เดียว (ไดอะตอม, เพอริดิเนียม) ค่อนข้างอุดมสมบูรณ์บางครั้งพัฒนาในชั้นผิวของทราย นี่เป็นปัจจัยที่สนับสนุนการพัฒนาของสัตว์จำพวก psammophilic เนื่องจากสัตว์ขนาดเล็กจำนวนมาก (รวมถึง ciliates) กินสาหร่าย
ในที่สุด ปัจจัยที่ส่งผลเสียอย่างมากต่อบรรดาสัตว์จำพวก psammophilic ก็คือการโต้คลื่น เป็นเรื่องที่เข้าใจได้ค่อนข้างมาก เนื่องจากคลื่นที่ซัดทับชั้นบนของทราย คร่าชีวิตสิ่งมีชีวิตทั้งหมดที่นี่ สัตว์จำพวก psammophilic เป็นสัตว์ที่ร่ำรวยที่สุดในอ่าวที่มีที่กำบังและมีความร้อนสูง การลดลงและการไหลไม่ได้ป้องกันการพัฒนาของสัตว์จำพวก psammophilic เมื่อน้ำออกจากน้ำชั่วคราวในเวลาน้ำลงเผยให้เห็นทรายจากนั้นในความหนาของทรายในช่วงเวลาระหว่างเม็ดทรายก็ยังคงอยู่และสิ่งนี้ไม่ได้ขัดขวางการดำรงอยู่ของสัตว์
ใน ciliates ซึ่งเป็นส่วนหนึ่งของสัตว์จำพวก psammophilous และอยู่ในกลุ่มระบบต่างๆ (คำสั่ง, ครอบครัว) ลักษณะทั่วไปหลายอย่างได้รับการพัฒนาในกระบวนการวิวัฒนาการซึ่งเป็นการปรับตัวให้เข้ากับสภาวะแปลกประหลาดของการดำรงอยู่ระหว่างอนุภาคทราย

รูปที่ 106 แสดงสัตว์บางชนิดของสัตว์จำพวกแมลงจำพวกซิลิเอตที่อยู่ในลำดับและวงศ์ที่แตกต่างกัน มีความคล้ายคลึงกันมากมายระหว่างพวกเขา ร่างกายส่วนใหญ่มีความยาวมากหรือน้อยเหมือนหนอน ทำให้ง่ายต่อการ "บีบ" ลงในรูที่เล็กที่สุดระหว่างเม็ดทราย ในหลาย ๆ สายพันธุ์ (รูปที่ 106) การยืดตัวของร่างกายจะรวมกับการแบน เครื่องมือปรับเลนส์ปรับเลนส์ได้รับการพัฒนามาอย่างดีเสมอซึ่งช่วยให้เคลื่อนไหวด้วยแรงบางอย่างในช่องว่างแคบ ๆ บ่อยครั้งที่ cilia พัฒนาที่ด้านหนึ่งของร่างกายที่แบนเหมือนหนอนและด้านตรงข้ามนั้นเปลือยเปล่า คุณลักษณะนี้น่าจะเกี่ยวข้องกับความสามารถ ซึ่งเด่นชัดในสปีชีส์ซามโมฟีลิกส่วนใหญ่ ในการติด (แนบ) กับสารตั้งต้นอย่างใกล้ชิดและแน่นหนามากผ่านเครื่องมือปรับเลนส์ (ปรากฏการณ์ที่เรียกว่า thigmotaxis) คุณสมบัตินี้ช่วยให้สัตว์อยู่ในสถานที่ในกรณีที่กระแสน้ำเกิดขึ้นในช่องว่างแคบ ๆ ที่พวกมันอาศัยอยู่ ในกรณีนี้ อาจเป็นประโยชน์มากกว่าสำหรับด้านตรงข้ามกับด้านที่สัตว์ยึดติดกับพื้นผิวเรียบ
psammophilic ciliates กินอะไร? ส่วนสำคัญของ "อาหาร" ในหลายสายพันธุ์คือสาหร่าย โดยเฉพาะไดอะตอม แบคทีเรียทำหน้าที่เป็นอาหารในระดับที่น้อยกว่า นอกจากนี้ยังขึ้นอยู่กับข้อเท็จจริงที่ว่าในทรายมีแบคทีเรียเพียงไม่กี่ตัวที่ไม่มีมลพิษอย่างหนัก ในที่สุด โดยเฉพาะอย่างยิ่งในหมู่ ciliates psammophilous ที่ใหญ่ที่สุด มีรูปแบบกินสัตว์อื่นจำนวนมากที่กิน ciliates อื่น ๆ ที่เป็นของสายพันธุ์ที่เล็กกว่า Psammophilic ciliates มีการกระจายทุกที่
อินฟูโซเรีย อะพอสโทเมทส์

ciliates spirophria(Spirophrya subparasitica) ในสภาวะที่กักขังมักจะพบได้นั่งบนก้านขนาดเล็กบนกุ้งทะเลขนาดเล็กที่มีแพลงก์โทนิก ในขณะที่ครัสเตเชียนแหวกว่ายอยู่ในน้ำทะเล สไปโรเฟรียที่นั่งอยู่บนนั้นจะไม่มีการเปลี่ยนแปลงใดๆ สำหรับการพัฒนาต่อไปของ ciliates จำเป็นต้องกินครัสเตเชียนโดยโพลิปไฮดรอยด์ทางทะเลซึ่งเกิดขึ้นค่อนข้างบ่อย (รูปที่ 107) ทันทีที่ซีสต์ของสาหร่ายเกลียวทองร่วมกับครัสเตเชียนเข้าสู่โพรงย่อยอาหาร ciliates ขนาดเล็กจะออกมาจากพวกเขาทันทีซึ่งเริ่มที่จะกินอย่างกระฉับกระเฉงบนสารละลายอาหารที่เกิดขึ้นจากการย่อยอาหารของครัสเตเชียนที่กลืนเข้าไป ภายในหนึ่งชั่วโมงขนาดของ infusoria จะเพิ่มขึ้น 3-4 เท่า อย่างไรก็ตาม การสืบพันธุ์ไม่เกิดขึ้นในขั้นตอนนี้ ก่อนที่เราจะเป็นขั้นตอนการเจริญเติบโตตามปกติของ ciliates ซึ่งเรียกว่า Trophont หลังจากนั้นครู่หนึ่งพร้อมกับเศษอาหารที่ไม่ได้แยกแยะ Trophont จะถูกโปลิปโยนลงไปในน้ำทะเล ที่นี่ว่ายน้ำอย่างแข็งขันมันลงมาตามร่างกายของโพลิปไปที่พื้นรองเท้าซึ่งติดอยู่กับตัวเองถูกล้อมรอบด้วยถุงน้ำ ขั้นตอนนี้ของ ciliate ขนาดใหญ่ที่ห่อหุ้มไว้บนติ่งเนื้อเรียกว่า tomont นี่คือระยะการผสมพันธุ์ Tomont ไม่กิน แต่แบ่งอย่างรวดเร็วหลายครั้งติดต่อกัน (รูปที่ 107, 7) ผลที่ได้คือกลุ่มของ ciliates ที่เล็กมากทั้งกลุ่ม จำนวนของพวกเขาขึ้นอยู่กับขนาดของ tomont ซึ่งจะถูกกำหนดโดยขนาดของ trophont ที่ให้กำเนิดของมัน ciliates ขนาดเล็กที่เกิดขึ้นจากการแบ่ง tomont (เรียกว่า tomites หรือ vagrants) เป็นตัวแทนของขั้นตอนการตั้งถิ่นฐาน
พวกเขาออกจากถุงว่ายน้ำอย่างรวดเร็ว (โดยไม่ต้องกินในเวลาเดียวกัน แต่ใช้สำรองที่มีอยู่ในไซโตพลาสซึม) หากพวกเขา "โชคดี" ที่เจอโคพพอด พวกมันจะยึดติดอยู่กับมันและปิดล้อมทันที นี่คือระยะที่เราเริ่มพิจารณาวัฏจักรของเรา
ในการตรวจสอบ วงจรชีวิตสาหร่ายเกลียวทองดึงความสนใจไปที่การแบ่งระยะที่ชัดเจนซึ่งมีความสำคัญทางชีวภาพที่แตกต่างกัน โทรพรเป็นช่วงการเจริญเติบโต มันเติบโตสะสมไซโตพลาสซึมจำนวนมากและสารสำรองทุกชนิดเนื่องจากโภชนาการที่แข็งแรงและรวดเร็ว โทรฟอนต์ไม่สามารถสืบพันธุ์ได้ ปรากฏการณ์ย้อนกลับถูกพบใน tomont - ไม่สามารถให้อาหารและการสืบพันธุ์ที่รวดเร็ว หลังจากแต่ละแผนก ไม่มีการเติบโต ดังนั้นการสืบพันธุ์ของ tomont จึงลดลงอย่างรวดเร็วจนกลายเป็นคนจรจัดจำนวนมาก ในที่สุด คนเร่ร่อนก็ทำหน้าที่พิเศษและมีลักษณะเฉพาะของพวกเขาเท่านั้น: พวกเขาเป็นบุคคล - ผู้ตั้งถิ่นฐานและผู้จัดจำหน่ายของสายพันธุ์ พวกเขาไม่สามารถกินหรือขยายพันธุ์ได้
วัฏจักรชีวิตของอิคธิโอฟทีเรียส

เมื่อสิ้นสุดระยะเวลาการเจริญเติบโต ichthyophthirius จะมีขนาดที่ใหญ่มากเมื่อเทียบกับคนจรจัด: เส้นผ่านศูนย์กลาง 0.5-1 มม. เมื่อถึงค่าที่จำกัด ciliates จะเคลื่อนออกจากเนื้อเยื่อของปลาลงไปในน้ำอย่างแข็งขันและว่ายอย่างช้าๆ เป็นระยะเวลาหนึ่งด้วยความช่วยเหลือของเครื่องมือปรับเลนส์ที่ครอบคลุมทั้งตัว ในไม่ช้า ichthyophthirius ขนาดใหญ่จะเกาะติดกับวัตถุใต้น้ำและหลั่งถุงน้ำออก ทันทีหลังจาก encysting การแบ่ง ciliates ที่ต่อเนื่องกันเริ่มต้น: ครึ่งแรกจากนั้นลูกสาวแต่ละคนจะถูกแบ่งออกเป็นสองส่วนอีกครั้งและอื่น ๆ มากถึง 10-11 ครั้ง เป็นผลให้มีบุคคลขนาดเล็กเกือบ 2,000 คนที่มีขนปกคลุมอยู่ภายในถุงน้ำ ภายในถุงน้ำ คนเร่ร่อนกำลังเคลื่อนไหวอย่างแข็งขัน พวกเขาเจาะเปลือกและออกมา คนจรจัดว่ายน้ำอย่างแข็งขันติดเชื้อปลาใหม่
อัตราการแบ่งตัวของ ichthyophthirius ในซีสต์รวมถึงอัตราการเจริญเติบโตในเนื้อเยื่อของปลานั้นขึ้นอยู่กับอุณหภูมิในระดับสูง จากการศึกษาของผู้เขียนหลายคน ให้ตัวเลขต่อไปนี้: ที่อุณหภูมิ 26-27°C การพัฒนาคนจรจัดในถุงน้ำจะใช้เวลา 10-12 ชั่วโมง ที่อุณหภูมิ 15-16°C ใช้เวลา 28-30 ชั่วโมง ที่ 4-5 °C อยู่ได้นาน 6 -7 วัน
การต่อสู้กับ ichthyophthirius นำเสนอปัญหาที่สำคัญ สิ่งสำคัญอันดับแรกที่นี่คือมาตรการป้องกันที่มุ่งป้องกันไม่ให้คนเร่ร่อนว่ายน้ำอิสระเจาะเนื้อเยื่อของปลา ในการทำเช่นนี้จะเป็นประโยชน์ในการย้ายปลาที่ป่วยไปยังอ่างเก็บน้ำหรือพิพิธภัณฑ์สัตว์น้ำใหม่ ๆ เพื่อสร้างสภาวะการไหลซึ่งมีประสิทธิภาพโดยเฉพาะอย่างยิ่งในการต่อสู้กับ ichthyophthirius
ciliates ไตรโคดีนส์

ระบบทั้งหมดของการปรับตัวของไตรโคดินให้มีชีวิตบนพื้นผิวของโฮสต์นั้นมีจุดมุ่งหมายที่จะไม่แตกออกจากร่างกายของโฮสต์ (ซึ่งเกือบจะเท่ากับความตาย) ในขณะที่ยังคงความคล่องตัว อุปกรณ์เหล่านี้สมบูรณ์แบบมาก ร่างกายของไตรโคดีนาส่วนใหญ่อยู่ในรูปของจานที่ค่อนข้างแบน บางครั้งก็เป็นหมวก ด้านที่หันไปทางลำตัวของโฮสต์จะเว้าเล็กน้อยมันเป็นตัวดูดสิ่งที่แนบมา ตามขอบด้านนอกของตัวดูดมีกลีบของตาที่พัฒนามาอย่างดีด้วยความช่วยเหลือซึ่งการเคลื่อนไหว (คลาน) ของ ciliates ส่วนใหญ่เกิดขึ้นบนพื้นผิวของตัวปลา กลีบนี้สอดคล้องกับกลีบดอกไม้ที่พบในคนจรจัด ciliate รอบนั่งที่กล่าวถึงข้างต้น ดังนั้นจึงสามารถเปรียบเทียบ tricodina กับคนจรจัดได้ บนพื้นผิวหน้าท้อง (บนเครื่องดูด) ไตรโคดินมีอุปกรณ์รองรับและสิ่งที่แนบมาที่ซับซ้อนมาก ซึ่งช่วยให้ ciliates อยู่บนโฮสต์ โดยไม่ต้องลงรายละเอียดเกี่ยวกับโครงสร้าง เราสังเกตว่าพื้นฐานของมันคือวงแหวนที่มีโครงร่างที่ซับซ้อน ซึ่งประกอบด้วยส่วนที่แยกจากกันซึ่งมีฟันภายนอกและภายใน (รูปที่ 109, B) แหวนนี้ก่อให้เกิดความยืดหยุ่นและเป็นพื้นฐานที่แข็งแกร่งของพื้นผิวหน้าท้องซึ่งทำหน้าที่เป็นตัวดูด ไตรโคดินประเภทต่าง ๆ แตกต่างกันไปตามจำนวนของส่วนที่ประกอบเป็นวงแหวน และในการกำหนดค่าของขอเกี่ยวด้านนอกและด้านใน
ที่ด้านข้างของลำตัวของ Trichodina ตรงข้ามกับดิสก์มีเครื่องมือแบบพินเนทและช่องปาก โครงสร้างของมันเป็นแบบอย่างของ .ไม่มากก็น้อย ciliates ขลิบ. เยื่อ Adoral บิดตามเข็มนาฬิกานำไปสู่ช่องด้านล่างซึ่งเป็นปาก อุปกรณ์นิวเคลียร์ของทริโคดินมักจะถูกจัดเรียงสำหรับ ciliates: หนึ่งมาโครนิวเคลียสที่เหมือนริบบิ้นและหนึ่งไมโครนิวเคลียสที่อยู่ติดกัน มีแวคิวโอลหดตัวหนึ่งอัน
Trichodins มีการกระจายอย่างกว้างขวางในอ่างเก็บน้ำทุกประเภท โดยเฉพาะอย่างยิ่งมักพบในลูกปลาหลายสายพันธุ์ ด้วยการสืบพันธุ์จำนวนมากจะใช้ไตรโคดิน อันตรายมากปลาโดยเฉพาะอย่างยิ่งถ้าฝูงครอบคลุมเหงือก สิ่งนี้รบกวนการหายใจตามปกติของปลา
เพื่อทำความสะอาดปลาของ Trichodin ขอแนะนำให้อาบน้ำบำบัดด้วยสารละลายโซเดียมคลอไรด์ 2% หรือสารละลายโพแทสเซียมเปอร์แมงกาเนต 0.01% (สำหรับทอด - เป็นเวลา 10-20 นาที)
อวัยวะภายในลำไส้ของอูงเกท
จากแผลเป็นผ่านตาข่าย อาหารจะถูกเรอเข้าไปในช่องปาก และเคี้ยวเพิ่มเติม (หมากฝรั่ง) มวลอาหารที่เคี้ยวแล้วกลืนเข้าไปอีกครั้งผ่านท่อพิเศษที่เกิดจากรอยพับของหลอดอาหารไม่ไปที่แผลเป็นอีกต่อไป แต่ไปที่หนังสือและจากที่นั่นไปยังอะโบมาซัมซึ่งสัมผัสกับน้ำย่อยของสัตว์เคี้ยวเอื้อง ในตับภายใต้เงื่อนไขของปฏิกิริยาที่เป็นกรดและการปรากฏตัวของเอนไซม์ย่อยอาหาร ciliates ตาย การเคี้ยวหมากฝรั่งจะถูกย่อย
จำนวนโปรโตซัวในกระเพาะรูเมน (เช่นเดียวกับในตาข่าย) สามารถเข้าถึงค่ามหาศาลได้ หากคุณหยดเนื้อหาของแผลเป็นแล้วตรวจดูด้วยกล้องจุลทรรศน์ (เมื่อได้รับความร้อน เนื่องจาก ciliates หยุดที่อุณหภูมิห้อง) แสดงว่า ciliates เป็นกลุ่มก้อนในมุมมอง เป็นเรื่องยากแม้แต่ในวัฒนธรรมที่จะได้รับ ciliates จำนวนมาก จำนวน ciliates ใน 1 cm3 ของเนื้อหาของรอยแผลเป็นถึงล้านและมักจะมากขึ้น ในแง่ของปริมาณรอยแผลเป็นทั้งหมด นี่ทำให้ตัวเลขทางดาราศาสตร์อย่างแท้จริง! ความสมบูรณ์ของอาหารในกระเพาะรูเมนที่มี ciliates นั้นขึ้นอยู่กับธรรมชาติของอาหารสัตว์เคี้ยวเอื้องเป็นอย่างมาก หากอาหารอุดมด้วยไฟเบอร์และมีคาร์โบไฮเดรตและโปรตีนต่ำ (หญ้า ฟาง) กระเพาะกระเพาะปลาจะมี ciliates ค่อนข้างน้อย เมื่อเพิ่มคาร์โบไฮเดรตและโปรตีน (รำ) ลงในอาหาร จำนวน ciliates จะเพิ่มขึ้นอย่างมากและถึงจำนวนมหาศาล ต้องระลึกไว้เสมอว่ามี ciliates ไหลออกอย่างต่อเนื่อง เมื่อรวมกับหมากฝรั่งเข้าไปใน abomasum พวกเขาตาย ระดับสูงจำนวน ciliates นั้นคงอยู่โดยการสืบพันธุ์ที่แข็งแรง
กีบเท้าคี่ (ม้า ลา ม้าลาย) ยังมี ciliates จำนวนมากในทางเดินอาหาร แต่การแปลเป็นภาษาท้องถิ่นในโฮสต์นั้นแตกต่างกัน กีบเท้าคี่ไม่มีกระเพาะที่ซับซ้อนเนื่องจากไม่มีความเป็นไปได้ในการพัฒนาโปรโตซัวในส่วนหน้าของทางเดินอาหาร แต่ในสัตว์น้ำ Equid ลำไส้ใหญ่และซีคัมได้รับการพัฒนาเป็นอย่างดี ซึ่งมักจะอุดตันด้วยมวลอาหารและมีบทบาทสำคัญในการย่อยอาหาร ในส่วนนี้ของลำไส้ เช่นเดียวกับในกระเพาะรูเมนและตาข่ายของสัตว์เคี้ยวเอื้อง โพรโตซัวที่อุดมสมบูรณ์มากพัฒนา ส่วนใหญ่เป็น ciliates ซึ่งส่วนใหญ่เป็นของเอนโดดินิโอมอร์ฟ อย่างไรก็ตามในแง่ขององค์ประกอบของสปีชีส์ สัตว์ในกระเพาะลำไส้ของสัตว์เคี้ยวเอื้องและสัตว์ในลำไส้ของสัตว์เคี้ยวเอื้องไม่ตรงกัน
พลเมืองของลำไส้ของสัตว์เคี้ยวเอื้อง
สิ่งที่น่าสนใจที่สุดคือ ciliates ตระกูล ofrioscolicid(Ophryoscolecidae) เกี่ยวข้องกับ สั่งเอ็นโดนิโอมอร์ฟ. ลักษณะเฉพาะของการปลดนี้คือไม่มีฝาครอบเลนส์ปรับเลนส์ต่อเนื่อง การสร้างเลนส์ปรับเลนส์ที่ซับซ้อน - cirres - ตั้งอยู่ที่ส่วนหน้าของร่างกาย ciliates ในบริเวณที่เปิดปาก สำหรับองค์ประกอบพื้นฐานของเครื่องมือปรับเลนส์เลนส์ คุณสามารถเพิ่มกลุ่มของตับแข็งเพิ่มเติม ซึ่งอยู่ที่ส่วนหน้าหรือส่วนหลังของร่างกาย จำนวนสปีชีส์ของ ciliates ของตระกูล ofrioscolecid ทั้งหมดประมาณ 120 ชนิด

รูปที่ 110 แสดง ofrioscolecid ทั่วไปบางส่วนจากกระเพาะรูเมนของสัตว์เคี้ยวเอื้อง ciliates ของสกุล Entodinium (Entodinium, รูปที่ 110, L) ส่วนใหญ่จัดเรียงอย่างเรียบง่าย ที่ส่วนหน้าของร่างกายมีโซนตับแข็งหนึ่งโซน ส่วนหน้าของร่างกายซึ่งเปิดปากอยู่สามารถดึงเข้าด้านในได้ เอ็กโทพลาสซึมและเอนโดพลาสซึมถูกแบ่งเขตอย่างชัดเจน ท่อทวารหนักมองเห็นได้ชัดเจนที่ปลายด้านหลัง ซึ่งทำหน้าที่กำจัดเศษอาหารที่ไม่ได้ย่อย โครงสร้างค่อนข้างซับซ้อน anoplodynia(Anoplodinium, รูปที่ 110, B). พวกมันมีสองโซนของอุปกรณ์ปรับเลนส์ - perioral cirrhi และ dorsal cirrhi ทั้งสองตั้งอยู่ที่ส่วนหน้า ที่ส่วนท้ายของลำตัวของสปีชีส์ที่แสดงในรูป มีผลพลอยได้ที่แหลมยาว ซึ่งเป็นเรื่องปกติสำหรับโอริโอสโคเลซิดหลายสายพันธุ์ ได้รับการแนะนำว่าผลพลอยได้เหล่านี้มีส่วนทำให้เกิด "การผลัก" ของ ciliates ระหว่างอนุภาคของพืชที่เติมรอยแผลเป็น
ชนิด สกุล Eudiplodynia(Eudiplodinium, รูปที่ 110, B) คล้ายกับ anoplodyniaแต่ต่างจากพวกมันตรงที่มีแผ่นฐานโครงกระดูกอยู่ที่ขอบด้านขวาตามแนวคอหอย แผ่นโครงร่างนี้ประกอบด้วยสารที่คล้ายคลึงกันใน ลักษณะทางเคมีสู่เส้นใย กล่าวคือ สารที่ประกอบเป็นเปลือกของเซลล์พืช
ที่ สกุลโพลิพลาสตรอน(Polyplastron, รูปที่ 110, D, E) มีภาวะแทรกซ้อนเพิ่มเติมของโครงกระดูก โครงสร้างของ ciliates เหล่านี้อยู่ใกล้กับ eudiplodynia ความแตกต่างหลักๆ มาจากข้อเท็จจริงที่ว่าแทนที่จะมีแผ่นโครงกระดูกเพียงแผ่นเดียว ciliates เหล่านี้มีห้าแผ่น สองอันที่ใหญ่ที่สุดตั้งอยู่ทางด้านขวาและอีกสามอันที่เล็กกว่าอยู่ทางด้านซ้ายของ ciliate ลักษณะที่สองของโพลิพลาสตรอนคือการเพิ่มจำนวนของแวคิวโอลที่หดตัว Entodynia มีแวคิวโอลที่หดตัวหนึ่งอัน anoplodynia และ eudiplodynia มี vacuoles ที่หดตัวสองตัวและ polyplastron มีประมาณหนึ่งโหล
ที่ เอพิดิเนียม(Epidinium, รูปที่ 110) ซึ่งมีโครงกระดูกคาร์โบไฮเดรตที่ได้รับการพัฒนามาอย่างดีซึ่งอยู่ทางด้านขวาของร่างกาย บริเวณหลังของตับแข็งจะเปลี่ยนจากส่วนหน้าไปด้านหลัง หนามมักจะพัฒนาที่ส่วนหลังของ ciliates ของสกุลนี้
โครงสร้างที่ซับซ้อนที่สุดเผยให้เห็น สกุล Ofrioscolex(Ophryoscolex) หลังจากนั้นตั้งชื่อทั้งตระกูล ciliates (รูปที่ 110, E) พวกเขามีโซนหลังของตับแข็งที่พัฒนามาอย่างดีซึ่งครอบคลุมประมาณ 2/3 ของเส้นรอบวงของร่างกายและแผ่นโครงร่าง กระดูกสันหลังจำนวนมากก่อตัวขึ้นที่ปลายด้านหลังซึ่งมักจะยาวเป็นพิเศษ
ทำความคุ้นเคยกับตัวแทนทั่วไป ofrioscolecidแสดงให้เห็นว่าภายในครอบครัวนี้มีภาวะแทรกซ้อนที่สำคัญขององค์กร (จากเอนโทไดเนียถึงโอริโอสโคลเล็กซ์)
นอกจาก ciliates ตระกูล ofrioscolicidในกระเพาะของสัตว์เคี้ยวเอื้องพบตัวแทนที่เรารู้จักแล้วในปริมาณเล็กน้อย การแยกตัวของ ciliates isociliary. พวกมันมีสปีชีส์จำนวนน้อย ร่างกายของพวกเขาถูกปกคลุมอย่างสม่ำเสมอด้วย cilia แถวตามยาวไม่มีองค์ประกอบของโครงกระดูก ในมวลรวมของประชากร ciliate ของกระเพาะรูเมน พวกเขาไม่ได้มีบทบาทสำคัญ ดังนั้นเราจะไม่อาศัยอยู่ที่นี่
ciliates ofrioscolecides กินอะไรและอย่างไร? ปัญหานี้ได้รับการศึกษาอย่างละเอียดโดยนักวิทยาศาสตร์หลายคน โดยเฉพาะอย่างยิ่งในรายละเอียดโดย Professor V. A. Dogel

อาหาร Ofrioscolecid ค่อนข้างหลากหลายและมีความเชี่ยวชาญเฉพาะทางในสายพันธุ์ต่างๆ สายพันธุ์ที่เล็กที่สุดของสกุล Entodynia กินแบคทีเรีย เมล็ดแป้ง เชื้อรา และอนุภาคขนาดเล็กอื่นๆ ofrioscolecids ขนาดกลางและขนาดใหญ่จำนวนมากดูดซับอนุภาคของเนื้อเยื่อพืชซึ่งประกอบขึ้นเป็นกลุ่มของเนื้อหาในกระเพาะรูเมน เอ็นโดพลาสซึมของบางชนิดอุดตันด้วยอนุภาคของพืชอย่างแท้จริง คุณจะเห็นได้ว่า ciliates กระโจนเข้าหาเศษกระดาษทิชชู่ของพืช ฉีกมันเป็นชิ้น ๆ แล้วกลืนเข้าไป มักจะบิดเป็นเกลียวในร่างกายของมัน (รูปที่ 111, 4) บางครั้งจำเป็นต้องสังเกตภาพดังกล่าว (รูปที่ 111, 2) เมื่อร่างกายของ ciliate นั้นมีรูปร่างผิดปกติเนื่องจากการกลืนอนุภาคขนาดใหญ่
ใน ofrioscolecid บางครั้งมีการสังเกตการปล้นสะดม สายพันธุ์ที่ใหญ่กว่ากินสัตว์ที่เล็กกว่า การปล้นสะดม (รูปที่ 112) รวมกับความสามารถของสายพันธุ์เดียวกันในการกินอนุภาคของพืช

ciliates เข้าสู่กระเพาะของสัตว์เคี้ยวเอื้องได้อย่างไร? เส้นทางของการติดเชื้อ ofrioscolecides คืออะไร? ปรากฎว่าสัตว์เคี้ยวเอื้องแรกเกิดยังไม่มี ciliates ในกระเพาะรูเมน พวกมันจะหายไปในขณะที่สัตว์กำลังกินนม แต่ทันทีที่สัตว์เคี้ยวเอื้องเปลี่ยนเป็นอาหารจากพืช ciliates จะปรากฏในแผลเป็นและตาข่ายทันที ซึ่งจำนวนดังกล่าวก็เพิ่มขึ้นอย่างรวดเร็ว พวกเขามาจากที่ไหน? เป็นเวลานาน สันนิษฐานว่า ciliates ในกระเพาะรูเมนก่อให้เกิดระยะพักบางประเภท (ส่วนใหญ่คือซีสต์) ซึ่งกระจายตัวอย่างกว้างขวางในธรรมชาติ และเมื่อกลืนเข้าไป จะทำให้เกิดระยะการทำงานของ ciliates การศึกษาเพิ่มเติมพบว่า ciliates สัตว์เคี้ยวเอื้องไม่มีระยะพัก เป็นไปได้ที่จะพิสูจน์ว่าการติดเชื้อเกิดขึ้นกับ ciliates เคลื่อนที่ที่เจาะเข้าไปในช่องปากเมื่อเรอ หากคุณตรวจสอบภายใต้กล้องจุลทรรศน์ที่นำมาจาก ช่องปากเคี้ยวหมากฝรั่ง แต่ก็มี ciliates ลอยอยู่จำนวนมากเสมอ รูปแบบที่ใช้งานเหล่านี้สามารถเจาะเข้าไปในปากได้ง่ายและเข้าไปในกระเพาะของสัตว์เคี้ยวเอื้องอื่น ๆ จากภาชนะทั่วไปสำหรับดื่มพร้อมกับหญ้า หญ้าแห้ง (ซึ่งอาจมีน้ำลายที่มี ciliates) ฯลฯ เส้นทางของการติดเชื้อนี้ได้รับการพิสูจน์แล้วจากการทดลอง
หากไม่มีระยะพักใน ofrioscolecid แสดงว่าสามารถหาสัตว์ที่ "ไม่ผสมสารกันเสีย" ได้โดยง่ายโดยแยกพวกมันออกเมื่อพวกมันยังกินนมอยู่ หากหลีกเลี่ยงการสัมผัสโดยตรงระหว่างการเจริญเติบโตของสัตว์เคี้ยวเอื้องและสัตว์เคี้ยวเอื้องที่มีขนอ่อน สัตว์เล็กอาจถูกปล่อยทิ้งไว้โดยไม่มี ciliates ในกระเพาะรูเมน การทดลองดังกล่าวดำเนินการโดยนักวิทยาศาสตร์หลายคนในประเทศต่างๆ ผลลัพธ์ก็ชัดเจน ในกรณีที่ไม่มีการติดต่อระหว่างลูกอ่อน (นำมาจากแม่ในช่วงที่ป้อนนม) และสัตว์เคี้ยวเอื้องที่มี ciliates ในกระเพาะรูเมน สัตว์จะเติบโตเป็นหมันเมื่อเทียบกับ ciliates อย่างไรก็ตาม แม้แต่การสัมผัสสัตว์ที่มี ciliates ในระยะสั้น (อาหารทั่วไป ถังดื่มทั่วไป ทุ่งหญ้าทั่วไป) ก็เพียงพอแล้วที่ ciliates จะปรากฏในกระเพาะรูเมนของสัตว์ปลอดเชื้อ
ข้างต้นเป็นผลจากการทดลองเกี่ยวกับเนื้อหาของสัตว์เคี้ยวเอื้องที่ไม่มี ciliates ในกระเพาะรูเมนและตาข่าย สิ่งนี้ประสบความสำเร็จดังที่เราได้เห็นโดยการแยกตัวของคนหนุ่มสาวตั้งแต่เนิ่นๆ ทำการทดลองกับแกะและแพะ
ด้วยวิธีนี้ จึงเป็นไปได้ที่จะทำการสังเกตสัตว์ที่ "ไม่มีเชื้อ" เป็นระยะเวลาที่สำคัญ (มากกว่าหนึ่งปี) การขาด ciliates ในกระเพาะรูเมนส่งผลต่อชีวิตของโฮสต์อย่างไร? การขาด ciliates ส่งผลกระทบต่อโฮสต์ในทางลบหรือทางบวกหรือไม่? เพื่อตอบคำถามนี้ ได้ทำการทดลองกับแพะดังต่อไปนี้ เด็กแฝด (เพศเดียวกันและครอกเดียวกัน) ถูกพามาเพื่อให้มีเนื้อหาที่คล้ายคลึงกันมากขึ้น จากนั้นฝาแฝดตัวหนึ่งของคู่นี้ถูกเลี้ยงดูมาโดยไม่มี ciliates ในกระเพาะรูเมน (การแยกตัวตั้งแต่เนิ่นๆ) ในขณะที่อีกคู่หนึ่งจากจุดเริ่มต้นการกินอาหารจากพืชติดเชื้อ ciliates หลายประเภทอย่างล้นเหลือ ทั้งสองได้รับอาหารเหมือนกันทุกประการและถูกเลี้ยงดูมาในสภาวะเดียวกัน ความแตกต่างเพียงอย่างเดียวระหว่างพวกเขาคือการมีหรือไม่มี ciliates ในเด็กหลายคู่ที่ศึกษาในลักษณะนี้ ไม่พบความแตกต่างในการพัฒนาสมาชิกของทั้งสองในแต่ละคู่ ("infusor" และ "non-infusor") ดังนั้นจึงเป็นที่ถกเถียงกันอยู่ว่า ciliates ที่อาศัยอยู่ในกระเพาะรูเมนและตาข่ายไม่ได้มีผลอย่างมากต่อการทำงานที่สำคัญของสัตว์ที่เป็นโฮสต์
ผลการทดลองข้างต้นไม่อนุญาตให้ยืนยันว่า ciliates ของกระเพาะรูเมนไม่สนใจเจ้าของอย่างสมบูรณ์ การทดลองเหล่านี้ดำเนินการด้วยการรับประทานอาหารตามปกติของเจ้าบ้าน เป็นไปได้ว่าภายใต้เงื่อนไขอื่นๆ ด้วยอาหารที่แตกต่างกัน (เช่น การให้อาหารไม่เพียงพอ) จะสามารถเปิดเผยผลกระทบต่อโฮสต์ของสัตว์ประจำถิ่นของโปรโตซัวที่อาศัยอยู่ในกระเพาะรูเมนได้
มีข้อเสนอแนะหลายประการในวรรณคดีเกี่ยวกับอิทธิพลเชิงบวกที่เป็นไปได้ของสัตว์โปรโตซัวในกระเพาะรูเมนต่อกระบวนการย่อยอาหารของโฮสต์ มีการชี้ให้เห็นว่า ciliates หลายล้านตัวที่ว่ายน้ำอย่างแข็งขันในกระเพาะรูเมนและบดเนื้อเยื่อพืชมีส่วนทำให้เกิดการหมักและการย่อยอาหารจำนวนมากที่อยู่ในส่วนหน้าของทางเดินอาหาร ciliates จำนวนมากที่เข้าสู่ abomasum พร้อมกับหมากฝรั่งจะถูกย่อยและโปรตีนซึ่งเป็นส่วนสำคัญของร่างกายของ ciliates จะถูกดูดซึม Infusoria สามารถเป็นแหล่งโปรตีนเพิ่มเติมสำหรับโฮสต์ นอกจากนี้ยังมีข้อเสนอแนะว่า ciliates มีส่วนช่วยในการย่อยเส้นใยซึ่งประกอบเป็นอาหารสัตว์เคี้ยวเอื้องจำนวนมากและถ่ายโอนไปยังสถานะที่ย่อยได้ง่ายกว่า
สมมติฐานทั้งหมดเหล่านี้ไม่ได้รับการพิสูจน์ และมีการคัดค้านบางข้อ ตัวอย่างเช่น มีการชี้ให้เห็นว่า ciliates สร้างโปรโตพลาสซึมของร่างกายจากโปรตีนที่เข้าสู่กระเพาะรูเมนด้วยอาหารของโฮสต์ การดูดซับโปรตีนจากพืชจะเปลี่ยนเป็นโปรตีนจากสัตว์ในร่างกายซึ่งจะถูกย่อยในอะโบมาซัม สิ่งนี้ให้ประโยชน์แก่โฮสต์หรือไม่นั้นยังไม่ชัดเจน ประเด็นเหล่านี้เป็นประโยชน์อย่างยิ่ง เนื่องจากเรากำลังพูดถึงการย่อยอาหารสัตว์เคี้ยวเอื้อง ซึ่งเป็นเป้าหมายหลักของการเลี้ยงสัตว์ การวิจัยเพิ่มเติมเกี่ยวกับบทบาทของกระเพาะรูเมน ciliates ในการย่อยอาหารสัตว์เคี้ยวเอื้องเป็นที่ต้องการอย่างมาก
Ofrioscolecides ของสัตว์เคี้ยวเอื้องมีความจำเพาะที่กว้าง ในแง่ของสายพันธุ์ จำนวนประชากรของรอยแผลเป็นและอวนของวัวควาย แกะ และแพะนั้นอยู่ใกล้กันมาก หากเราเปรียบเทียบองค์ประกอบสปีชีส์ของกระเพาะรูเมนของแอนทีโลปในแอฟริกากับวัวควาย ที่นี่เช่นกัน ประมาณ 40% ของจำนวนสปีชีส์ทั้งหมดจะเป็นเรื่องธรรมดา อย่างไรก็ตาม มีโอริโอสโคเลซิดหลายสายพันธุ์ที่พบได้เฉพาะในแอนทีโลปหรือกวางเท่านั้น ดังนั้นเมื่อเทียบกับพื้นหลังของความจำเพาะในวงกว้างทั่วไปของ ofrioscolecides เราสามารถพูดถึงประเภทที่แยกจากกันและเฉพาะเจาะจงมากขึ้น
Infusoria ของลำไส้ของ equids
ตอนนี้ให้เรามาทำความรู้จักกับ ciliates ที่อาศัยอยู่ในถ้ำขนาดใหญ่และในช่องท้อง
ในแง่ของสปีชีส์ สัตว์ชนิดนี้ก็มีความหลากหลายมากเช่นเดียวกันกับสัตว์ในกระเพาะรูเมนของสัตว์เคี้ยวเอื้อง ปัจจุบันมี ciliates ประมาณ 100 สายพันธุ์ที่อาศัยอยู่ในลำไส้ใหญ่ของสัตว์ในตระกูลม้า ciliates ที่พบในนี้ในแง่ของการเป็นของกลุ่มระบบที่แตกต่างกันมีความหลากหลายมากกว่า ciliates ของกระเพาะรูเมนของสัตว์เคี้ยวเอื้อง

ในลำไส้ของม้ามี ciliates ค่อนข้างสองสามชนิดที่อยู่ในลำดับของ isociliary นั่นคือ ciliates ซึ่งเครื่องมือปรับเลนส์ไม่ก่อตัวเป็นพังผืดหรือตับแข็งใกล้กับบริเวณช่องปาก (รูปที่ 113, 1)
สั่งซื้อเอนโทไดนิโอมอร์ฟ(Entodiniomorpha) ยังเป็นตัวแทนอย่างมั่งคั่งในลำไส้ของม้า ในขณะที่มีเพียงตระกูลเดียวของ endodiniomorphs (ofrioscolecid family) ที่พบในกระเพาะของสัตว์เคี้ยวเอื้องตัวแทนของสามตระกูลอาศัยอยู่ในลำไส้ของม้า แต่เราจะไม่พูดถึงลักษณะเฉพาะที่นี่ จำกัด ตัวเราเพียงไม่กี่ภาพวาดของ สายพันธุ์ม้าทั่วไป (รูปที่ 113) .
การศึกษาโดยละเอียดโดย A. Strelkov พบว่า ciliates ประเภทต่างๆ นั้นยังห่างไกลจากการแพร่กระจายอย่างสม่ำเสมอตามลำไส้ใหญ่ของม้า มีสองกลุ่มของสายพันธุ์ที่แตกต่างกันสองสัตว์ตามที่เป็นอยู่ หนึ่งในนั้นอาศัยอยู่ในช่องท้องและส่วนท้องของลำไส้ใหญ่ (ส่วนเริ่มต้นของลำไส้ใหญ่) และอีกส่วนหนึ่งอาศัยอยู่ในส่วนหลังของลำไส้ใหญ่ขนาดใหญ่และลำไส้ใหญ่ขนาดเล็ก คอมเพล็กซ์ทั้งสองของสปีชีส์นี้มีการแบ่งเขตค่อนข้างชัดเจน มีไม่กี่ชนิดที่พบได้ทั่วไปในสองส่วนนี้ - น้อยกว่าหนึ่งโหล
, 
เป็นที่น่าสนใจที่จะสังเกตว่าในบรรดา ciliates หลายสายพันธุ์ที่อาศัยอยู่ในลำไส้ใหญ่ของ equids มีตัวแทนของสกุลเดียวกันที่เกี่ยวข้องกับการดูด ciliates ดังที่เราเห็นข้างต้น ดูด ciliates(Suctoria) เป็นสิ่งมีชีวิตที่อาศัยอยู่อย่างอิสระโดยทั่วไปโดยมีวิธีการเลี้ยงแบบพิเศษโดยใช้หนวด (รูปที่ 103) หนึ่งใน ที่ดูดนมปรับให้เข้ากับที่อยู่อาศัยที่ดูไม่ปกติ เช่น ลำไส้ใหญ่ของม้า เป็นต้น หลายชนิด อัลลันโทซิส(อัลลันโทโซมา). สัตว์ที่แปลกประหลาดเหล่านี้ (รูปที่ 114) ไม่มีก้านไม่มีตามีหนวดรูปไม้กระบองที่ปลายหนาขึ้น
ด้วยความช่วยเหลือของหนวด allantosomes ยึดติดกับ ciliates ประเภทต่างๆและดูดออก บ่อยครั้งเหยื่อมากกว่าผู้ล่าหลายเท่า
คำถามเกี่ยวกับธรรมชาติของความสัมพันธ์ระหว่าง ciliates ของลำไส้ใหญ่กับเจ้าภาพยังไม่ชัดเจน จำนวน ciliates อาจสูงและบางครั้งก็มากกว่าในกระเพาะของสัตว์เคี้ยวเอื้อง มีข้อมูลที่แสดงว่าจำนวน ciliates ในลำไส้ใหญ่ของม้าสามารถสูงถึง 3 ล้านใน 1 ซม. 3 ความสำคัญทางชีวภาพที่นักวิทยาศาสตร์บางคนแนะนำนั้นมีโอกาสน้อยกว่าสำหรับ ciliates ในกระเพาะรูเมน
ความคิดเห็นที่เป็นไปได้มากที่สุดคือพวกมันก่อให้เกิดอันตรายต่อโฮสต์โดยดูดซับอาหารจำนวนมาก ส่วนหนึ่งของ ciliates ถูกนำออกมาด้วยอุจจาระ ดังนั้นสารอินทรีย์ (รวมถึงโปรตีน) ที่ประกอบเป็นร่างกายของพวกมันยังคงไม่ได้ใช้โดยโฮสต์
คำถามเกี่ยวกับวิธีการติดเชื้อของม้าที่มี ciliates ที่อาศัยอยู่ในลำไส้ใหญ่ยังไม่ได้รับการแก้ไข

Balantidia จับอนุภาคอาหารที่หลากหลายจากเนื้อหาของลำไส้ใหญ่ เขากินเมล็ดแป้งด้วยความเต็มใจโดยเฉพาะอย่างยิ่ง หาก balantidia อาศัยอยู่ในรูของลำไส้ใหญ่ของมนุษย์ มันก็กินเนื้อหาของลำไส้และไม่มีผลร้ายใดๆ นี่เป็น "การขนส่ง" ทั่วไปซึ่งเราพบแล้วเมื่อพิจารณาถึงอะมีบาบิด อย่างไรก็ตาม balantidia มีโอกาสน้อยกว่าอะมีบาบิดเบี้ยวที่จะยังคงเป็น "ผู้เช่าที่ไม่เป็นอันตราย"
ปัจจุบันผู้เชี่ยวชาญได้พัฒนาอย่างดี วิธีการต่างๆทำให้สามารถเพาะเลี้ยง balantidia ในสภาพแวดล้อมเทียม - ภายนอกสิ่งมีชีวิตที่เป็นโฮสต์

ดังที่เห็นได้จากรูป troglodptella เป็นหนึ่งในเอนโดดินิโอมอร์ฟที่ซับซ้อน เธอนอกเหนือไปจากโซน perioral ของโรคตับแข็ง (ที่ส่วนหน้าของร่างกาย) มีอีกสามโซนของตับแข็งที่พัฒนามาอย่างดีซึ่งครอบคลุมร่างกายของ ciliate เป็นรูปวงแหวน Trogloditells มีเครื่องมือโครงกระดูกที่ได้รับการพัฒนามาอย่างดีซึ่งประกอบด้วยคาร์โบไฮเดรตซึ่งครอบคลุมส่วนหน้าเกือบทั้งหมดของร่างกาย ขนาดของ ciliates ที่แปลกประหลาดเหล่านี้ค่อนข้างสำคัญ มีความยาวถึง 200-280 ไมครอน
Ciliates ไร้ปาก ASTOMATS

การสนับสนุนการก่อตัวของโครงกระดูกส่วนใหญ่เกิดขึ้นที่ส่วนหน้าของร่างกาย ซึ่งจะต้องประสบกับความเครียดทางกลและเอาชนะอุปสรรค ผลักดันผ่านลูเมนของลำไส้ระหว่างอนุภาคอาหาร สายพันธุ์ สกุล radiophria(รังสีอัลตราไวโอเลต) ที่ส่วนหน้าด้านใดด้านหนึ่งของร่างกาย (ซึ่งตามอัตภาพถือว่าเป็นด้านท้อง) มีซี่โครงยืดหยุ่นที่แข็งแรงมาก (spicules) นอนอยู่ในชั้นผิวของ ectoplasm (รูปที่ 117, B, D, E) . สายพันธุ์ สกุล Menilella(Mesnilella) ยังมีรังสีค้ำจุน (spicules) ซึ่งส่วนใหญ่มีความยาวอยู่ในชั้นลึกของไซโตพลาสซึม (ในเอนโดพลาสซึม, รูปที่ 117, A). โครงสร้างรองรับที่จัดเรียงในทำนองเดียวกันนั้นได้รับการพัฒนาในสายพันธุ์ของสกุล Astomat อื่น ๆ
การสืบพันธุ์แบบไม่อาศัยเพศในบาง ciliates astomat ดำเนินการในลักษณะที่แปลกประหลาด แทนที่จะเป็นการแบ่งตามขวางเป็นสองส่วน ซึ่งเป็นลักษณะของ ciliates ส่วนใหญ่ แอสโทแมตจำนวนมากมีการแตกตัวไม่เท่ากัน (แตกหน่อ) ในเวลาเดียวกัน ไตที่แยกออกจากส่วนหลังยังคงสัมพันธ์กับมารดาเป็นระยะเวลาหนึ่ง (รูปที่ 117, B) เป็นผลให้ได้รับโซ่ซึ่งประกอบด้วยบุคคลที่เล็กกว่าด้านหน้าและด้านหลัง (ไต) ในอนาคตไตจะค่อยๆ แยกออกจากโซ่และเคลื่อนไปสู่การดำรงอยู่อย่างอิสระ รูปแบบการสืบพันธุ์ที่แปลกประหลาดนี้แพร่หลายเช่นใน radiophria ที่เรารู้จักแล้ว ห่วงโซ่ของแอสโทมาตาบางส่วนที่เกิดจากการแตกหน่อคล้ายกับ รูปร่างห่วงโซ่ของพยาธิตัวตืด ที่นี่อีกครั้งเราพบปรากฏการณ์ของการบรรจบกัน
เครื่องมือนิวเคลียร์ของแอสโทแมทมีลักษณะโครงสร้างของซิลิเอต: มาโครนิวเคลียส ซึ่งส่วนใหญ่มักจะมีรูปร่างคล้ายริบบิ้น (รูปที่ 117) และหนึ่งไมโครนิวเคลียส แวคิวโอลที่หดตัวมักจะได้รับการพัฒนาอย่างดี สปีชีส์ส่วนใหญ่มีแวคิวโอลหดตัวหลายอัน (บางครั้งมีมากกว่าหนึ่งโหล) เรียงกันเป็นแถวตามยาวหนึ่งแถว
การศึกษาการกระจายของสายพันธุ์ Astomat ตามโฮสต์ประเภทต่างๆ แสดงให้เห็นว่าสายพันธุ์ Astomat ส่วนใหญ่ถูกกักขังอยู่ในสายพันธุ์โฮสต์ที่กำหนดไว้อย่างเคร่งครัด astomats ส่วนใหญ่มีลักษณะเฉพาะที่แคบ: มีสัตว์เพียงชนิดเดียวเท่านั้นที่สามารถทำหน้าที่เป็นโฮสต์สำหรับพวกมัน
แม้จะมีการศึกษาจำนวนมากที่อุทิศให้กับการศึกษาแอสโทแมทซิลิเอต แต่แง่มุมที่สำคัญมากอย่างหนึ่งของชีววิทยาของพวกมันก็ยังไม่ชัดเจนนัก: การถ่ายทอดของ ciliates เหล่านี้จากโฮสต์หนึ่งไปยังอีกบุคคลหนึ่งเกิดขึ้นได้อย่างไร ไม่เคยสังเกตการก่อตัวของซีสต์ใน ciliates เหล่านี้มาก่อน
ดังนั้นจึงแนะนำว่าการติดเชื้อเกิดขึ้นอย่างแข็งขัน - ระยะเคลื่อนที่
อวัยวะภายในลำไส้ของเม่นทะเล
เม่นทะเลมีอยู่มากมายในเขตชายฝั่งทะเลทางตอนเหนือของเรา (เรนท์) และทะเลตะวันออกไกล (ทะเลญี่ปุ่น ชายฝั่งแปซิฟิกของหมู่เกาะคูริล) ข้างมาก เม่นทะเลกินอาหารจากพืช ซึ่งส่วนใหญ่เป็นสาหร่าย ซึ่งพวกมันขูดวัตถุใต้น้ำด้วย "ฟัน" ที่แหลมเป็นพิเศษรอบปากที่เปิดอยู่ ในลำไส้ของเม่นที่กินพืชเป็นอาหารมีสัตว์จำพวก ciliates มากมาย บ่อยครั้งที่พวกมันพัฒนาในปริมาณมาก และเนื้อหาของลำไส้ของเม่นทะเลภายใต้กล้องจุลทรรศน์นั้นเกือบจะ "เต็มไปด้วย" กับ ciliates เป็นเนื้อหาของกระเพาะรูเมนของสัตว์เคี้ยวเอื้อง ต้องบอกว่านอกจากความแตกต่างอย่างลึกซึ้งในสภาพความเป็นอยู่ของ ciliates ของลำไส้ของเม่นทะเลและกระเพาะของสัตว์เคี้ยวเอื้องแล้ว มีความคล้ายคลึงกันบางประการ พวกเขาอยู่ในความจริงที่ว่าทั้งที่นี่และที่นั่น ciliates อาศัยอยู่ในสภาพแวดล้อมที่อุดมไปด้วยซากพืช ปัจจุบัน ciliates กว่า 50 สายพันธุ์เป็นที่รู้กันว่าอาศัยอยู่ในลำไส้ของเม่นทะเล ซึ่งพบได้เฉพาะในเขตชายฝั่งทะเลที่ซึ่งหอยเม่นกินสาหร่าย ที่ระดับความลึกมาก ซึ่งสาหร่ายไม่เติบโตอีกต่อไป ไม่มี ciliates ในเม่นทะเล

ตามวิถีชีวิตและธรรมชาติของโภชนาการ ciliates ลำไส้ของเม่นทะเลส่วนใหญ่เป็นพืชกินพืช พวกมันกินสาหร่ายซึ่งในปริมาณมากจะเติมลำไส้ของเจ้าบ้าน บางชนิดค่อนข้าง "จู้จี้จุกจิก" ในการเลือกอาหาร ตัวอย่างเช่น, สโตรบิลิเดียม(สโตรบิลิเดียม, รูปที่ 118, A) กินไดอะตอมขนาดใหญ่เกือบทั้งหมด นอกจากนี้ยังมีนักล่าที่กินตัวแทนของผู้อื่นอีกด้วย สายพันธุ์เล็กซิลิเอต
ใน ciliates จากลำไส้ของเม่นทะเลซึ่งแตกต่างจาก astomat ไม่มีการกักขังที่เข้มงวดสำหรับโฮสต์บางประเภท พวกมันอาศัยอยู่ในเม่นทะเลที่กินสาหร่ายหลากหลายชนิด
ยังไม่มีการศึกษาวิธีการติดเชื้อของเม่นทะเลที่มี ciliates อย่างไรก็ตาม ที่นี่มีความเป็นไปได้สูงที่สามารถสันนิษฐานได้ว่ามันเกิดขึ้นในรูปแบบลอยอิสระที่ใช้งานอยู่ ความจริงก็คือ ciliates จากลำไส้ของเม่นทะเลสามารถมีชีวิตอยู่เป็นเวลานาน (หลายชั่วโมง) ในน้ำทะเล อย่างไรก็ตาม พวกมันได้ปรับตัวเข้ากับชีวิตในลำไส้ของเม่นอย่างมากจนตายนอกร่างกายในน้ำทะเลไม่ช้าก็เร็ว
เมื่อทำความคุ้นเคยกับ ciliates เสร็จแล้วควรเน้นอีกครั้งว่าพวกเขาเป็นตัวแทนของสายพันธุ์ที่อุดมสมบูรณ์ซึ่งเป็นกลุ่ม (คลาส) ที่กว้างขวางและเจริญรุ่งเรืองของสัตว์โลก ยังคงอยู่ในระดับของการจัดระเบียบเซลล์ infusoria ได้มาถึงเมื่อเทียบกับโปรโตซัวประเภทอื่น ๆ ความซับซ้อนที่ยิ่งใหญ่ที่สุดของโครงสร้างและหน้าที่
บทบาทสำคัญโดยเฉพาะอย่างยิ่งในการพัฒนาที่ก้าวหน้า (วิวัฒนาการ) นี้อาจเล่นโดยการเปลี่ยนแปลงของอุปกรณ์นิวเคลียร์และการเกิดขึ้นของความเป็นคู่ทางนิวเคลียร์ (เชิงคุณภาพไม่สมมูลของนิวเคลียส) ความสมบูรณ์ของมาโครนิวเคลียสในสารนิวคลีอิกนั้นสัมพันธ์กับกระบวนการเมแทบอลิซึมที่ทำงานอยู่ โดยมีกระบวนการสังเคราะห์โปรตีนที่เข้มข้นในไซโตพลาสซึมและนิวเคลียส
บทสรุป
เรามาถึงจุดสิ้นสุดของการทบทวนโครงสร้างและวิถีชีวิตของสัตว์โลกที่กว้างขวาง - โปรโตซัว. คุณลักษณะเฉพาะของพวกมันดังที่ได้กล่าวไว้ข้างต้นซ้ำแล้วซ้ำเล่าคือความเป็นเซลล์เดียว ในแง่ของโครงสร้างโปรโตซัวเป็นเซลล์ อย่างไรก็ตาม พวกมันเทียบไม่ได้กับเซลล์ที่ประกอบเป็นร่างกายของสิ่งมีชีวิตหลายเซลล์ เพราะพวกมันเองเป็นสิ่งมีชีวิต ดังนั้นโปรโตซัวจึงเป็นสิ่งมีชีวิตในระดับเซลล์ขององค์กร โปรโตซัวที่มีการจัดระเบียบสูงบางตัวซึ่งมีนิวเคลียสจำนวนมาก ดูเหมือนจะเกินขีดจำกัดทางสัณฐานวิทยาของโครงสร้างเซลล์แล้ว ซึ่งทำให้นักวิทยาศาสตร์บางคนมีเหตุผลที่จะเรียกโปรโตซัวดังกล่าวว่า "เหนือเซลล์" สิ่งนี้เปลี่ยนแปลงสาระสำคัญของเรื่องนี้เพียงเล็กน้อย เนื่องจากองค์กรที่มีเซลล์เดียวยังคงเป็นแบบอย่างของโปรโตซัว
ภายในขอบเขตของความเป็นเซลล์เดียว โปรโตซัวมีการพัฒนาทางวิวัฒนาการที่ยาวนานและได้ให้รูปแบบที่หลากหลายซึ่งปรับให้เข้ากับสภาพชีวิตที่หลากหลายที่สุด ที่ใจกลางของต้นกำเนิดสายเลือดของโปรโตซัวมีสองประเภท: sarcodes และ flagellates คำถามที่ว่าชั้นเรียนเหล่านี้มีพื้นฐานมากขึ้นยังคงถูกถกเถียงกันในทางวิทยาศาสตร์ ในอีกด้านหนึ่งตัวแทนที่ต่ำกว่าของ Sarcodidae (ameba) มีโครงสร้างดั้งเดิมที่สุด แต่แฟลเจลเลตแสดงให้เห็นถึงความเป็นพลาสติกที่ยิ่งใหญ่ที่สุดของประเภทของการเผาผลาญและยืนอยู่บนพรมแดนระหว่างโลกของสัตว์และพืช ในวงจรชีวิตของ sarcodes บางตัว (เช่น foraminifera) มีแฟลเจลลาร์สเตจ (gametes) ซึ่งบ่งชี้ความสัมพันธ์ของพวกมันกับแฟลเจลเลต เป็นที่ชัดเจนว่าทั้ง sarcods สมัยใหม่และ flagellates สมัยใหม่ไม่สามารถเป็นกลุ่มเริ่มต้นของวิวัฒนาการของโลกของสัตว์ได้เนื่องจากพวกเขาได้พัฒนาประวัติศาสตร์ไปไกลแล้วและได้พัฒนาการปรับตัวให้เข้ากับสภาพความเป็นอยู่สมัยใหม่บนโลก อาจเป็นไปได้ว่าโปรโตซัวสมัยใหม่ทั้งสองคลาสนี้ควรได้รับการพิจารณาว่าเป็นสองลำต้นในวิวัฒนาการซึ่งมีต้นกำเนิดมาจากรูปแบบโบราณที่ยังไม่รอดมาจนถึงทุกวันนี้ซึ่งอาศัยอยู่ในช่วงรุ่งอรุณของการพัฒนาชีวิตบนโลกของเรา
ในการวิวัฒนาการต่อไปของโปรโตซัว การเปลี่ยนแปลงของธรรมชาติที่แตกต่างกันเกิดขึ้น บางส่วนนำไปสู่การเพิ่มระดับขององค์กรโดยทั่วไป กิจกรรมที่เพิ่มขึ้น และความเข้มข้นของกระบวนการชีวิต ในบรรดาการเปลี่ยนแปลงทางสายวิวัฒนาการ (วิวัฒนาการ) นั้น เราควรรวมถึงการพัฒนาออร์แกเนลล์ของการเคลื่อนไหวและการดักจับอาหาร ซึ่งได้บรรลุถึงความสมบูรณ์แบบในระดับสูงในกลุ่ม ciliates ปฏิเสธไม่ได้ว่า cilia เป็นออร์แกเนลล์ที่สัมพันธ์กัน (คล้ายคลึงกัน) กับแฟลกเจลลา ในขณะที่อยู่ใน flagellates มีข้อยกเว้นบางประการจำนวน flagella มีขนาดเล็กใน ciliates จำนวน cilia ถึงหลายพัน การพัฒนาเครื่องมือปรับเลนส์ปรับเลนส์เพิ่มกิจกรรมของโปรโตซัวอย่างรวดเร็วทำให้รูปแบบของความสัมพันธ์กับสิ่งแวดล้อมมีความหลากหลายและซับซ้อนมากขึ้นรูปแบบของปฏิกิริยาต่อสิ่งเร้าภายนอก การปรากฏตัวของเครื่องมือปรับเลนส์ที่แตกต่างกันอย่างไม่ต้องสงสัยเป็นหนึ่งในสาเหตุหลักของวิวัฒนาการที่ก้าวหน้าในกลุ่ม ciliates ที่มีรูปแบบที่หลากหลายซึ่งปรับให้เข้ากับแหล่งที่อยู่อาศัยที่แตกต่างกัน
การพัฒนาเครื่องมือปรับเลนส์ของ ciliates เป็นตัวอย่างของการเปลี่ยนแปลงเชิงวิวัฒนาการประเภทนี้ซึ่ง Acad ตั้งชื่อให้ Severtsov อะโรมอร์โฟส Aromorphoses มีลักษณะเฉพาะในองค์กรที่เพิ่มขึ้นโดยทั่วไปการพัฒนาการปรับตัวที่มีนัยสำคัญในวงกว้าง การเพิ่มขึ้นขององค์กรเป็นที่เข้าใจกันว่าการเปลี่ยนแปลงที่ทำให้กิจกรรมที่สำคัญของสิ่งมีชีวิตเพิ่มขึ้น พวกมันสัมพันธ์กับความแตกต่างในหน้าที่ของส่วนต่าง ๆ ซึ่งนำไปสู่รูปแบบการสื่อสารที่หลากหลายมากขึ้นระหว่างสิ่งมีชีวิตกับสิ่งแวดล้อม การพัฒนาเครื่องมือปรับเลนส์ของ ciliates หมายถึงการเปลี่ยนแปลงโครงสร้างในกระบวนการวิวัฒนาการอย่างแม่นยำ นี่เป็นลักษณะอะโรมอร์โฟซิสทั่วไป
ในโปรโตซัวตามที่ V. A. Dogel เน้น การเปลี่ยนแปลงประเภทของอะโรมอร์โฟสมักจะเกี่ยวข้องกับการเพิ่มจำนวนของออร์แกเนลล์ การเกิดพอลิเมอไรเซชันของออร์แกเนลล์เกิดขึ้น การพัฒนาเครื่องมือปรับเลนส์ใน ciliates เป็นตัวอย่างทั่วไปของการเปลี่ยนแปลงดังกล่าว ตัวอย่างที่สองของอะโรมอร์โฟซิสในวิวัฒนาการของ ciliates คือเครื่องมือนิวเคลียร์ของพวกมัน เราได้ตรวจสอบลักษณะโครงสร้างของแกน ciliates ข้างต้นแล้ว การเกิดคู่ของนิวเคลียสของ ciliates (การปรากฏตัวของไมโครนิวเคลียสและมาโครนิวเคลียส) มาพร้อมกับการเพิ่มจำนวนของโครโมโซมในมาโครนิวเคลียส (ปรากฏการณ์ของ polyploidy) เนื่องจากโครโมโซมสัมพันธ์กับกระบวนการสังเคราะห์หลักในเซลล์ โดยหลักแล้วกับการสังเคราะห์โปรตีน กระบวนการนี้จึงทำให้ความเข้มของหน้าที่หลักที่สำคัญเพิ่มขึ้นโดยทั่วไป ที่นี่เช่นกันการเกิดพอลิเมอไรเซชันซึ่งส่งผลต่อโครโมโซมเชิงซ้อนของนิวเคลียส
ciliates- หนึ่งในกลุ่มโปรโตซัวที่มีจำนวนมากที่สุดและก้าวหน้าที่สุด มาจากแฟลเจลเลต นี่คือหลักฐานโดยความคล้ายคลึงกันทางสัณฐานวิทยาที่สมบูรณ์ของออร์กานอยด์ของการเคลื่อนไหว ขั้นตอนของการวิวัฒนาการนี้เกี่ยวข้องกับอะโรมอร์โฟสขนาดใหญ่สองชนิด: หนึ่งในนั้นส่งผลกระทบต่อออร์แกเนลล์ของการเคลื่อนไหว ประการที่สอง - เครื่องมือนิวเคลียร์ การเปลี่ยนแปลงทั้งสองประเภทนี้เชื่อมโยงถึงกัน เนื่องจากทั้งสองนำไปสู่การเพิ่มกิจกรรมที่สำคัญและความซับซ้อนของรูปแบบของความสัมพันธ์กับสภาพแวดล้อมภายนอก
นอกจากอะโรมอร์โฟสแล้ว ยังมีการเปลี่ยนแปลงเชิงวิวัฒนาการอีกประเภทหนึ่ง ซึ่งแสดงออกมาในการพัฒนาการปรับตัว (การปรับตัว) ให้เข้ากับเงื่อนไขการดำรงอยู่ที่กำหนดไว้อย่างชัดเจน การเปลี่ยนแปลงเชิงวิวัฒนาการประเภทนี้เรียกว่า idioadaptation โดย Severtsov ในวิวัฒนาการของโปรโตซัว การเปลี่ยนแปลงประเภทนี้มีบทบาทสำคัญมาก ข้างต้นเมื่อพิจารณา คลาสต่างๆโปรโตซัว มีตัวอย่างมากมายของการเปลี่ยนแปลงที่ไม่ทราบสาเหตุ การปรับตัวให้เข้ากับวิถีชีวิตของแพลงก์ตอนในกลุ่มโปรโตซัวต่าง ๆ การปรับตัวให้เข้ากับชีวิตในทรายใน ciliates การก่อตัวของเปลือกป้องกันของโอโอซิสต์ใน coccidia และอื่น ๆ อีกมากมาย - ทั้งหมดนี้เป็นการปรับตัวที่มีบทบาทสำคัญในการเกิดขึ้นและการพัฒนา ของแต่ละกลุ่ม แต่ไม่เกี่ยวข้องกับการเปลี่ยนแปลงทั่วไปในองค์กร
การปรับตัวให้เข้ากับแหล่งที่อยู่อาศัยเฉพาะต่างๆ ในโปรโตซัวนั้นมีความหลากหลายมาก พวกเขารับประกันการกระจายของประเภทนี้ในแหล่งที่อยู่อาศัยที่หลากหลาย ซึ่งได้กล่าวถึงในรายละเอียดข้างต้น เมื่ออธิบายแต่ละชั้นเรียน
สารานุกรมทางการแพทย์ขนาดใหญ่
Infusoria Tetrahymena thermophila ... Wikipedia
- (Infusoria) คลาสของโปรโตซัวที่พัฒนาอย่างสูงที่สุด (Protozoa) คุณสมบัติหลักของ I.: การปรากฏตัวของ cilia (สำหรับการเคลื่อนไหวและโภชนาการ), นิวเคลียสสองประเภท (polyploid macronucleus และ diploid micronucleus, โครงสร้างที่แตกต่างกันและ ... ... สารานุกรมแห่งสหภาพโซเวียตผู้ยิ่งใหญ่
หรือสัตว์ของเหลว (Infusoria) กลุ่มของสัตว์โปรโตซัวที่มีผมเปียหรือตาเป็นประกาย บางครั้งถูกแทนที่ในวัยผู้ใหญ่ด้วยผลพลอยได้จากการดูดทรงกระบอกที่มีรูปร่างที่แน่นอน ส่วนใหญ่มี ... ... พจนานุกรมสารานุกรมเอฟ Brockhaus และ I.A. เอฟรอน
อนุกรมวิธานของการจำแนก ifusoria และการจัดระบบของสิ่งมีชีวิตที่เป็นของประเภทของ Ciliates ตามแนวคิดทั่วไปของอนุกรมวิธานของโปรโตซัวโดยเฉพาะและสัตว์ในเนื้อหาทั่วไป 1 ระบบ 1.1 คลาสสิก ... Wikipedia
หรือสัตว์ของเหลว (Infusoria) กลุ่มของสัตว์โปรโตซัวพร้อมกับถักเปียหรือตาที่ส่องแสงซึ่งในสภาพผู้ใหญ่บางครั้งจะถูกแทนที่ด้วยผลพลอยได้จากการดูดทรงกระบอกที่มีรูปร่างที่แน่นอนโดยส่วนใหญ่มี ... ... สารานุกรมของ Brockhaus และ Efron
ประเภทของ Ciliates เรียกอีกอย่างว่า Ciliated - อวัยวะของการเคลื่อนไหวของโปรโตซัวเหล่านี้ cilia. เซลล์ซิลิเอตมี 2 นิวเคลียส เรียกว่าเซลล์ขนาดเล็กและขนาดใหญ่ ประการแรกควบคุมกระบวนการสืบพันธุ์และส่วนที่สองรับผิดชอบกระบวนการด้านโภชนาการการเคลื่อนไหวและการหายใจ
คุณลักษณะของชีวิตประเภทนี้ควรพิจารณาในตัวอย่างของรองเท้า ciliates
การเคลื่อนไหวและการหายใจ
รองเท้า infusoria ซึ่งมีความยาวประมาณ 0.5 มม. เลือกแหล่งน้ำเป็นที่อยู่อาศัย รูปร่างของร่างกายที่ง่ายที่สุดนั้นง่ายต่อการเดาโดยใช้ชื่อ - คล้ายกับรองเท้า ความเร็วในการเคลื่อนที่ประมาณ 2.5 มม. ต่อวินาที
การปรากฏตัวของเปลือกยืดหยุ่นด้านนอกทำให้รูปร่างมั่นคง
ในไซโตพลาสซึมซึ่งอยู่ติดกับเปลือกมีเส้นใยรองรับการพัฒนาของพวกมันคือการรับประกันการรักษารูปแบบคงที่ของ ciliate
บนพื้นผิวของ ciliates มี 15,000 cilia ที่ฐานมีร่างกายเป็นฐาน การเคลื่อนไหวเกิดขึ้นด้วยความช่วยเหลือของการสั่นสะเทือนของ cilia: พวกเขาสร้างประมาณ 30 จังหวะต่อวินาทีจึงผลัก ciliate-shoe ไปข้างหน้า
มันหายใจผ่านพื้นผิวของร่างกาย
โภชนาการ
คุณสมบัติของ ciliates คือการมีอยู่ ปากเซลล์ใกล้ๆ ซึ่งมีตาที่ยาวและหนาแน่นเป็นพิเศษ ปากเซลล์ดำเนินต่อไปด้วยคอหอยเซลล์: ตาดันน้ำและอาหารซิลิเอต - แบคทีเรียเข้าไป
Infusoria สัมผัสสารเคมีซึ่งปล่อยกลุ่มแบคทีเรีย ดังนั้นเธอจึงแสวงหาเหยื่อ
อาหารจะจบลงในแวคิวโอลย่อยอาหารซึ่งจะถูกย่อย จากที่นี่ไปตามไซโตพลาสซึมแล้ว
การคัดเลือก
การคัดเลือกจะดำเนินการโดยใช้ สองแวคิวโอลหดตัวอันหนึ่งอยู่ที่ส่วนหน้า อีกอันหนึ่งอยู่ที่ด้านหลัง Vacuoles ประกอบด้วยอ่างเก็บน้ำและช่อง
ของเหลวจะเติมลงในช่องต่างๆ จากนั้นจึงไหลไปตามอ่างเก็บน้ำตรงกลาง จากนั้นจึงออกจากช่อง infusoria กระบวนการหดตัวของแวคิวโอลใช้เวลา 10-20 วินาที
การสืบพันธุ์
Infusoria สืบพันธุ์แบบไม่อาศัยเพศ - แบ่งออกเป็นสองส่วน ลักษณะของมันคือการแบ่งตามร่างกาย
นิวเคลียสของ ciliates แบ่งออกเป็นสองส่วน: ciliates ที่เพิ่งสร้างใหม่มีนิวเคลียสขนาดเล็กและใหญ่ ciliates ลูกสาวมีส่วนของออร์กานอยด์และส่วนที่หายไปนั้นถูกสร้างขึ้นอย่างอิสระ การสืบพันธุ์เกิดขึ้นหลายครั้งต่อวัน
สำหรับรองเท้า ciliates การสืบพันธุ์แบบอาศัยเพศก็เป็นไปได้เช่นกัน แต่ในกรณีนี้ไม่มีการเพิ่มจำนวนบุคคล ชั่วคราว โปรโตซัวเชื่อมต่อกัน ก่อตัวเป็นสะพานเชื่อมจากไซโตพลาสซึม
ในแต่ละบุคคล นิวเคลียสขนาดใหญ่จะหายไป และนิวเคลียสขนาดเล็กจะถูกแบ่งสองครั้ง - สี่นิวเคลียสปรากฏขึ้น ในจำนวนนี้เหลือเพียงนิวเคลียสเดียวเท่านั้นซึ่งแบ่งออกด้วย บุคคลประกอบด้วยนิวเคลียสสองนิวเคลียส จากนั้นเกิดการแลกเปลี่ยนนิวเคลียส - นิวเคลียสตัวหนึ่งเคลื่อนที่ไปยังอีกบุคคลหนึ่ง
ที่นั่นจะรวมเข้ากับนิวเคลียสที่หลงเหลืออยู่ ดังนั้นจึงมีนิวเคลียสขนาดเล็กและใหญ่ขึ้นในแต่ละบุคคล กระบวนการนี้เรียกว่า ผันเป็นสิ่งจำเป็นสำหรับการต่ออายุสารพันธุกรรมระหว่างบุคคล
ประเภทของ ciliates
Ciliates จัดเป็นโปรโตซัวที่ซับซ้อน มีประมาณ 7,000 สายพันธุ์
1. sarcodes ที่หลากหลาย
2. แฟลกเจลลาต่างๆ
ตัวแทนของประเภทย่อยที่สองย้ายด้วยความช่วยเหลือของแฟลกเจลลาซึ่งสะท้อนให้เห็นในชื่อของกลุ่มนี้
- แฟลกเจลลา ยูกลีนากรีนเป็นตัวแทนทั่วไปของกลุ่มนี้ โปรโตซัวนี้มีรูปร่างเป็นฟิวซิฟอร์มปกคลุมด้วยเมมเบรนยืดหยุ่นหนาแน่น ที่ส่วนหน้าของร่างกายมีแฟลเจลลัมใกล้กับฐานซึ่งมีตาที่ไวต่อแสง - มลทินและแวคิวโอลที่หดตัว ใกล้กับปลายด้านหลังในความหนาของไซโตพลาสซึมมีนิวเคลียสที่ควบคุมกระบวนการสำคัญทั้งหมดของร่างกาย สีเขียวของยูกลีนาเกิดจากคลอโรพลาสต์จำนวนมาก แสงแฟลเจลเลตนี้สังเคราะห์แสง แต่เมื่ออยู่ในพื้นที่ที่ไม่มีแสงสว่างของอ่างเก็บน้ำเป็นเวลานาน มันจะเปลี่ยนไปกินผลิตภัณฑ์ที่เน่าเปื่อยของสารอินทรีย์ที่ซับซ้อนโดยแยกพวกมันออกจากสิ่งแวดล้อม ดังนั้น ยูกลีนาสีเขียวจึงสามารถรวมทั้งการเผาผลาญ autotrophic และ heterotrophic ได้ การดำรงอยู่ของสิ่งมีชีวิตที่มีสารอาหารแบบผสมดังกล่าวบ่งบอกถึงความสัมพันธ์ระหว่างโลกของสัตว์และพืช แฟลกเจลลาส่วนใหญ่
3. สปอโรซัวและ ciliates ที่หลากหลาย
ประเภท Ciliates รวมถึงโปรโตซัวซึ่งมีความโดดเด่นด้วยองค์กรที่ซับซ้อนที่สุดในบรรดาสัตว์ที่มีเซลล์เดียว ตัวแทนทั่วไปของกลุ่มนี้คือ infusoria-shoe ซึ่งเป็นสัตว์น้ำจืดทั่วไป ร่างกายของเธอถูกปกคลุมไปด้วยเปลือกหนาแน่น - เกล็ดจึงมีรูปร่างค่อนข้างคงที่ ลักษณะเฉพาะของโครงสร้างคือการมี cilia ซึ่งปกคลุมทั่วทั้งตัวรองเท้าอย่างสม่ำเสมอ การเคลื่อนไหวของ cilia นั้นประสานกันเนื่องจากเครือข่ายของเส้นใยหดตัวที่อยู่ในชั้นผิวของไซโตพลาสซึม ลักษณะเด่นประการที่สองคือการมีอยู่ของนิวเคลียสสองนิวเคลียส ขนาดใหญ่ (มาโครนิวเคลียส) และขนาดเล็ก (ไมโครนิวเคลียส) นิวเคลียสยังแตกต่างกันไปตามหน้าที่: นิวเคลียสขนาดใหญ่ควบคุมการเผาผลาญและนิวเคลียสขนาดเล็กมีส่วนร่วมในกระบวนการทางเพศ (ผัน) รองเท้า infusoria กินแบคทีเรีย สาหร่ายเซลล์เดียว ซึ่งปรับโดย cilia ไปที่ปากเซลล์ ซึ่งอยู่ที่ด้านล่างของช่องปากก่อนช่องปาก หลังจากผ่านคอหอยของเซลล์ซึ่งสิ้นสุดในไซโตพลาสซึม อนุภาคอาหารจะถูกปิดล้อมในแวคิวโอลย่อยอาหาร ซึ่งพวกมันถูกทำลายลงโดยการกระทำของเอนไซม์ สารตกค้างที่ไม่ได้แยกแยะจะถูกโยนออกทางผง ในร่างกายของรองเท้ามีแวคิวโอลการหดตัวเชิงซ้อนที่ซับซ้อนเป็นจังหวะสลับกันสองอัน รองเท้าสืบพันธุ์แบบไม่อาศัยเพศโดยแบ่งครึ่ง การสืบพันธุ์แบบไม่อาศัยเพศหลายครั้งถูกแทนที่ในรองเท้าด้วยกระบวนการทางเพศ - การผันคำกริยา ในระหว่างที่รองเท้าสองคู่มารวมกันและแลกเปลี่ยนสารพันธุกรรม หลังจากนั้นรองเท้าก็จะแยกออกและเริ่มการสืบพันธุ์แบบไม่อาศัยเพศอีกครั้งในไม่ช้า ความสำคัญทางชีวภาพของการผันคำกริยาอยู่ในการรวมกันในสิ่งมีชีวิตเดียวของคุณสมบัติทางพันธุกรรมของบุคคลสองคน สิ่งนี้เพิ่มความมีชีวิตซึ่งแสดงออกถึงความสามารถในการปรับตัวให้เข้ากับสภาพแวดล้อมได้ดีขึ้น
ไม่พบสิ่งที่คุณกำลังมองหา? ใช้การค้นหา
ในหน้านี้ เนื้อหาในหัวข้อ:
- สั้น ๆ เกี่ยวกับ flagellates, pseudopods, sporozoans และ ciliates
- sarcodes และ sporozoans
- แฟลกเจลลาหลากหลายชนิด
- สรุปรายงานรองเท้าแตะ infusoria
- อะไรคือความแตกต่างระหว่าง ciliates และ sarcode species
หนึ่งในตัวแทนที่รู้จักกันดีที่สุดของเลนส์ปรับเลนส์คือรองเท้า ciliate ตามกฎแล้วอาศัยอยู่ในน้ำที่มีทิศทางนิ่งเช่นเดียวกับในอ่างเก็บน้ำประเภทน้ำจืดซึ่งกระแสมีความโดดเด่นยกเว้นความแน่วแน่ ที่อยู่อาศัยของมันจะต้องมีอินทรียวัตถุที่เน่าเปื่อย ขอแนะนำให้พิจารณารายละเอียดทุกด้านของชีวิตของตัวแทนสัตว์เหล่านี้อย่างละเอียด
ตัวแทนขนตา

ควรสังเกตว่า Ciliates เป็นประเภทที่มีชื่อมาจากคำว่า "tincture" (แปลจาก ละติน). สิ่งนี้สามารถอธิบายได้ด้วยความจริงที่ว่าตัวแทนแรกของโปรโตซัวถูกพบอย่างแม่นยำในทิงเจอร์สมุนไพร เมื่อเวลาผ่านไป การพัฒนาประเภทนี้เริ่มได้รับแรงผลักดันอย่างรวดเร็ว ดังนั้นในปัจจุบันทางชีววิทยาจึงรู้จักประมาณ 6-7,000 สปีชีส์ซึ่งรวมถึงประเภทของ Ciliates หากเราอาศัยข้อมูลของทศวรรษ 1980 ก็อาจกล่าวได้ว่าประเภทที่เป็นปัญหาประกอบด้วยสองคลาสในโครงสร้าง: Ciliated ciliates (มีสาม superorders) และ Sucking ciliates ในการเชื่อมต่อกับข้อมูลนี้ เราสามารถสรุปได้ว่าความหลากหลายของสิ่งมีชีวิตนั้นกว้างมาก ซึ่งเป็นเรื่องที่น่าสนใจอย่างแท้จริง
ประเภทของ Infusoria: ตัวแทน

ตัวแทนที่โดดเด่นของประเภทนี้คือ ciliates-balantidia และ ciliates-shoe คุณสมบัติที่โดดเด่นสัตว์เหล่านี้เป็นเปลือกของเปลือกที่มีตาซึ่งใช้สำหรับการเคลื่อนไหวการป้องกัน ciliates ผ่านอวัยวะที่ออกแบบมาเป็นพิเศษ Trichocysts (ตั้งอยู่ใน ectoplasm ของเปลือก) รวมถึงการปรากฏตัวในเซลล์ของสองนิวเคลียส (พืช) และกำเนิด) นอกจากนี้ ช่องปากในร่างกายของ ciliate ยังก่อให้เกิดช่องทางในช่องปาก ซึ่งมีแนวโน้มที่จะผ่านเข้าไปในปากของเซลล์ที่นำไปสู่คอหอย มีการสร้างแวคิวโอลย่อยอาหารซึ่งทำหน้าที่ย่อยอาหารโดยตรง แต่ส่วนประกอบที่ไม่ได้แยกแยะจะถูกลบออกจากร่างกายผ่านทางแป้ง ลักษณะของชนิดของ ciliatesหลากหลายมาก แต่ประเด็นหลักจะกล่าวถึงข้างต้น สิ่งเดียวที่ต้องเพิ่มคือ ciliates ทั้งสองอยู่ในส่วนตรงข้ามของร่างกาย โดยการทำงานของพวกเขาที่น้ำส่วนเกินหรือผลิตภัณฑ์เมตาบอลิซึมถูกขับออกจากร่างกาย
รองเท้า Infusoria

เพื่อพิจารณาคุณภาพโครงสร้างและวิถีชีวิตของสิ่งมีชีวิตที่น่าสนใจดังกล่าวของโครงสร้างเซลล์เดียว ขอแนะนำให้อ้างอิงถึงตัวอย่างที่เกี่ยวข้อง สิ่งนี้ต้องใช้รองเท้า infusoria ซึ่งแพร่หลายในแหล่งน้ำจืด พวกเขาสามารถผสมพันธุ์ในภาชนะธรรมดาได้อย่างง่ายดาย (เช่นในตู้ปลา) อ่าวหญ้าแห้งทุ่งหญ้านั้นง่ายที่สุด น้ำจืดเพราะในทิงเจอร์ประเภทนี้ตามกฎแล้วโปรโตซัวหลายชนิดพัฒนารวมถึงรองเท้า ciliates ดังนั้นคุณจึงสามารถศึกษาข้อมูลทั้งหมดที่มีอยู่ในบทความได้โดยใช้กล้องจุลทรรศน์
ลักษณะของรองเท้า ciliates
ดังที่กล่าวไว้ข้างต้น Ciliates เป็นประเภทที่มีองค์ประกอบหลายอย่าง สิ่งที่น่าสนใจที่สุดคือรองเท้า ciliate นี่คือความยาวครึ่งมิลลิเมตรซึ่งมีรูปร่างเป็นแกนหมุน ควรสังเกตว่าการมองเห็นสิ่งมีชีวิตนี้คล้ายกับรองเท้าดังนั้นจึงเป็นชื่อที่น่าสนใจ รองเท้า infusoria มีการเคลื่อนไหวตลอดเวลา และว่ายโดยปลายทู่ไปข้างหน้า ที่น่าสนใจคือความเร็วของการเคลื่อนที่มักจะสูงถึง 2.5 มม. ต่อวินาที ซึ่งดีมากสำหรับตัวแทนประเภทนี้ บนพื้นผิวของร่างกายของ ciliates-shoes สามารถสังเกตได้ว่า cilia ทำหน้าที่เป็นอวัยวะของมอเตอร์ เช่นเดียวกับ ciliates ทั้งหมด สิ่งมีชีวิตที่เป็นปัญหามีนิวเคลียสสองนิวเคลียสในโครงสร้างของมัน: นิวเคลียสขนาดใหญ่มีหน้าที่รับผิดชอบในกระบวนการทางโภชนาการ ทางเดินหายใจ ยนต์ และเมตาบอลิซึม และนิวเคลียสที่มีขนาดเล็กมีส่วนร่วมในด้านเพศ
ร่างกายของรองเท้า ciliates

โครงสร้างลำตัวของรองเท้า ciliates นั้นซับซ้อนมาก การเคลือบด้านนอกของตัวแทนนี้เป็นเปลือกยางยืดแบบบาง สามารถรักษารูปร่างของร่างกายให้ถูกต้องตลอดชีวิต ผู้ช่วยที่ซื่อสัตย์ในเรื่องนี้ได้รับการพัฒนาอย่างสมบูรณ์เส้นใยรองรับที่อยู่ในชั้นไซโตพลาสซึมซึ่งติดกับเมมเบรนอย่างแน่นหนา พื้นผิวของลำตัวของรองเท้า ciliate มีขนจำนวนมาก (ประมาณ 15,000) ซึ่งผันผวนโดยไม่คำนึงถึงสถานการณ์ภายนอก ที่ฐานของพวกมันแต่ละตัวเป็นฐาน ตาเคลื่อนที่ประมาณ 30 ครั้งต่อวินาทีเพื่อดันร่างกายไปข้างหน้า สิ่งสำคัญคือต้องสังเกตว่าการเคลื่อนไหวที่เหมือนคลื่นของอุปกรณ์เหล่านี้มีการประสานกันอย่างมาก ซึ่งช่วยให้ infusoria หมุนช้าๆ และสวยงามรอบแกนตามยาวของร่างกายในกระบวนการเคลื่อนไหว
Ciliates - ประเภทของดอกเบี้ยที่แน่นอน
เพื่อให้เข้าใจถึงคุณสมบัติทั้งหมดของรองเท้า ciliates ขอแนะนำให้พิจารณากระบวนการหลักของชีวิต ดังนั้นการกินแบคทีเรียและสาหร่ายจึงลงมา ร่างกายของสิ่งมีชีวิตนั้นมีช่องที่เรียกว่าปากเซลล์และผ่านเข้าไปในคอหอยที่ด้านล่างของอาหารเข้าสู่ vacuole โดยตรง ที่นั่นจะถูกย่อยเป็นเวลาประมาณหนึ่งชั่วโมง ทำให้กระบวนการเปลี่ยนจากสภาพแวดล้อมที่เป็นกรดไปเป็นด่างในกระบวนการ แวคิวโอลเคลื่อนตัวในร่างกายของซิลิเอตผ่านการไหลของไซโตพลาสซึม และสารตกค้างที่ไม่ได้แยกแยะออกไปทางด้านหลังร่างกายผ่านผง
การหายใจของรองเท้า ciliates ดำเนินการโดยออกซิเจนเข้าสู่ไซโตพลาสซึมผ่านผิวหนังของร่างกาย และกระบวนการขับถ่ายเกิดขึ้นผ่านสองแวคิวโอลที่หดตัว สำหรับความหงุดหงิดของสิ่งมีชีวิต รองเท้า ciliates มักจะรวมตัวกันเป็นคอมเพล็กซ์ของแบคทีเรียเพื่อตอบสนองต่อการกระทำของสารที่หลั่งออกมาจากแบคทีเรีย และพวกมันก็ลอยจากสิ่งระคายเคืองเช่นเกลือแกง
การสืบพันธุ์

ciliate รองเท้าสามารถทำซ้ำได้สองวิธี การสืบพันธุ์แบบไม่อาศัยเพศแพร่หลายมากขึ้นตามที่นิวเคลียสแบ่งออกเป็นสองส่วน จากการดำเนินการนี้ แต่ละ ciliate มี 2 นิวเคลียส (ใหญ่และเล็ก) การสืบพันธุ์แบบอาศัยเพศมีความเหมาะสมเมื่อมีภาวะขาดสารอาหารหรือการเปลี่ยนแปลงของอุณหภูมิในร่างกายของสัตว์ ควรสังเกตว่าหลังจากนี้ infusoria สามารถกลายเป็นถุงน้ำได้ แต่ด้วยลักษณะการสืบพันธุ์แบบอาศัยเพศ จึงไม่นับการเพิ่มจำนวนบุคคล ดังนั้น ciliates สองตัวจึงเชื่อมต่อกันในช่วงระยะเวลาหนึ่งอันเป็นผลมาจากการที่เปลือกละลายและสะพานเชื่อมต่อระหว่างสัตว์ต่างๆ มันเป็นสิ่งสำคัญที่นิวเคลียสขนาดใหญ่ของพวกมันแต่ละตัวจะหายไปอย่างไร้ร่องรอย และนิวเคลียสขนาดใหญ่ของพวกมันแต่ละตัวจะต้องผ่านกระบวนการฟิชชันสองครั้ง ดังนั้นในแต่ละ ciliate จะมีการสร้างนิวเคลียสลูกสาว 4 ตัวหลังจากนั้นสามตัวจะถูกทำลายและส่วนที่สี่จะถูกแบ่งอีกครั้ง กระบวนการทางเพศนี้เรียกว่าการผันคำกริยา และระยะเวลาอาจนานถึง 12 ชั่วโมง
รองเท้า infusoria อาศัยอยู่ในอ่างเก็บน้ำขนาดเล็กที่นิ่ง สัตว์เซลล์เดียวตัวนี้ ยาว 0.5 มม. มีลำตัวเป็นแกนหมุน คล้ายกับรองเท้า Ciliates เคลื่อนไหวตลอดเวลาว่ายน้ำโดยให้ปลายทู่ไปข้างหน้า ความเร็วในการเคลื่อนที่ของสัตว์ตัวนี้ถึง 2.5 มม. ต่อวินาที บนพื้นผิวของร่างกายพวกมันมีออร์แกเนลล์ของการเคลื่อนไหว - cilia มีสองนิวเคลียสในเซลล์: นิวเคลียสขนาดใหญ่มีหน้าที่เกี่ยวกับโภชนาการ การหายใจ การเคลื่อนไหว เมแทบอลิซึม นิวเคลียสขนาดเล็กมีส่วนร่วมในกระบวนการทางเพศ
โครงสร้างของรองเท้า infusoria
สิ่งมีชีวิตของ ciliates นั้นซับซ้อนกว่า เปลือกยืดหยุ่นบางที่หุ้มด้านนอกของ ciliate ช่วยรักษารูปร่างของร่างกายให้คงที่ นอกจากนี้ยังอำนวยความสะดวกด้วยเส้นใยรองรับที่พัฒนามาอย่างดีซึ่งตั้งอยู่ในชั้นไซโตพลาสซึมที่อยู่ติดกับเปลือก ซิลิเอตสั่นประมาณ 15,000 ซี่ตั้งอยู่บนผิวลำตัวของซิลิเอต ที่ฐานของแต่ละซีลีเนียมจะมีร่างกายเป็นฐาน การเคลื่อนไหวของขนตาแต่ละข้างประกอบด้วยการลากเส้นที่แหลมคมในทิศทางเดียว และค่อยๆ กลับคืนสู่ตำแหน่งเดิมที่นุ่มนวลขึ้น ตาจะสั่นประมาณ 30 ครั้งต่อวินาที และเหมือนพาย ดัน infusoria ไปข้างหน้า การเคลื่อนไหวเหมือนคลื่นของ cilia นั้นประสานกัน เมื่อรองเท้า ciliate แหวกว่าย มันจะค่อยๆ หมุนรอบแกนตามยาวของร่างกาย
กระบวนการชีวิต
โภชนาการ
รองเท้าแตะและ ciliates ที่มีชีวิตอิสระบางชนิดกินแบคทีเรียและสาหร่าย
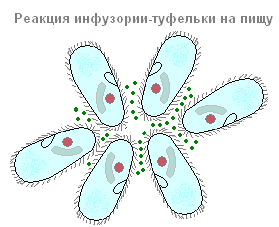
ปฏิกิริยาของรองเท้า ciliates ต่ออาหาร
เปลือกยืดหยุ่นบาง ( เยื่อหุ้มเซลล์) ปกปิด ciliate จากภายนอก รักษารูปร่างให้คงที่ ประมาณ 15,000 cilia อยู่บนพื้นผิวของร่างกาย ในร่างกายมีช่อง - ปากเซลล์ซึ่งผ่านเข้าไปในคอหอยเซลล์ ที่ด้านล่างของคอหอย อาหารจะเข้าสู่ vacuole ย่อยอาหาร ในแวคิวโอลย่อยอาหาร อาหารจะถูกย่อยภายในหนึ่งชั่วโมง ขั้นแรกด้วยกรดและปฏิกิริยาอัลคาไลน์ แวคิวโอลทางเดินอาหารเคลื่อนที่ในร่างกายของ ciliates โดยกระแสของไซโตพลาสซึม สารตกค้างที่ไม่ได้แยกแยะจะถูกโยนออกที่ส่วนท้ายของร่างกายผ่านโครงสร้างพิเศษ - ผงที่อยู่ด้านหลังช่องเปิดปาก
ลมหายใจ
การหายใจเกิดขึ้นผ่านทางผิวหนังของร่างกาย ออกซิเจนเข้าสู่ไซโตพลาสซึมผ่านพื้นผิวทั้งหมดของร่างกายและออกซิไดซ์สารอินทรีย์ที่ซับซ้อนซึ่งเป็นผลมาจากการที่พวกมันกลายเป็นน้ำคาร์บอนไดออกไซด์และสารประกอบอื่น ๆ ในขณะเดียวกัน พลังงานก็ถูกปล่อยออกมาซึ่งจำเป็นต่อชีวิตของสัตว์ คาร์บอนไดออกไซด์ถูกกำจัดออกไปทั่วทั้งร่างกายระหว่างการหายใจ
การคัดเลือก
ในร่างกายของรองเท้า ciliates มี vacuoles หดตัวสองอันซึ่งตั้งอยู่ที่ส่วนหน้าและส่วนหลังของร่างกาย พวกเขารวบรวมน้ำที่มีสารละลายที่เกิดขึ้นในระหว่างการออกซิเดชันของสารอินทรีย์ที่ซับซ้อน เมื่อถึงค่าขีด จำกัด แล้ว vacuoles ที่หดตัวเข้าหาพื้นผิวของร่างกายและเนื้อหาก็ไหลออกมา ในสัตว์น้ำจืดที่มีเซลล์เดียว น้ำส่วนเกินจะถูกลบออกผ่านทางแวคิวโอลที่หดตัว ซึ่งเข้าสู่ร่างกายของพวกมันจากสิ่งแวดล้อมอย่างต่อเนื่อง
ความหงุดหงิด
รองเท้า Ciliates รวมตัวกันเป็นกลุ่มแบคทีเรียเพื่อตอบสนองต่อการกระทำของสารที่หลั่งออกมา แต่ว่ายน้ำให้ห่างจากสารระคายเคืองเช่นเกลือแกง
ความหงุดหงิดเป็นคุณสมบัติของสิ่งมีชีวิตทุกชนิดที่ตอบสนองต่อการกระทำของสิ่งเร้า - แสง ความร้อน ความชื้น สารเคมี อิทธิพลทางกล ต้องขอบคุณความหงุดหงิดที่ทำให้สัตว์ที่มีเซลล์เดียวหลีกเลี่ยงสภาวะที่ไม่พึงประสงค์ ค้นหาอาหาร บุคคลแห่งปีของพวกมัน
การสืบพันธุ์
กะเทย
Ciliates มักจะสืบพันธุ์แบบไม่อาศัยเพศโดยแบ่งเป็นสองส่วน นิวเคลียสถูกแบ่งออกเป็นสองส่วน และซิลิเอตใหม่แต่ละอันประกอบด้วยนิวเคลียสขนาดใหญ่หนึ่งอันและนิวเคลียสขนาดเล็กหนึ่งอัน ลูกสาวสองคนแต่ละคนได้รับส่วนหนึ่งของออร์แกเนลล์ ในขณะที่คนอื่นๆ จะก่อตัวขึ้นใหม่
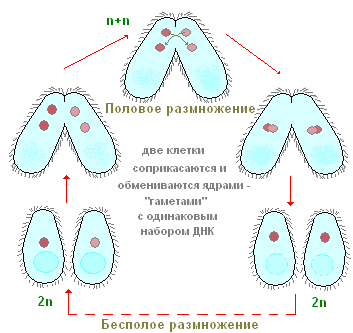
การสืบพันธุ์ของ ciliates-shoes
ทางเพศ
เมื่อขาดอาหารหรืออุณหภูมิเปลี่ยนแปลง ciliates ไปสู่การสืบพันธุ์แบบอาศัยเพศและจากนั้นก็จะกลายเป็นซีสต์ได้
ในระหว่างกระบวนการทางเพศจะไม่เพิ่มจำนวนบุคคล ciliates สองอันเชื่อมต่อกันชั่วคราว เมื่อสัมผัสกัน เปลือกจะละลายและเกิดเป็นสะพานเชื่อมระหว่างสัตว์ นิวเคลียสขนาดใหญ่ของแต่ละ ciliate หายไป นิวเคลียสขนาดเล็กแบ่งสองครั้ง ในแต่ละ ciliate จะเกิดนิวเคลียสลูกสาวสี่ตัว สามคนถูกทำลายและคนที่สี่ถูกแบ่งอีกครั้ง เป็นผลให้แกนทั้งสองยังคงอยู่ในแต่ละแกน การแลกเปลี่ยนนิวเคลียร์เกิดขึ้นตามสะพานไซโตพลาสซึมและรวมเข้ากับนิวเคลียสที่เหลือที่นั่น นิวเคลียสที่ก่อตัวใหม่ก่อตัวเป็นนิวเคลียสขนาดใหญ่และขนาดเล็ก และซิลิเอตแยกจากกัน กระบวนการทางเพศนี้เรียกว่าการผันคำกริยา ใช้เวลาประมาณ 12 ชั่วโมง กระบวนการทางเพศนำไปสู่การต่ออายุ การแลกเปลี่ยนระหว่างปัจเจกบุคคล และการแจกจ่ายสารพันธุกรรม (พันธุกรรม) ซึ่งเพิ่มความมีชีวิตของสิ่งมีชีวิต
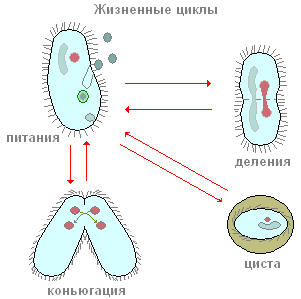
วงจรชีวิตของรองเท้า ciliates
นอกจากนี้เรายังแนะนำ
 ความคิดที่มีประสิทธิผลและการเจริญพันธุ์
ความคิดที่มีประสิทธิผลและการเจริญพันธุ์
 ความเห็นแก่ตัวที่สมเหตุสมผล - ทฤษฎีความเห็นแก่ตัวที่สมเหตุสมผลคืออะไร?
ความเห็นแก่ตัวที่สมเหตุสมผล - ทฤษฎีความเห็นแก่ตัวที่สมเหตุสมผลคืออะไร?
 Boris Nikolaevich Yeltsin ประธานาธิบดีคนแรกของรัสเซีย
Boris Nikolaevich Yeltsin ประธานาธิบดีคนแรกของรัสเซีย
 การต่อสู้ใต้ดิน ราชาใต้ดิน. อะไรคือ "การต่อสู้ไม่ใช่เพื่อมวลชน"? คุณจะต่อสู้เพื่อเงินได้ที่ไหน?
การต่อสู้ใต้ดิน ราชาใต้ดิน. อะไรคือ "การต่อสู้ไม่ใช่เพื่อมวลชน"? คุณจะต่อสู้เพื่อเงินได้ที่ไหน?
 Yakov Pavlov และวีรบุรุษคนอื่น ๆ ของ Stalingrad ที่คุณต้องรู้
Yakov Pavlov และวีรบุรุษคนอื่น ๆ ของ Stalingrad ที่คุณต้องรู้
 เอาชีวิตรอดจากอุบัติเหตุกลางทะเลในฝัน - สัมผัสรักครั้งใหม่ในชีวิตจริง
เอาชีวิตรอดจากอุบัติเหตุกลางทะเลในฝัน - สัมผัสรักครั้งใหม่ในชีวิตจริง