บริเวณหน้าที่ของ tRNA โครงสร้างและหน้าที่ของ tRNA คุณสมบัติของการกระตุ้นกรดอะมิโน
70-90N | หน้ารอง - cloverleaf | CCA 3" const สำหรับ tRNA ทั้งหมด |
การปรากฏตัวของ thymine, pseudouridine-psi, digirouridine DGU ใน D-loop - ป้องกันไรโบนิวคลีเอส? อายุยืนยาว | โครงสร้างหลักของ tRNA ที่หลากหลาย - 61 + 1 - โดยจำนวนของ codon + formylmethionine tRNA แอนติโคดอนของแมวจะเหมือนกับของเมไทโอนีน tRNA ความหลากหลายของโครงสร้างตติยภูมิ - 20 (ตามจำนวนกรดอะมิโน) | การรับรู้ - การก่อตัวของพันธะโควาเลนต์ m-y tRNA และกระทำ | การสังเคราะห์ aminoacyl-tRNA แนบการกระทำกับ tRNA
หน้าที่ของ tRNA คือการถ่ายโอนกรดอะมิโนจากไซโตพลาสซึมไปยังไรโบโซมซึ่งเกิดการสังเคราะห์โปรตีน
tRNAs ที่จับกรดอะมิโนหนึ่งตัวเรียกว่าไอโซแอคเซพเตอร์
โดยรวมแล้ว มี tRNA ที่แตกต่างกัน 64 ตัวในเซลล์
tRNA แต่ละตัวจับคู่กับ codon ของตัวเองเท่านั้น
แต่ละ tRNA รับรู้ codon ของตัวเองโดยไม่เกี่ยวข้องกับกรดอะมิโน กรดอะมิโนที่จับกับ tRNA ถูกดัดแปลงทางเคมี หลังจากนั้นโพลีเปปไทด์ที่เป็นผลลัพธ์ ซึ่งมีกรดอะมิโนดัดแปลง ถูกวิเคราะห์ Cysteinyl-tRNACys (R=CH2-SH) ลดลงเป็น alanyl-tRNACys (R=CH3)
tRNA ส่วนใหญ่โดยไม่คำนึงถึงลำดับนิวคลีโอไทด์มีโครงสร้างทุติยภูมิรูปใบโคลเวอร์ลีฟเนื่องจากมีกิ๊บติดผมสามอัน
คุณสมบัติโครงสร้างของ tRNA
มีนิวคลีโอไทด์ที่ไม่ได้จับคู่สี่ตัวเสมอที่ส่วนท้ายของโมเลกุล 3 "และสามในนั้นจำเป็นต้องมี CCA ปลาย 5" และ 3 "ของสาย RNA ก่อให้เกิดก้านตัวรับ โซ่ถูกยึดเข้าด้วยกันเนื่องจากการจับคู่เสริมของ นิวคลีโอไทด์เจ็ดตัว 5" - สิ้นสุดด้วยนิวคลีโอไทด์เจ็ดตัวที่อยู่ใกล้กับจุดสิ้นสุด 3 " 2. โมเลกุลทั้งหมดมีกิ๊บติดผม T? C ดังนั้นจึงกำหนดไว้เพราะมันประกอบด้วยสารตกค้างที่ผิดปกติสองชนิด: ไรโบไทมิดีน (T) และซูโดริดีน (? กิ๊บประกอบด้วยสองเท่า - ก้านเกลียวที่มีฐานห้าคู่ รวมทั้งคู่ G-C และวงยาวเจ็ดนิวคลีโอไทด์
ที่จุดเดียวกันในวง 3. ในกิ๊บติดผมแอนติโคดอน ก้านจะเป็นตัวแทนของครอบครัวที่จับคู่เสมอ
บริเวณ Triplet ประกอบกับ codon ที่เกี่ยวข้อง, anticodon ตั้งอยู่ในลูป
le ประกอบด้วยนิวคลีโอไทด์เจ็ดตัว ค่าคงที่ ura-
cyl และ cytosine ที่ดัดแปลงและ purine ที่ดัดแปลงจะติดกับปลาย 3 "ตามกฎ
อะดีนีน 4. กิ๊บติดผมอีกอันประกอบด้วยก้านนิวคลีโอไทด์ยาวสามถึงสี่คู่และลูปผันแปร
ขนาดมักประกอบด้วย uracil ในรูปแบบย่อ - dihydrouracil (DU) ลำดับนิวคลีโอไทด์ของก้าน จำนวนของนิวคลีโอไทด์ระหว่างก้านแอนติโคดอนและต้นกำเนิด T?C (ลูปแบบแปรผัน) ตลอดจนขนาดของลูปและการแปลตำแหน่งของเรซิดิวไดไฮโดรยูราซิลในลูป DU แตกต่างกันอย่างมาก
[นักร้อง, 1998].
โครงสร้างตติยของ tRNA
โครงสร้างรูปตัว L
การเกาะติดของกรดอะมิโนกับ tRNA
เพื่อให้กรดอะมิโนก่อตัวเป็นสายโพลีเปปไทด์ จะต้องยึดติดกับ tRNA โดยเอ็นไซม์ aminoacyl-tRNA synthetase เอนไซม์นี้สร้างพันธะโควาเลนต์ระหว่างหมู่คาร์บอกซิลของกรดอะมิโนและกลุ่มไรโบสไฮดรอกซิลที่ส่วนท้าย 3' ของ tRNA โดยมีส่วนร่วมของ ATP Aminoacyl-tRNA synthetase รับรู้ codon เฉพาะ ไม่ใช่เพราะมี anticodon บน tRNA แต่โดยการมีอยู่ของตำแหน่งการจดจำเฉพาะบน tRNA
โดยรวมแล้ว มีการสังเคราะห์ aminoacyl-tRNA ที่แตกต่างกัน 21 ชนิดในเซลล์
การเข้าร่วมเกิดขึ้นในสองขั้นตอน:
1. หมู่คาร์บอกซิลของกรดอะมิโนติดอยู่กับเอทีพี เอ-ฟอสเฟต อะมิโนอะซิลอะดีนิเลตที่ไม่เสถียรที่ได้นั้นถูกทำให้เสถียรโดยการจับกับเอนไซม์
2. การถ่ายโอนหมู่อะมิโนอะซิลของอะมิโนอะซิลอะดีนิเลตไปยังกลุ่ม 2' หรือ 3'-OH ของไรโบสปลายทางของ tRNA
การสังเคราะห์อะมิโนอะซิล-tRNA บางชนิดประกอบด้วยสายโซ่พอลิเปปไทด์เดี่ยว ในขณะที่บางชนิดประกอบด้วยสายโซ่ที่เหมือนกันสองหรือสี่สาย โดยแต่ละสายมีน้ำหนักโมเลกุล 35 ถึง 115 kDa เอนไซม์ dimeric และ tetrameric บางชนิดประกอบด้วยหน่วยย่อยสองประเภท ไม่มีความสัมพันธ์ที่ชัดเจนระหว่างขนาดของโมเลกุลของเอนไซม์หรือลักษณะของโครงสร้างหน่วยย่อยและความจำเพาะ
ความจำเพาะของเอ็นไซม์ถูกกำหนดโดยการจับอย่างแรงกับปลายตัวรับของ tRNA, ขอบเขต DU และลูปตัวแปร เอ็นไซม์บางตัวไม่รู้จักแอนติโคดอนทริปเปิ้ลและกระตุ้นปฏิกิริยาอะมิโนอะซิติเลชันแม้ว่าแอนติโคดอนจะเปลี่ยนแปลงไปก็ตาม อย่างไรก็ตาม เอนไซม์บางตัวแสดงกิจกรรมที่ลดลงซึ่งสัมพันธ์กับ tRNA ที่ถูกดัดแปลงดังกล่าว และเพิ่มกรดอะมิโนที่ไม่ถูกต้องเมื่อแทนที่แอนติโคดอน
70-90n | หน้ารอง - cloverleaf | CCA 3" const สำหรับ tRNA ทั้งหมด |
การปรากฏตัวของ thymine, pseudouridine-psi, digirouridine DGU ใน D-loop - ป้องกันไรโบนิวคลีเอส? อายุยืนยาว | โครงสร้างหลักของ tRNA ที่หลากหลาย - 61 + 1 - โดยจำนวนของ codon + formylmethionine tRNA แอนติโคดอนของแมวจะเหมือนกับของเมไทโอนีน tRNA ความหลากหลายของโครงสร้างตติยภูมิ - 20 (ตามจำนวนกรดอะมิโน)
มีสองประเภทของ tRNA ที่จับ methionine tRNAFMet และ tRNAMMet ในโปรคาริโอตและ tRNAIMet และ tRNAMMet ในยูคาริโอต เติมเมไทโอนีนในแต่ละ tRNA โดยใช้การสังเคราะห์ aminoacyl-tRNA ที่เหมาะสม เมไทโอนีนที่ติดอยู่กับ tRNAFMet และ tRNAIMet เกิดจากเอนไซม์ methionyl-tRNA-transformylase กับ Fmet-tRNAFMet tRNA ที่บรรจุฟอร์มิลเมไทโอนีนรับรู้การเริ่มต้น codon AUG
วรรณกรรม:
น่าเสียดายที่ไม่มีบรรณานุกรม
หนังสือเรียน. แม้ว่าข้อเท็จจริงที่ว่า tRNA จะเล็กกว่ามาก แต่เรื่องราวเกี่ยวกับโครงสร้าง คุณลักษณะ และการทำงานของมันสมควรได้รับบทที่แยกจากกัน
ดังนั้น tRNA จึงเป็น “อะแดปเตอร์” ซึ่งจดจำลำดับตัวอักษรสามตัวของรหัสพันธุกรรมที่ปลายด้านหนึ่ง โดยจับคู่กับกรดอะมิโนที่สอดคล้องกันเพียงตัวเดียวซึ่งจับจ้องอยู่ที่ปลายอีกด้านหนึ่งของ tRNA ในตอนท้ายของการถ่ายโอน RNA ที่สัมผัสกับ RNA ของผู้ส่งสาร จะมีนิวคลีโอไทด์ 3 ตัวที่ก่อตัว แอนติโคดอน. เฉพาะในกรณีที่แอนติโคดอนเป็นส่วนเสริมของภูมิภาค mRNA เท่านั้นจึงจะสามารถถ่ายโอน RNA เข้าร่วมได้ แต่ถึงกระนั้นในกรณีนี้ tRNA ก็ไม่สามารถเข้าร่วม mRNA ได้ด้วยตัวเอง มันต้องการความช่วยเหลือจากไรโบโซม ซึ่งเป็นที่ตั้งของปฏิสัมพันธ์ของพวกมัน เช่นเดียวกับผู้เข้าร่วมการแปลอย่างแข็งขัน ตัวอย่างเช่น เป็นไรโบโซมที่สร้างพันธะระหว่างกรดอะมิโนที่นำโดย tRNA ทำให้เกิดสายโปรตีน
ลักษณะโครงสร้างของ tRNA ถูกกำหนดโดยรหัสพันธุกรรม กล่าวคือ กฎสำหรับการสร้างโปรตีนตามยีนที่ RNA การถ่ายโอนอ่าน รหัสนี้ใช้ได้กับทุกสิ่งมีชีวิตบนโลก: การสร้างไวรัสนั้นเขียนด้วย codon สามตัวอักษรเดียวกันกับที่ใช้เขียน "คำแนะนำในการประกอบ" ของโลมา ได้รับการตรวจสอบจากการทดลองแล้วว่ายีนของสิ่งมีชีวิตตัวหนึ่งซึ่งอยู่ในเซลล์ของอีกตัวหนึ่งนั้นถูกคัดลอกและแปลอย่างสมบูรณ์เป็นโปรตีนที่แยกไม่ออกจากยีนที่สืบพันธุ์ในเซลล์ของโฮสต์ ความสม่ำเสมอของรหัสพันธุกรรมเป็นพื้นฐานสำหรับการผลิตอีโคไลดัดแปลงโดยอาณานิคมของอินซูลินและเอ็นไซม์อื่น ๆ ของมนุษย์ที่ใช้เป็นยาสำหรับผู้ที่ร่างกายไม่สามารถผลิตได้หรือผลิตได้ไม่เพียงพอ แม้จะมีความแตกต่างที่ชัดเจนระหว่างมนุษย์และอี. โคไล โปรตีนของมนุษย์ก็ถูกสร้างขึ้นได้อย่างง่ายดายจากพิมพ์เขียวของมนุษย์โดยใช้เครื่องถ่ายเอกสาร E. coli ไม่น่าแปลกใจเลยที่ RNA การถ่ายโอนของสิ่งมีชีวิตต่าง ๆ ต่างกันเพียงเล็กน้อย

แต่ละ codon จากรายการนี้ ยกเว้นสาม หยุด codonsการถ่ายโอน RNA ควรรับรู้โดยส่งสัญญาณว่าการแปลเสร็จสิ้น การรับรู้ทำได้โดยการแนบแอนติโคดอนเข้ากับ RNA ของผู้ส่งสาร ซึ่งสามารถผูกกับโคดอนเพียงตัวเดียวจากรายการ ดังนั้น tRNA สามารถจดจำโคดอนได้เพียงตัวเดียวเท่านั้น ซึ่งหมายความว่ามีโมเลกุลเหล่านี้อย่างน้อย 61 ชนิดในเซลล์ ในความเป็นจริง มีมากกว่านั้น เนื่องจากในบางสถานการณ์สำหรับการอ่าน RNA ของผู้ส่งสาร การมีแอนติโคดอนที่ถูกต้องไม่เพียงพอเพียงต้องมีเงื่อนไขอื่น ๆ ซึ่งจำเป็นต้องมีการสร้าง tRNA แบบพิเศษที่ดัดแปลงขึ้นมา
เมื่อมองแวบแรก tRNA ที่หลากหลายดังกล่าวควรทำให้กระบวนการแปลซับซ้อนขึ้นอย่างมีนัยสำคัญ: ท้ายที่สุดแล้ว แต่ละโมเลกุลเหล่านี้จะตรวจสอบเมทริกซ์ RNA codon ที่แทนที่ด้วยไรโบโซมเพื่อให้สอดคล้องกับแอนติโคดอนของมัน - ดูเหมือนว่างานทางกลที่ไร้จุดหมายมาก เสียเวลาและพลังงานไปมาก แต่เป็นผลมาจากวิวัฒนาการ กลไกของเซลล์ได้ถูกสร้างขึ้นที่ป้องกันปัญหานี้เช่นกัน ตัวอย่างเช่น ปริมาณของ tRNA ของแต่ละสปีชีส์ในเซลล์สอดคล้องกับความถี่ที่กรดอะมิโนที่สปีชีส์นั้นพบในโปรตีนที่ถูกสร้างขึ้น มีกรดอะมิโนที่เซลล์ไม่ค่อยได้ใช้ และยังมีกรดอะมิโนที่มักใช้ และถ้าจำนวน tRNA ที่บรรทุกพวกมันเท่ากัน จะทำให้การประกอบโปรตีนยุ่งยากมาก ดังนั้นจึงมีกรดอะมิโนที่ "หายาก" และ tRNAs ที่สอดคล้องกันในเซลล์เพียงไม่กี่ชนิด ในขณะที่กรดอะมิโนที่เกิดขึ้นบ่อยครั้งจะถูกสร้างขึ้นในปริมาณมาก

ด้วยโมเลกุล tRNA ที่หลากหลาย พวกมันทั้งหมดจึงคล้ายกันมาก ดังนั้น เมื่อพิจารณาถึงโครงสร้างและหน้าที่ของพวกมัน เราจะศึกษาลักษณะเฉพาะของสัตว์ทุกชนิดเป็นหลัก เมื่อคุณดูที่เลย์เอาต์ 3 มิติของ tRNA ดูเหมือนอะตอมที่หนาแน่น ดูน่าเหลือเชื่อที่โมเลกุลที่ขดอย่างประณีตนี้เป็นผลมาจากการพับของนิวคลีโอไทด์สายโซ่ยาว แต่นั่นเป็นวิธีที่มันเกิดขึ้น
เป็นไปได้ที่จะติดตามขั้นตอนของการก่อตัวของมันตั้งแต่แรก: การรวบรวมลำดับนิวคลีโอไทด์โดย RNA polymerase ตามยีนที่มีข้อมูลเกี่ยวกับการถ่ายโอน RNA ลำดับที่นิวคลีโอไทด์เหล่านี้ติดตามกันและเรียกหมายเลข โครงสร้างหลักของ tRNA. ปรากฎว่าเป็นโครงสร้างหลักของ tRNA ที่เข้ารหัสในยีนที่อ่านโดย RNA polymerase โดยทั่วไป โครงสร้างหลักคือลำดับของโมเลกุลที่ค่อนข้างง่ายในประเภทเดียวกัน ซึ่งประกอบขึ้นด้วยโมเลกุลโพลีเมอร์ที่พับซ้อนและซับซ้อนกว่า ตัวอย่างเช่น โครงสร้างหลักของโมเลกุลโปรตีนคือลำดับอย่างง่ายของกรดอะมิโนที่เป็นส่วนประกอบ

สายโซ่ของนิวคลีโอไทด์ไม่สามารถอยู่ในสถานะที่กางออกในเซลล์ได้ เพียงแค่ยืดออกเป็นเส้น มีชิ้นส่วนที่มีประจุบวกและลบมากเกินไปที่ขอบของนิวคลีโอไทด์ ซึ่งสร้างพันธะไฮโดรเจนซึ่งกันและกันได้ง่าย มีการอธิบายวิธีการสร้างพันธะเดียวกันระหว่างนิวคลีโอไทด์ของโมเลกุล DNA สองโมเลกุลที่เชื่อมต่อกันเป็นเกลียวคู่ และคุณสามารถดูรายละเอียดเกี่ยวกับพันธะไฮโดรเจนได้ พันธะไฮโดรเจนมีความแข็งแรงน้อยกว่าพันธะระหว่างอะตอมในโมเลกุล แต่ก็เพียงพอที่จะบิดเกลียว tRNA อย่างประณีตและเก็บไว้ในตำแหน่งนั้น ในตอนแรก พันธะเหล่านี้เกิดขึ้นระหว่างนิวคลีโอไทด์บางตัวเท่านั้น โดยพับ tRNA ให้เป็นรูปใบโคลเวอร์ ผลของการพับครั้งแรกนี้เรียกว่า โครงสร้างรอง tRNA. แผนภาพทางด้านซ้ายแสดงให้เห็นว่ามีเพียงนิวคลีโอไทด์บางตัวเท่านั้นที่เชื่อมโยงกันด้วยพันธะไฮโดรเจน ในขณะที่บางตัวยังคงไม่จับคู่กัน ทำให้เกิดวงแหวนและลูป ความแตกต่างระหว่างโครงสร้างทุติยภูมิของ tRNA ประเภทต่างๆ เกิดจากความแตกต่างในโครงสร้างหลัก สิ่งนี้แสดงให้เห็นในความยาวที่แตกต่างกันของ "ใบโคลเวอร์" หรือ "ก้าน" เนื่องจากความยาวต่างกันของสายโซ่เริ่มต้นของนิวคลีโอไทด์
ความแตกต่างอีกประการในโครงสร้างหลักของ tRNA ที่ต่างกันคือมีนิวคลีโอไทด์เหมือนกันในบางตำแหน่งเท่านั้น (ในแผนภาพด้านบนจะมีตัวอักษรตัวแรกของชื่อกำกับอยู่) ในขณะที่นิวคลีโอไทด์ส่วนใหญ่ใน tRNA ต่างกันต่างกัน รูปแบบข้างต้นเป็นเรื่องปกติสำหรับ tRNA ทั้งหมด ดังนั้นนิวคลีโอไทด์ที่แตกต่างกันจึงถูกทำเครื่องหมายด้วยตัวเลข
ส่วนหน้าที่หลักของ tRNA คือ:
=) แอนติโคดอนนั่นคือลำดับนิวคลีโอไทด์ที่ประกอบกับ codon เดียวของ messenger RNA ที่อยู่บน กิ๊บติดผมแอนติโคดอน
=) ปลายตัวรับซึ่งสามารถติดกรดอะมิโนได้ ตั้งอยู่ฝั่งตรงข้ามของกิ๊บติดผมแอนติโคดอน
ในความเป็นจริง ไม่มี tRNA เดียวที่ดูเหมือนในแผนภาพโครงสร้างทุติยภูมิ เพราะมีเพียงนิวคลีโอไทด์บางตัวเท่านั้นที่รวมตัวกันเพื่อสร้างมัน ในขณะที่ส่วนที่เหลือยังคงไม่มีการจับคู่ เนื่องจากการก่อตัวของพันธะไฮโดรเจนระหว่างนิวคลีโอไทด์จากส่วนต่าง ๆ ของใบโคลเวอร์ มันพับเพิ่มเติมเข้าไปซับซ้อนมากขึ้น โครงสร้างระดับอุดมศึกษาในรูปตัว L คุณสามารถเข้าใจได้อย่างชัดเจนว่าส่วนต่างๆ ของโครงสร้างทุติยภูมิโค้งอย่างไรเพื่อสร้างโครงสร้างระดับอุดมศึกษาด้วยการจับคู่สีในไดอะแกรมด้านล่าง กิ๊บติดผม anticodon ซึ่งทำเครื่องหมายด้วยสีน้ำเงินและสีเทายังคงอยู่ที่ด้านล่าง (ควรจำไว้ว่า "ด้านล่าง" นี้มีเงื่อนไข: สะดวกในการพรรณนา tRNA ในการวางแนวเชิงพื้นที่ในรูปแบบการแปลโปรตีน) และจุดสิ้นสุดของตัวรับ (สีเหลือง) จะงอไปด้านข้าง

นี่คือลักษณะของ tRNA เมื่อพร้อมที่จะแนบกรดอะมิโน tRNA ไม่สามารถรวมกับกรดอะมิโนได้ด้วยตัวเอง จึงต้องอาศัยเอนไซม์พิเศษร่วมด้วย: การสังเคราะห์อะมิโนอะซิล-tRNA. จำนวนประเภทการสังเคราะห์ในเซลล์ตรงกับจำนวนของประเภท tRNA
ความสม่ำเสมอของรูปร่างของ tRNA ทุกประเภทเป็นสิ่งจำเป็นเพื่อให้ไรโบโซมสามารถจดจำพวกมันได้ อำนวยความสะดวกในการเชื่อมต่อด้วย mRNA และย้ายภายในตัวมันเองจากไซต์หนึ่งไปยังอีกที่หนึ่ง หาก tRNA ประเภทต่างๆ แตกต่างกันอย่างมีนัยสำคัญ จะทำให้การทำงานของไรโบโซมทำได้ยากอย่างยิ่ง ซึ่งจะช่วยลดอัตราการสังเคราะห์โปรตีนในขั้นวิกฤต การคัดเลือกโดยธรรมชาติจึงมีจุดมุ่งหมายเพื่อทำให้ tRNAs มีความคล้ายคลึงกัน แต่ในขณะเดียวกัน มีปัจจัยอื่นที่ต้องการการมีอยู่ของความแตกต่างที่เห็นได้ชัดเจนระหว่าง tRNA ประเภทต่างๆ: ท้ายที่สุด จำเป็นต้องรู้จักแต่ละประเภทและแนบกรดอะมิโนที่สอดคล้องกันเพียงตัวเดียวเข้ากับมัน เห็นได้ชัดว่าความแตกต่างเหล่านี้ควรสังเกตได้ชัดเจน แต่ไม่สำคัญเกินไป เพื่อให้งานในการจำแนกชนิดของ tRNA กลายเป็นกระบวนการเครื่องประดับ และนี่คือสิ่งที่กระทำโดย aminoacyl-tRNA synthetases อย่างแม่นยำ: แต่ละตัวสามารถจับกับกรดอะมิโนเพียงตัวเดียวจาก 20 ชนิดและยึดติดกับ tRNA ที่สอดคล้องกับกรดอะมิโนนี้อย่างแม่นยำ จากตารางที่มีรหัสพันธุกรรม จะเห็นได้ว่ากรดอะมิโนแต่ละชนิดถูกเข้ารหัสโดยลำดับนิวคลีโอไทด์หลายลำดับ ตัวอย่างเช่น tRNA ทั้งสี่ตัวที่มีแอนติโคดอน CGA, CGG, CGU และ CGC จะรับรู้โดยซินธิเทสเดียวกันกับอะลานีน ถึงพวกเขา. tRNA ดังกล่าวถูกประมวลผลโดยซินธิเทสเดียวเรียกว่า ที่เกี่ยวข้อง.
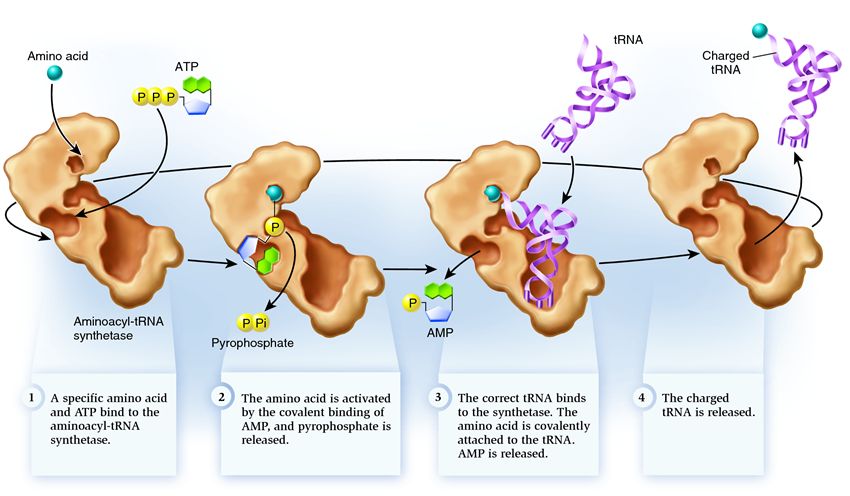
ซินธิเตสอยู่ในกลุ่มของเอนไซม์ที่ทำหน้าที่จับกับโมเลกุลที่มีอยู่แยกจากกันและรวมเข้าเป็นหนึ่งเดียว:
1 . ซินธิเทสเชื่อมต่อกรดอะมิโนกับโมเลกุล ATP กลุ่มฟอสเฟตสองกลุ่มแยกตัวออกจาก ATP โดยปล่อยพลังงานที่จำเป็นสำหรับกิจกรรมต่อไปนี้ อะดีโนซีนโมโนฟอสเฟต (AMP) ที่เหลืออยู่จากโมเลกุลที่ถูกทำลายจะเกาะติดกับกรดอะมิโน เพื่อเตรียมสำหรับการเชื่อมต่อกับกิ๊บของตัวรับ
2 . ซินธิเตสยึดติด tRNA ที่เกี่ยวข้องตัวใดตัวหนึ่งซึ่งสอดคล้องกับกรดอะมิโนนี้

ในขั้นตอนนี้จะมีการตรวจสอบความสอดคล้องของการถ่ายโอน RNA ไปยังการสังเคราะห์ มีหลายวิธีในการจดจำและแต่ละซินธิเตสมีการผสมผสานที่เป็นเอกลักษณ์ อย่างน้อยหนึ่งแอนติโคดอนนิวคลีโอไทด์มีส่วนเกี่ยวข้องในอันตรกิริยาระหว่างซินธิเทสและ tRNA ต้องตรวจสอบกิ๊บของตัวรับด้วย: การปรากฏตัวของนิวคลีโอไทด์จำเพาะบนมันซึ่งเป็นเรื่องปกติของ tRNA ที่เกี่ยวข้องซึ่งสอดคล้องกับกรดอะมิโนที่ต้องการจะถูกกำหนด นิวคลีโอไทด์จากส่วนอื่น ๆ ของ tRNA ยังสามารถมีส่วนร่วมในการจับคู่โดยจับกับไซต์สังเคราะห์บางตัว tRNA ที่ไม่ถูกต้องอาจตรงกับค่าที่ต้องการในทางใดทางหนึ่ง แต่เนื่องจากการจับคู่ที่ไม่สมบูรณ์ tRNA จะเข้าร่วมการสังเคราะห์อย่างช้าๆ และหลวม และหลุดออกมาได้ง่าย และ tRNA ที่ถูกต้องจะเกาะติดกับการสังเคราะห์อย่างรวดเร็วและแน่นหนาอันเป็นผลมาจากการเปลี่ยนแปลงโครงสร้างของการสังเคราะห์ที่เริ่มกระบวนการ อะมิโนอะซิเลชัน นั่นคือการเกาะติดของกรดอะมิโนกับ tRNA

3 . ประกอบด้วยการแทนที่โมเลกุล AMP ที่ติดอยู่กับกรดอะมิโนด้วยโมเลกุล tRNA หลังจากการแทนที่นี้ AMP จะออกจากการสังเคราะห์และ tRNA จะถูกกักไว้เพื่อตรวจสอบกรดอะมิโนครั้งสุดท้าย ถ้ารู้ว่ากรดอะมิโนที่ติดอยู่นั้นไม่ถูกต้อง จะถูกแยกออกจาก tRNA ตำแหน่งของกรดอะมิโนในการสังเคราะห์จะว่างเปล่า และโมเลกุลอื่นสามารถรวมกันที่นั่นได้ กรดอะมิโนใหม่จะผ่านขั้นตอนของการเชื่อมต่อกับ ATP และ tRNA และจะได้รับการทดสอบด้วย หากไม่มีข้อผิดพลาด tRNA ที่มีกรดอะมิโนจะถูกปล่อยออกมา: มันพร้อมที่จะมีบทบาทในการแปลโปรตีน และสารสังเคราะห์ก็พร้อมที่จะเกาะติดกับกรดอะมิโนและ tRNA ใหม่ และวัฏจักรก็จะเริ่มใหม่
มากขึ้นอยู่กับการทำงานที่ถูกต้องของการสังเคราะห์ aminoacyl-tRNA: หากความล้มเหลวเกิดขึ้นในขั้นตอนนี้ กรดอะมิโนที่ไม่ถูกต้องจะถูกแนบกับ tRNA และมันจะถูกสร้างขึ้นในโปรตีนที่เติบโตบนไรโบโซม เพราะ tRNA และไรโบโซมไม่มีหน้าที่ตรวจสอบความสอดคล้องของโคดอนและกรดอะมิโน ผลที่ตามมาจากความผิดพลาดอาจมีเพียงเล็กน้อยหรือเป็นหายนะ และโดยการคัดเลือกโดยธรรมชาติ สิ่งมีชีวิตที่มีเอนไซม์ที่ไม่มีหน้าที่ของการตรวจสอบดังกล่าว ได้ถูกแทนที่ด้วยสิ่งมีชีวิตที่ปรับเปลี่ยนได้ โดยมีตัวเลือกต่างๆ สำหรับการจับคู่ระหว่างกรดอะมิโนกับ tRNA ดังนั้นในเซลล์สมัยใหม่ การสังเคราะห์จึงรวมเข้ากับกรดอะมิโนที่ไม่ถูกต้องโดยเฉลี่ยในหนึ่งกรณีจาก 50,000 รายการ และด้วย tRNA ที่ไม่ถูกต้องเพียงครั้งเดียวใน 100,000 สิ่งที่แนบมา

กรดอะมิโนบางชนิดมีความแตกต่างกันเพียงไม่กี่อะตอม หากคุณดูแผนผังของมัน จะเห็นได้ชัดว่าโอกาสที่จะสับสนระหว่างอาร์จินีนกับอะลานีนนั้นน้อยกว่าไอโซลิวซีนที่ทำให้เกิดความสับสนสำหรับลิวซีนหรือวาลีน ดังนั้นแต่ละซินธิเตสที่จับกับกรดอะมิโนตัวใดตัวหนึ่งที่คล้ายคลึงกันจึงมีกลไกการตรวจสอบเพิ่มเติม นี่คือตัวอย่างของการดัดแปลงใน isoleucine-tRNA synthetase:

ซินเธสแต่ละตัวมี ศูนย์สังเคราะห์โดยที่กรดอะมิโนติดอยู่กับ tRNA กิ๊บตัวรับของ tRNA ที่จับได้โดยซินธิเทสจะไปที่นั่น เช่นเดียวกับกรดอะมิโนที่พร้อมจะทำปฏิกิริยากับมัน การทำงานของสารสังเคราะห์บางชนิดสิ้นสุดลงทันทีหลังจากการเชื่อมต่อของกรดอะมิโนและ tRNA แต่การสังเคราะห์ Ile-tRNA มีโอกาสเกิดข้อผิดพลาดเพิ่มขึ้นเนื่องจากการมีอยู่ของกรดอะมิโนคล้ายไอโซลิวซีนอื่นๆ ดังนั้นเธอจึงมี ราชทัณฑ์: จากชื่อ เห็นได้ชัดว่ามีบทบาทอย่างไรในกระบวนการเชื่อมต่อ tRNA และกรดอะมิโน รูปทางด้านขวาแสดงให้เห็นว่าตำแหน่งปลายของกิ๊บตัวรับ tRNA ในศูนย์กลางการสังเคราะห์ของการสังเคราะห์ Ile-tRNA ทำให้กิ๊บนี้โค้งงอผิดธรรมชาติ อย่างไรก็ตาม ซินธิเทสถือ tRNA ในตำแหน่งนี้จนกว่ากรดอะมิโนจะติดอยู่กับมัน หลังจากการเชื่อมต่อนี้เกิดขึ้น ความจำเป็นในการค้นหากิ๊บตัวรับในศูนย์สังเคราะห์หมดลง และ tRNA จะยืดออก โดยให้กรดอะมิโนติดอยู่ที่ศูนย์แก้ไข

แน่นอนว่าศูนย์สังเคราะห์ก็มีบทบาทในการคัดกรองกรดอะมิโนที่ไม่เหมาะสำหรับการสังเคราะห์ด้วย โมเลกุลต้องเป็นไปตามเงื่อนไขหลายประการรวมถึงการมีขนาดที่เหมาะสม แม้ว่าลิวซีนและไอโซลิวซีนมีจำนวนอะตอมเท่ากัน เนื่องจากความแตกต่างในโครงสร้างเชิงพื้นที่ ลิวซีนมีขนาดใหญ่กว่า ดังนั้นจึงไม่สามารถเจาะเข้าไปในศูนย์สังเคราะห์ซึ่งมีขนาดเท่ากับไอโซลิวซีนที่มีขนาดกะทัดรัดกว่า และกระเด้งออกจากการสังเคราะห์ Ile-tRNA
แต่วาลีนซึ่งเป็นโมเลกุลที่เล็กที่สุดในสามโมเลกุลนี้มีโครงสร้างอะตอมคล้ายคลึงกัน แทนที่ไอโซลิวซีนในศูนย์กลางการสังเคราะห์อย่างง่ายดาย และสังเคราะห์จับกับ tRNA ในกรณีนี้คือศูนย์ราชทัณฑ์ของการสังเคราะห์เข้ามาเล่น หากกิ๊บตัวรับการยืดผมถูกชาร์จอย่างถูกต้องและมีไอโซลิวซีน กิ๊บจะไม่สามารถบีบเข้าไปในศูนย์แก้ไขได้: มันเล็กเกินไปสำหรับโมเลกุลนี้ ดังนั้น tRNA ที่ยืดออกจึงไม่ถูกยึดโดยสิ่งใดอีกต่อไป และมันถูกแยกออกจากการสังเคราะห์ แต่ถ้าวาลีนติดอยู่กับ tRNA มันจะหลุดเข้าไปในศูนย์แก้ไข ดังนั้นจึงทำให้ tRNA เชื่อมต่อกับมันในการสังเคราะห์ การอยู่ภายใน tRNA ที่ยาวเกินไปเช่นนี้เป็นสัญญาณผิดพลาดสำหรับซินธิเทส ซึ่งเปลี่ยนการกำหนดค่าเชิงพื้นที่ของมัน ผลที่ตามมา:
=) วาลีนแยกออกจาก tRNA และนำออกจากซินธิเทส
=) กิ๊บของตัวรับกลับไปที่ไซต์สังเคราะห์เพื่อรอการเกาะติดกับกรดอะมิโน
=) ซินธิเทสจับกับกรดอะมิโนใหม่ "ชาร์จ" tRNA กับมัน แล้วตรวจสอบอีกครั้งว่าไอโซลิวซีนถูกใช้สำหรับสิ่งนี้หรือไม่
ซินธิเตสอื่นใช้กลไกการจดจำคู่ที่คล้ายกัน
คุณสมบัติทางกายภาพและเคมีของ DNA
ปัจจัยต่างๆ ที่ทำลายพันธะไฮโดรเจน (อุณหภูมิเพิ่มขึ้นสูงกว่า 80 C, การเปลี่ยนแปลงของ pH และความแข็งแรงของไอออนิก, การกระทำของยูเรีย ฯลฯ) ทำให้เกิดการเสื่อมสภาพของดีเอ็นเอ กล่าวคือ การเปลี่ยนแปลงการจัดเรียงเชิงพื้นที่ของสาย DNA โดยไม่ทำลายพันธะโควาเลนต์ เกลียวคู่ของ DNA ในระหว่างการแปลงสภาพถูกแบ่งออกเป็นสายโซ่ส่วนประกอบทั้งหมดหรือบางส่วน การเปลี่ยนสภาพของ DNA นั้นมาพร้อมกับการเพิ่มขึ้นของการดูดกลืนแสงในบริเวณ UV ของฐาน purine และ pyrimidine ปรากฏการณ์นี้เรียกว่า เอฟเฟกต์ไฮเปอร์โครมิก . การเปลี่ยนสภาพยังช่วยลดความหนืดสูงที่มีอยู่ในสารละลาย DNA ดั้งเดิม เมื่อโครงสร้าง DNA ที่มีสายสองเส้นเดิมกลับคืนมา อันเป็นผลมาจากการเปลี่ยนสภาพใหม่ การดูดซับที่ 260 นาโนเมตรโดยเบสไนโตรเจนจะลดลงเนื่องจาก "การป้องกัน" ของพวกมัน ปรากฏการณ์นี้เรียกว่า เอฟเฟกต์ไฮโปโครมิก .
การ "คลี่คลาย" ของ DNA แต่ละตัวในสายโซ่ส่วนประกอบนั้นดำเนินการภายในช่วงอุณหภูมิที่กำหนด จุดกึ่งกลางของช่วงเวลานี้เรียกว่าจุดหลอมเหลว อุณหภูมิหลอมเหลวของ DNA ขึ้นอยู่กับสภาวะมาตรฐาน (ค่า pH และความแรงของอิออนิกที่แน่นอน) กับอัตราส่วนของเบสไนโตรเจน คู่ G-C ที่มีพันธะไฮโดรเจนสามพันธะนั้นแข็งแกร่งกว่า ดังนั้น ยิ่งเนื้อหาของคู่ G-C ใน DNA สูงขึ้น จุดหลอมเหลวก็จะยิ่งสูงขึ้น
หน้าที่ของ DNA. ในลำดับของนิวคลีโอไทด์ในโมเลกุลดีเอ็นเอ ข้อมูลทางพันธุกรรมจะถูกเข้ารหัส หน้าที่หลักของ DNA ประการแรกคือเพื่อให้แน่ใจว่ามีการสืบพันธุ์ของตัวเองในรุ่นเซลล์และรุ่นของสิ่งมีชีวิตและประการที่สองเพื่อให้แน่ใจว่าการสังเคราะห์โปรตีน หน้าที่เหล่านี้เกิดจากการที่โมเลกุลดีเอ็นเอทำหน้าที่เป็นเมทริกซ์ในกรณีแรกสำหรับการจำลองแบบ กล่าวคือ คัดลอกข้อมูลในโมเลกุล DNA ของลูกสาวในวินาที - เพื่อการถอดความเช่น เพื่อเข้ารหัสข้อมูลลงในโครงสร้างอาร์เอ็นเอ
ข้าว. 5 เส้นหลอมเหลว (DNA denaturation)
สายเสริมของ DNA ที่แยกจากกันระหว่างการเปลี่ยนสภาพสามารถเชื่อมต่อใหม่เป็นเกลียวคู่ได้ภายใต้เงื่อนไขบางประการ กระบวนการนี้เรียกว่า RENATURATION หากการเสียสภาพธรรมชาติไม่เกิดขึ้นอย่างสมบูรณ์และอย่างน้อยก็ยังมีเบสสองสามตัวที่ไม่สูญเสียปฏิกิริยาจากพันธะไฮโดรเจน การเปลี่ยนสภาพจะดำเนินไปอย่างรวดเร็ว
ไซโตพลาสซึมของเซลล์ประกอบด้วย RNA หน้าที่หลักสามประเภท เหล่านี้คือ RNA ของผู้ส่งสาร - mRNA ที่ทำหน้าที่เป็นแม่แบบสำหรับการสังเคราะห์โปรตีน ไรโบโซม RNAs - rRNAs ที่ทำหน้าที่เป็นส่วนประกอบโครงสร้างของไรโบโซม และถ่ายโอน RNAs - tRNA ที่เกี่ยวข้องกับการแปล (การแปล) ของข้อมูล mRNA เป็นลำดับกรดอะมิโนในโปรตีน
ตารางที่ 2 แสดงความแตกต่างระหว่าง DNA และ RNA ในแง่ของโครงสร้าง การแปลเป็นภาษาท้องถิ่นในเซลล์ และหน้าที่
ตารางที่ 2 ความแตกต่างระหว่าง DNA และ RNA
โอน RNA, tRNA-กรดไรโบนิวคลีอิก ซึ่งทำหน้าที่ขนส่ง AA ไปยังบริเวณที่สังเคราะห์โปรตีน มีความยาวโดยทั่วไป 73 ถึง 93 นิวคลีโอไทด์และมีขนาดประมาณ 5 นาโนเมตร นอกจากนี้ tRNA ยังเกี่ยวข้องโดยตรงกับการเติบโตของสายโซ่พอลิเปปไทด์ โดยเชื่อมต่อ - อยู่ในคอมเพล็กซ์ที่มีกรดอะมิโน - กับโคดอน mRNA และให้โครงสร้างของคอมเพล็กซ์ที่จำเป็นสำหรับการก่อตัวของพันธะเปปไทด์ใหม่ กรดอะมิโนแต่ละชนิดมี tRNA ของตัวเอง tRNA เป็น RNA สายเดี่ยว แต่ในรูปแบบการทำงานจะมีรูปแบบ "โคลเวอร์ลีฟ" AA ยึดติดกับปลายโมเลกุล 3 "อย่างโควาเลนต์โดยใช้เอ็นไซม์ aminoacyl-tRNA synthetase ซึ่งจำเพาะสำหรับ tRNA แต่ละประเภท ที่ไซต์ C มีแอนติโคดอนที่สอดคล้องกับ AA-te tRNA ถูกสังเคราะห์โดย RNA polymerase ธรรมดาในกรณีนี้ ของโปรคาริโอตและโดย RNA polymerase III ในกรณีของยูคาริโอต การถอดเสียงของยีน tRNA ได้รับการประมวลผลแบบหลายขั้นตอน ซึ่งนำไปสู่การก่อตัวของโครงสร้างเชิงพื้นที่ตามแบบฉบับของ tRNA
การประมวลผล tRNA ประกอบด้วยขั้นตอนสำคัญ 5 ขั้นตอน:
การกำจัดลำดับนิวคลีโอไทด์ผู้นำ 5";
การลบลำดับเทอร์มินัล 3';
เพิ่มลำดับ CCA ที่ส่วนท้าย 3";
การตัดตอนของ introns (ในยูคาริโอตและอาร์เคีย);
การดัดแปลงของนิวคลีโอไทด์แต่ละตัว
การขนส่ง tRNA ดำเนินการตามเส้นทางที่ขึ้นกับ Ran โดยมีส่วนร่วมของปัจจัยการขนส่งที่ส่งออกใน เสื้อ ซึ่งรับรู้ถึงลักษณะเฉพาะของ str-ru ทุติยภูมิและตติยภูมิของ tRNA ที่โตเต็มที่: ส่วนที่เป็นเกลียวคู่สั้นและประมวลผลอย่างถูกต้อง 5 "- และ 3" สิ้นสุด กลไกนี้ช่วยให้แน่ใจว่ามีเพียง tRNA ที่โตเต็มที่เท่านั้นที่ส่งออกจากนิวเคลียส
62. การแปล - การจดจำ codon mRNA
การแปลคือการสังเคราะห์โปรตีนที่ดำเนินการโดยไรโบโซมจากกรดอะมิโนบนเทมเพลต mRNA (หรือ RNA) องค์ประกอบที่เป็นส่วนประกอบของกระบวนการแปล: กรดอะมิโน tRNA ไรโบโซม mRNA เอนไซม์สำหรับ aminoacylation ของ tRNA ปัจจัยการแปลโปรตีน (ปัจจัยโปรตีนของการเริ่มต้น การยืดตัว การสิ้นสุด - โปรตีน extraribosomal จำเพาะที่จำเป็นสำหรับกระบวนการแปล), ATP และ GTP แหล่งพลังงาน , แมกนีเซียมไอออน (ทำให้โครงสร้างไรโบโซมเสถียร) กรดอะมิโน 20 ชนิดมีส่วนร่วมในการสังเคราะห์โปรตีน เพื่อให้กรดอะมิโน "รับรู้" ตำแหน่งของมันในสายโซ่พอลิเปปไทด์ในอนาคต กรดอะมิโนจะต้องจับกับ RNA การถ่ายโอน (tRNA) ที่ทำหน้าที่อะแดปเตอร์ tRNA ที่จับกับกรดอะมิโนจะรับรู้ codon ที่สอดคล้องกันบน mRNA mRNA codon การรับรู้:
การทำงานร่วมกันของ codon-anticodon นั้นขึ้นอยู่กับหลักการของการเติมเต็มและการต่อต้านขนาน:
3'----C - G-A*------5' tRNA แอนติโคดอน
5'-----G-C-Y*------3' mRNA codon
สมมติฐานการวอกแวกถูกเสนอโดย F. Crick:
3'-เบสของโคดอน mRNA มีการจับคู่ที่ไม่เข้มงวดกับ 5'-เบสของแอนติโคดอน tRNA: ตัวอย่างเช่น Y (mRNA) สามารถโต้ตอบกับ A และ G (tRNA)
tRNA บางตัวสามารถจับคู่กับโคดอนได้มากกว่าหนึ่งโคดอน
63. ลักษณะขององค์ประกอบที่เป็นส่วนประกอบของกระบวนการแปลการแปล (translatio-translation) เป็นกระบวนการสังเคราะห์โปรตีนจากกรดอะมิโนบนเมทริกซ์ของข้อมูล (เมทริกซ์) RNA (mRNA, mRNA) ที่ดำเนินการโดยไรโบโซม
การสังเคราะห์โปรตีนเป็นพื้นฐานของชีวิตเซลล์ เพื่อดำเนินการตามกระบวนการนี้ในเซลล์ของสิ่งมีชีวิตทั้งหมดมีออร์แกเนลล์พิเศษ - ไรโบโซม- ไรโบนิวคลีโอโปรตีนเชิงซ้อนที่สร้างขึ้นจาก 2 หน่วยย่อย: ใหญ่และเล็ก. หน้าที่ของไรโบโซมคือการจดจำตัวอักษรสามตัว (สามนิวคลีโอไทด์) codons mRNA เปรียบเทียบกับ tRNA anticodons ที่สอดคล้องกัน กรดอะมิโนและการเพิ่มกรดอะมิโนเหล่านี้ไปยังสายโปรตีนที่กำลังเติบโต เมื่อเคลื่อนที่ไปตามโมเลกุล mRNA ไรโบโซมจะสังเคราะห์โปรตีนตามข้อมูลที่มีอยู่ในโมเลกุล mRNA
สำหรับการรับรู้ AK-t ในเซลล์มี "อะแดปเตอร์" พิเศษ ถ่ายโอนโมเลกุลอาร์เอ็นเอ(tRNA). โมเลกุลรูปใบโคลเวอร์ลีฟเหล่านี้มีไซต์ (แอนติโคดอน) ประกอบกับโคดอน mRNA เช่นเดียวกับไซต์อื่นที่มีกรดอะมิโนที่สอดคล้องกับโคดอนนั้นติดอยู่ การเกาะติดของกรดอะมิโนกับ tRNA เกิดขึ้นในปฏิกิริยาที่ขึ้นกับพลังงานโดยเอ็นไซม์ aminoacyl-tRNA synthetases และโมเลกุลที่ได้จะเรียกว่า aminoacyl-tRNA ดังนั้น ความจำเพาะของการแปลจะถูกกำหนดโดยการทำงานร่วมกันระหว่าง mRNA codon และ tRNA anticodon เช่นเดียวกับความจำเพาะของการสังเคราะห์ aminoacyl-tRNA ที่ยึดกรดอะมิโนเข้ากับ tRNA ที่สอดคล้องกันอย่างเคร่งครัด (ตัวอย่างเช่น GGU codon จะสอดคล้องกับ tRNA ที่มี CCA anticodon และ AK glycine เท่านั้น)
โปรคาริโอต ไรโบโซม
5S และ 23S rRNA 16S rRNA
กระรอก 34 ตัว กระรอก 21 ตัว
โปรคาริโอตไรโบโซมมีค่าคงที่การตกตะกอนที่ 70S ซึ่งเป็นสาเหตุที่เรียกว่าอนุภาค 70S สร้างขึ้นจากหน่วยย่อยที่แตกต่างกันสองหน่วย: หน่วยย่อย 30S และ 50S แต่ละหน่วยย่อยเป็นคอมเพล็กซ์ของโปรตีน rRNA และไรโบโซม
อนุภาค 30S ประกอบด้วยโมเลกุล 16S rRNA หนึ่งโมเลกุล และในกรณีส่วนใหญ่ โมเลกุลโปรตีนหนึ่งโมเลกุลจากกว่า 20 สปีชีส์ (21) หน่วยย่อย 50S ประกอบด้วยโมเลกุล rRNA สองโมเลกุล (23S และ 5S) ประกอบด้วยโปรตีนที่แตกต่างกันมากกว่า 30 ชนิด (34) ซึ่งเป็นตัวแทนของสำเนาหนึ่งชุด โปรตีนไรโบโซมส่วนใหญ่ทำหน้าที่เกี่ยวกับโครงสร้าง
ยูคาริโอตไรโบโซม
5ส; 5,8S และ 28S rRNA 18S rRNA
อย่างน้อย 50 โปรตีน อย่างน้อย 33 โปรตีน
ไรโบโซมประกอบด้วยหน่วยย่อยขนาดใหญ่และขนาดเล็ก พื้นฐานของโครงสร้างของแต่ละหน่วยย่อยคือ rRNA ที่พับซ้อนอย่างซับซ้อน โปรตีนไรโบโซมติดอยู่กับโครงนั่งร้าน rRNA
ค่าสัมประสิทธิ์การตกตะกอนของไรโบโซมยูคาริโอตที่สมบูรณ์คือประมาณ 80 หน่วย Svedberg (80S) และค่าสัมประสิทธิ์การตกตะกอนของอนุภาคย่อยคือ 40S และ 60S
หน่วยย่อย 40S ที่เล็กกว่าประกอบด้วยโมเลกุล 18S rRNA หนึ่งโมเลกุลและโมเลกุลโปรตีน 30-40 ตัว หน่วยย่อย 60S ขนาดใหญ่ประกอบด้วย rRNA สามประเภทโดยมีค่าสัมประสิทธิ์การตกตะกอนของโปรตีน 5S, 5.8S และ 28S และ 40-50 (ตัวอย่างเช่น ไรโบโซมตับของหนูแรทมีโปรตีน 49 ตัว)
บริเวณหน้าที่ของไรโบโซม
P - ไซต์เปปทิดิลสำหรับเปปทิดิล tRNA
ไซต์ A - aminoacyl สำหรับ aminoacyl tRNA
E - ไซต์สำหรับการปล่อย tRNA จากไรโบโซม
ไรโบโซมประกอบด้วยตำแหน่งทำงาน 2 ตำแหน่งสำหรับการโต้ตอบกับ tRNA: อะมิโนอะซิล (ตัวรับ) และเปปทิดิล (ผู้ให้) Aminoacyl-tRNA เข้าสู่ไซต์ตัวรับของไรโบโซมและโต้ตอบเพื่อสร้างพันธะไฮโดรเจนระหว่าง codon และ anticodon triplets หลังจากการก่อตัวของพันธะไฮโดรเจน ระบบก้าวหน้า 1 codon และไปสิ้นสุดที่ไซต์ผู้บริจาค ในเวลาเดียวกัน codon ใหม่จะปรากฏในไซต์ตัวรับที่ว่างและแนบ aminoacyl-t-RNA ที่สอดคล้องกัน
ไรโบโซม: โครงสร้าง, หน้าที่
ไรโบโซมเป็นศูนย์กลางของไซโตพลาสซึมของการสังเคราะห์โปรตีน ประกอบด้วยหน่วยย่อยขนาดใหญ่และขนาดเล็กซึ่งแตกต่างกันในค่าสัมประสิทธิ์การตกตะกอน (อัตราการตกตะกอนในระหว่างการหมุนเหวี่ยง) แสดงในหน่วยของ Svedberg - S.
ไรโบโซมมีอยู่ทั้งในเซลล์ยูคาริโอตและเซลล์โปรคาริโอต เนื่องจากพวกมันทำหน้าที่สำคัญใน การสังเคราะห์โปรตีนแต่ละเซลล์ประกอบด้วยออร์แกเนลล์ขนาดเล็กที่โค้งมนนับหมื่น (มากถึงหลายล้าน) เป็นอนุภาคไรโบนิวคลีโอโปรตีนกลม เส้นผ่านศูนย์กลาง 20-30 นาโนเมตร ไรโบโซมประกอบด้วยหน่วยย่อยขนาดใหญ่และขนาดเล็ก ซึ่งแตกต่างกันในค่าสัมประสิทธิ์การตกตะกอน (อัตราการตกตะกอนในระหว่างการหมุนเหวี่ยง) แสดงในหน่วย Svedberg - S หน่วยย่อยเหล่านี้รวมกันเมื่อมีสายของ m-RNA (เมทริกซ์หรือข้อมูล RNA) คอมเพล็กซ์ของกลุ่มไรโบโซมที่รวมตัวกันโดยโมเลกุล mRNA เดียวเช่นลูกปัดเรียกว่า polysome. โครงสร้างเหล่านี้อาจอยู่ในไซโตพลาสซึมอย่างอิสระหรือติดอยู่กับเยื่อหุ้มของ ER แบบเม็ด (ในทั้งสองกรณี การสังเคราะห์โปรตีนจะดำเนินการอย่างแข็งขัน)
โพลีโซมของ ER แบบเม็ดจะก่อตัวเป็นโปรตีนที่ถูกขับออกจากเซลล์และใช้สำหรับความต้องการของสิ่งมีชีวิตทั้งหมด (เช่น เอนไซม์ย่อยอาหาร โปรตีนจากน้ำนมแม่) นอกจากนี้ ไรโบโซมยังมีอยู่บนผิวด้านในของเยื่อหุ้มไมโตคอนเดรีย ซึ่งพวกมันยังมีส่วนร่วมในการสังเคราะห์โมเลกุลโปรตีนอีกด้วย
Transfer RNA (tRNA) มีบทบาทสำคัญในกระบวนการใช้ข้อมูลทางพันธุกรรมโดยเซลล์ การส่งกรดอะมิโนที่จำเป็นไปยังสถานที่ประกอบของสายเปปไทด์ tRNA ทำหน้าที่เป็นตัวกลางในการแปล
โมเลกุล tRNA เป็นสายโซ่โพลีนิวคลีโอไทด์ที่สังเคราะห์ในลำดับดีเอ็นเอจำเพาะ ประกอบด้วยนิวคลีโอไทด์จำนวนค่อนข้างน้อย -75-95 อันเป็นผลมาจากการเชื่อมต่อเสริมของฐานที่ตั้งอยู่ในส่วนต่าง ๆ ของสายโซ่โพลีนิวคลีโอไทด์ tRNA มันได้โครงสร้างที่มีรูปร่างคล้ายใบโคลเวอร์ (รูปที่ 3.26)
ข้าว. 3.26. โครงสร้างของโมเลกุล tRNA ทั่วไป
มีสี่ส่วนหลักที่ทำหน้าที่ต่างกัน ตัวรับ"ก้าน" ประกอบขึ้นจากส่วนปลายเชื่อมต่อเสริมสองส่วนของ tRNA ประกอบด้วยเจ็ดคู่ฐาน ส่วนปลาย 3' ของก้านนี้ค่อนข้างยาวกว่าและก่อตัวเป็นบริเวณเกลียวเดี่ยวที่ลงท้ายด้วยลำดับ CCA ที่มีหมู่ OH อิสระ กรดอะมิโนที่ขนส่งได้ติดอยู่ที่ส่วนท้ายนี้ อีกสามกิ่งที่เหลือเป็นลำดับนิวคลีโอไทด์ที่จับคู่เสริมที่สิ้นสุดในส่วนที่ไม่มีการจับคู่ซึ่งก่อตัวเป็นลูป แกนกลางของกิ่งเหล่านี้ - แอนติโคดอน - ประกอบด้วยนิวคลีโอไทด์ห้าคู่และมีแอนติโคดอนอยู่ตรงกลางของวง แอนติโคดอนคือนิวคลีโอไทด์สามชนิดที่ประกอบกันกับโคดอน mRNA ซึ่งเข้ารหัสกรดอะมิโนที่ขนส่งโดย tRNA นี้ไปยังบริเวณที่มีการสังเคราะห์เปปไทด์
ระหว่างตัวรับและกิ่งแอนติโคดอนมีกิ่งสองข้าง ในลูปของมัน พวกมันมีเบสดัดแปลง - ไดไฮโดรริดีน (D-loop) และแฝดสามของ TψC โดยที่ \y คือ pseudouriain (T^C-loop)
ระหว่างกิ่ง aiticodone และ T^C มีลูปเพิ่มเติมซึ่งรวมถึงนิวคลีโอไทด์ตั้งแต่ 3-5 ถึง 13-21
โดยทั่วไป tRNA ประเภทต่างๆ มีลักษณะเฉพาะโดยความคงตัวบางอย่างของลำดับนิวคลีโอไทด์ ซึ่งส่วนใหญ่มักประกอบด้วย 76 นิวคลีโอไทด์ ความแปรปรวนของจำนวนส่วนใหญ่เกิดจากการเปลี่ยนแปลงจำนวนนิวคลีโอไทด์ในวงเพิ่มเติม บริเวณเสริมที่รองรับโครงสร้าง tRNA มักจะได้รับการอนุรักษ์ไว้ โครงสร้างหลักของ tRNA ซึ่งกำหนดโดยลำดับของนิวคลีโอไทด์สร้างโครงสร้างรองของ tRNA ซึ่งมีรูปร่างเหมือนใบโคลเวอร์ ในทางกลับกัน โครงสร้างทุติยภูมิทำให้เกิดโครงสร้างตติยภูมิสามมิติ ซึ่งมีลักษณะเป็นเกลียวคู่ตั้งฉากสองอัน (รูปที่ 3.27) หนึ่งในนั้นเกิดจากตัวรับและกิ่ง TψC อีกอันเกิดจากกิ่ง anticodon และ D
ที่ปลายเกลียวคู่อันใดอันหนึ่งคือกรดอะมิโนที่ขนส่ง อีกอันหนึ่งคือแอนติโคดอน พื้นที่เหล่านี้อยู่ห่างไกลจากกันมากที่สุด ความเสถียรของโครงสร้างตติยภูมิของ tRNA นั้นยังคงอยู่เนื่องจากการปรากฏตัวของพันธะไฮโดรเจนเพิ่มเติมระหว่างฐานของสายโซ่พอลินิวคลีโอไทด์ ซึ่งตั้งอยู่ในส่วนต่างๆ ของมัน แต่ในโครงสร้างตติยภูมิใกล้เคียงกัน
tRNA ประเภทต่างๆ มีโครงสร้างระดับอุดมศึกษาที่คล้ายคลึงกัน แม้ว่าจะมีรูปแบบที่แตกต่างกันบ้าง
ข้าว. 3.27. การจัดระเบียบเชิงพื้นที่ของ tRNA:
I - โครงสร้างรองของ tRNA ในรูปแบบของ "ใบโคลเวอร์" กำหนดโดยโครงสร้างหลัก (ลำดับของนิวคลีโอไทด์ในสายโซ่);
II - การฉายภาพสองมิติของโครงสร้างระดับอุดมศึกษาของ tRNA;
III - เลย์เอาต์ของโมเลกุล tRNA ในอวกาศ
ภาคผนวก (ในกรณีที่มีคนไม่เข้าใจสิ่งนี้)
ฟันฟ้าผ่า - นิวคลีโอไทด์ (Adenine-Tymine / Uracil /, Guanine-Cytazine) สายฟ้าทั้งหมดเป็นดีเอ็นเอ

ในการถ่ายโอนข้อมูลจาก DNA คุณต้องทำลาย 2 เส้น พันธะระหว่าง A-T และ G-C คือไฮโดรเจน เอนไซม์เฮลิเคสแตกง่าย:

เพื่อป้องกันไม่ให้เกิดปม (ตัวอย่างเช่นฉันบิดผ้าเช็ดตัว):



Topoisomerase ตัด DNA หนึ่งเส้นที่ต้นกำเนิดของการจำลองแบบเพื่อไม่ให้สายโซ่บิด

เมื่อด้ายหนึ่งว่าง ด้ายที่สองสามารถหมุนรอบแกนได้อย่างง่ายดาย จึงช่วยลดความตึงเครียดในระหว่างการ "คลี่คลาย" โหนดไม่ปรากฏขึ้น ประหยัดพลังงาน
จากนั้นจึงจำเป็นต้องใช้ไพรเมอร์ RNA เพื่อเริ่มรวบรวม RNA โปรตีนที่ประกอบ mRNA ไม่สามารถประกอบแค่นิวคลีโอไทด์แรกได้ มันต้องการชิ้นส่วนของ RNA เพื่อเริ่มต้น (มีการเขียนรายละเอียดไว้ที่นั่น ฉันจะเขียนในภายหลัง) ชิ้นนี้เรียกว่าไพรเมอร์อาร์เอ็นเอ และโปรตีนนี้จับนิวคลีโอไทด์ตัวแรกไว้อยู่แล้ว


เมื่ออธิบายโครงสร้างของกรดนิวคลีอิก ระดับต่างๆ ของการจัดเรียงตัวของโมเลกุลขนาดใหญ่จะถูกนำมาพิจารณาด้วย: โครงสร้างปฐมภูมิและทุติยภูมิ
โครงสร้างหลักของกรดนิวคลีอิกคือองค์ประกอบของนิวคลีโอไทด์และลำดับที่แน่นอนของหน่วยนิวคลีโอไทด์ในสายโซ่โพลีเมอร์
โครงสร้างรองของอาร์เอ็นเอ โมเลกุลของกรดไรโบนิวคลีอิกถูกสร้างขึ้นจากสายพอลินิวคลีโอไทด์เดี่ยว
โครงสร้างรองของ RNA
ส่วนที่แยกจากกันของสายโซ่ RNA จะสร้างวงวนเป็นวง - "กิ๊บติดผม" เนื่องจากพันธะไฮโดรเจนระหว่างฐานไนโตรเจนเสริม A-U และ GC ส่วนของสายโซ่ RNA ในโครงสร้างที่เป็นเกลียวนั้นมีลักษณะตรงข้ามขนานกันแต่ไม่ได้เป็นส่วนเสริมอย่างสมบูรณ์เสมอไป พวกมันประกอบด้วยนิวคลีโอไทด์เรซิดิวที่ไม่ถูกจับคู่หรือแม้แต่ลูปที่เป็นเกลียวเดียวที่ไม่เข้ากับเกลียวคู่ การปรากฏตัวของบริเวณที่เป็นเกลียวเป็นลักษณะของ RNA ทุกประเภท
บทบาทหลักของ RNA คือการมีส่วนร่วมโดยตรงในการสังเคราะห์โปรตีน
เป็นที่ทราบกันดีว่า RNA ของเซลล์สามประเภทซึ่งแตกต่างกันในตำแหน่งในเซลล์ องค์ประกอบ ขนาด และคุณสมบัติที่กำหนดบทบาทเฉพาะของพวกมันในการก่อตัวของโมเลกุลโปรตีน:
- RNA ข้อมูล (เมทริกซ์) ส่งข้อมูลที่เข้ารหัสใน DNA เกี่ยวกับโครงสร้างของโปรตีนจากนิวเคลียสของเซลล์ไปยังไรโบโซมซึ่งดำเนินการสังเคราะห์โปรตีน โครงสร้างหลักของ mRNA ทั้งหมด โดยไม่คำนึงถึงเอกลักษณ์ของลำดับการเข้ารหัส มีโครงสร้างเดียวกันของปลาย 5'- และ 3'-
ดังนั้น ที่ปลาย 5'- จึงมีนิวคลีโอไทด์ 7-เมทิลกัวโนซีน-5'-ไตรฟอสเฟตที่ดัดแปลงแล้ว (หมวก) นิวคลีโอไทด์หลายสิบตัวแยกแคปออกจากโคดอนเริ่มต้น ปกติคือ -AUG- triplet ขอบเขตการเข้ารหัสตามด้วยหนึ่งใน codon การสิ้นสุด -UGA-, -UUA-, -UAG- ที่ส่วนท้าย 3' ของ mRNA ส่วนใหญ่ มีลำดับนิวคลีโอไทด์ของอะดีโนซีนโมโนฟอสเฟต 100-200 เรซิดิว
- ถ่ายโอน RNAs รวบรวมกรดอะมิโนในไซโตพลาสซึมของเซลล์และถ่ายโอนไปยังไรโบโซม โมเลกุล RNA ประเภทนี้ "เรียนรู้" จากส่วนที่เกี่ยวข้องของสายโซ่ RNA ของผู้ส่งสาร ซึ่งกรดอะมิโนควรมีส่วนร่วมในการสังเคราะห์โปรตีน
โครงสร้างเชิงพื้นที่ของ tRNA ใดๆ โดยไม่คำนึงถึงความแตกต่างในลำดับนิวคลีโอไทด์ อธิบายโดยแบบจำลองโคลเวอร์ลีฟสากล โมเลกุล tRNA แต่ละตัวมีส่วนลูกโซ่ที่ไม่เกี่ยวข้องกับการก่อตัวของพันธะไฮโดรเจนระหว่างสารตกค้างของนิวคลีโอไทด์
ซึ่งรวมถึง โดยเฉพาะอย่างยิ่ง ตำแหน่งที่รับผิดชอบในการจับกับกรดอะมิโนที่ปลาย 3' ของโมเลกุลและแอนติโคดอน ซึ่งเป็นทริปเปิ้ลที่จำเพาะของนิวคลีโอไทด์ที่มีปฏิสัมพันธ์กับโคดอน mRNA
- ไรโบโซม RNA ให้การสังเคราะห์โปรตีนของโครงสร้างบางอย่าง อ่านข้อมูลจากข้อมูล (เมทริกซ์) RNA rRNAs สร้างสารเชิงซ้อนที่มีโปรตีนที่เรียกว่าไรโบโซม
ไรโบโซมแต่ละตัวประกอบด้วยสองหน่วยย่อย - เล็ก (40S) และใหญ่ (60S) หน่วยย่อยของไรโบโซมแตกต่างกันไม่เพียงแต่ในชุดของ rRNA แต่ยังรวมถึงจำนวนและโครงสร้างของโปรตีนด้วย
วันที่ตีพิมพ์: 2015-02-03; อ่าน: 2729 | เพจละเมิดลิขสิทธิ์

RNA เป็นพอลิเมอร์ที่มีโมโนเมอร์เป็นไรโบนิวคลีโอไทด์
ต่างจาก DNA ตรงที่ RNA ไม่ได้เกิดขึ้นจากสองสาย แต่เกิดจากสายโซ่โพลีนิวคลีโอไทด์หนึ่งสาย (ยกเว้น ไวรัสที่ประกอบด้วย RNA บางตัวมี RNA แบบสองสาย) RNA nucleotides สามารถสร้างพันธะไฮโดรเจนซึ่งกันและกันได้ RNA chain นั้นสั้นกว่า DNA chain มาก
อาร์เอ็นเอโมโนเมอร์ - นิวคลีโอไทด์ (ไรโบนิวคลีโอไทด์) - ประกอบด้วยสารตกค้างสาม: 1) ฐานไนโตรเจน 2) โมโนแซ็กคาไรด์ห้าคาร์บอน (เพนโทส) และ 3) กรดฟอสฟอริก ฐานไนโตรเจนของอาร์เอ็นเอยังอยู่ในคลาสของไพริมิดีนและพิวรีน
เบสไพริมิดีนของอาร์เอ็นเอ ได้แก่ ยูราซิล ไซโตซีน เบสพิวรีน - อะดีนีนและกวานีน
31. ประเภทของ RNA และคุณสมบัติของโครงสร้าง
โมโนแซ็กคาไรด์ของ RNA นิวคลีโอไทด์ถูกแทนด้วยไรโบส
RNA มีสามประเภท: 1) ข้อมูล (เมทริกซ์) RNA - mRNA (mRNA), 2) ถ่ายโอน RNA - tRNA, 3) ไรโบโซม RNA - rRNA
RNA ทุกประเภทเป็นโพลีนิวคลีโอไทด์ที่ไม่มีการแบ่งแยก มีโครงสร้างเชิงพื้นที่จำเพาะ และมีส่วนร่วมในกระบวนการสังเคราะห์โปรตีน
ข้อมูลเกี่ยวกับโครงสร้างของ RNA ทุกประเภทถูกเก็บไว้ใน DNA กระบวนการสังเคราะห์อาร์เอ็นเอบนเทมเพลตดีเอ็นเอมักเรียกว่าการถอดความ

RNA การถ่ายโอนมักจะมีนิวคลีโอไทด์ 76 (จาก 75 ถึง 95) น้ำหนักโมเลกุล - 25,000–30,000
tRNA คิดเป็นประมาณ 10% ของเนื้อหา RNA ทั้งหมดในเซลล์ หน้าที่ของ tRNA: 1) การขนส่งกรดอะมิโนไปยังบริเวณที่มีการสังเคราะห์โปรตีน ไปยังไรโบโซม 2) ผู้ไกล่เกลี่ยการแปล พบ tRNA ประมาณ 40 ชนิดในเซลล์ แต่ละชนิดมีลักษณะเฉพาะของลำดับนิวคลีโอไทด์สำหรับมันเท่านั้น ในเวลาเดียวกัน tRNA ทั้งหมดมีบริเวณเสริมภายในโมเลกุลหลายส่วน เนื่องจาก tRNAs ได้รับโครงสร้างที่คล้ายกับใบโคลเวอร์
tRNA ใดๆ มีลูปสำหรับการสัมผัสกับไรโบโซม (1), ลูปแอนติโคดอน (2), ลูปสำหรับการสัมผัสกับเอ็นไซม์ (3), ก้านตัวรับ (4) และแอนติโคดอน (5) กรดอะมิโนติดอยู่ที่ปลาย 3' ของก้านตัวรับ แอนติโคดอน - นิวคลีโอไทด์สามตัวที่ "รับรู้" โคดอน mRNA
ควรเน้นว่า tRNA เฉพาะสามารถขนส่งกรดอะมิโนที่กำหนดไว้อย่างเคร่งครัดซึ่งสอดคล้องกับแอนติโคดอนของมัน ความจำเพาะของการเชื่อมต่อของกรดอะมิโนและ tRNA ทำได้เนื่องจากคุณสมบัติของเอนไซม์ aminoacyl-tRNA synthetase
Ribosomal RNAs มีนิวคลีโอไทด์ 3000–5000; น้ำหนักโมเลกุล - 1,000,000–1.500,000
rRNA คิดเป็น 80–85% ของเนื้อหา RNA ทั้งหมดในเซลล์ ในความซับซ้อนที่มีโปรตีนไรโบโซม rRNA จะสร้างไรโบโซม - ออร์แกเนลล์ที่ทำการสังเคราะห์โปรตีน ในเซลล์ยูคาริโอต การสังเคราะห์ rRNA เกิดขึ้นในนิวเคลียส หน้าที่ของ rRNA: 1) องค์ประกอบโครงสร้างที่จำเป็นของไรโบโซม และทำให้แน่ใจในการทำงานของไรโบโซม; 2) สร้างความมั่นใจปฏิสัมพันธ์ของไรโบโซมและ tRNA; 3) การจับเริ่มต้นของไรโบโซมและโคดอนตัวเริ่มต้น mRNA และการกำหนดกรอบการอ่าน 4) การก่อตัวของจุดศูนย์กลางแอคทีฟของไรโบโซม
Messenger RNAs มีความหลากหลายในเนื้อหานิวคลีโอไทด์และน้ำหนักโมเลกุล (จาก 50,000 ถึง 4,000,000)
ส่วนแบ่งของ mRNA คิดเป็น 5% ของเนื้อหา RNA ทั้งหมดในเซลล์ หน้าที่ของ mRNA: 1) การถ่ายโอนข้อมูลทางพันธุกรรมจาก DNA ไปยังไรโบโซม 2) เมทริกซ์สำหรับการสังเคราะห์โมเลกุลโปรตีน 3) การกำหนดลำดับกรดอะมิโนของโครงสร้างหลักของโมเลกุลโปรตีน
อ่านยัง
RNA เป็นพอลิเมอร์ที่มีโมโนเมอร์เป็นนิวคลีโอไทด์
เบสไนโตรเจนสามชนิดนั้นเหมือนกับใน DNA (อะดีนีน, กัวนีน, ไซโตซีน); ที่สี่ - uracil - มีอยู่ในโมเลกุล RNA แทนที่จะเป็นไทมีน RNA nucleotides มีไรโบสแทนที่จะเป็นดีออกซีไรโบส ในสายโซ่ RNA...
RNA สามประเภทหลัก: ข้อมูล(mRNA) หรือ เมทริกซ์(เอ็มอาร์เอ็นเอ) ไรโบโซมอล(rRNA) และ ขนส่ง(tRNA). พวกเขาแตกต่างกันในขนาดโมเลกุลและหน้าที่ RNA ทุกประเภทถูกสังเคราะห์บน DNA โดยมีส่วนร่วมของเอนไซม์ - RNA polymerase Messenger RNA คิดเป็น 2-3% ของ RNA มือถือทั้งหมด, ไรโบโซม - 80-85, การขนส่ง - ประมาณ 15%
mRNA.
มันอ่านข้อมูลทางพันธุกรรมจากส่วน DNA และในรูปแบบของลำดับเบสไนโตรเจนที่คัดลอกมา ถ่ายโอนไปยังไรโบโซม ซึ่งเป็นที่สังเคราะห์โปรตีนบางชนิด โมเลกุล mRNA แต่ละโมเลกุลในลำดับนิวคลีโอไทด์และขนาดสอดคล้องกับยีนใน DNA ที่คัดลอกมา โดยเฉลี่ย mRNA มี 1500 นิวคลีโอไทด์ (75-3000) แฝดสามแต่ละตัว (สามนิวคลีโอไทด์) บน mRNA เรียกว่าโคดอนขึ้นอยู่กับ codon ที่กรดอะมิโนจะปรากฏในตำแหน่งที่กำหนดในระหว่างการสังเคราะห์โปรตีน
(ทีอาร์เอ็นเอ)มีน้ำหนักโมเลกุลค่อนข้างต่ำประมาณ 24-29 พัน
D และมีนิวคลีโอไทด์ตั้งแต่ 75 ถึง 90 ในโมเลกุล นิวคลีโอไทด์ tRNA มากถึง 10% เป็นเบสรองซึ่งเห็นได้ชัดว่าปกป้องมันจากการกระทำของเอนไซม์ไฮโดรไลติก บทบาทของ tRNA คือพวกมันถ่ายโอนกรดอะมิโนไปยังไรโบโซมและมีส่วนร่วมในกระบวนการสังเคราะห์โปรตีน กรดอะมิโนแต่ละตัวยึดติดกับ tRNA เฉพาะ กรดอะมิโนจำนวนหนึ่งมี tRNA มากกว่าหนึ่งตัว จนถึงปัจจุบัน มีการค้นพบ tRNA มากกว่า 60 ตัวที่แตกต่างกันในโครงสร้างหลัก (ลำดับเบส)
โครงสร้างทุติยภูมิของ tRNAs ทั้งหมดถูกนำเสนอในรูปแบบของใบโคลเวอร์ที่มีก้านสองเกลียวและสามเกลียวเดี่ยว) ที่ส่วนท้ายของห่วงโซ่หนึ่งมีไซต์ตัวรับ - CCA triplet ไปยัง adenine ซึ่งมีกรดอะมิโนเฉพาะติดอยู่
(อาร์อาร์เอ็นเอ). ประกอบด้วยนิวคลีโอไทด์ 120-3100 Ribosomal RNA สะสมในนิวเคลียสในนิวคลีโอลี
โปรตีนไรโบโซมถูกส่งไปยังนิวคลีโอลัสจากไซโตพลาสซึม และการก่อตัวของอนุภาคย่อยไรโบโซมที่เกิดขึ้นเองนั้นเกิดขึ้นที่นั่นโดยการรวมโปรตีนเข้ากับ rRNA ที่สอดคล้องกัน อนุภาคย่อยของไรโบโซมถูกลำเลียงเข้าด้วยกันหรือแยกจากกันผ่านรูพรุนของเยื่อหุ้มนิวเคลียสไปยังไซโตพลาสซึม ไรโบโซมเป็นออร์แกเนลล์ขนาด 20-30 นาโนเมตร
พวกมันถูกสร้างขึ้นจากอนุภาคย่อยสองอนุภาคที่มีขนาดและรูปร่างต่างกัน ในบางช่วงของการสังเคราะห์โปรตีนในเซลล์ ไรโบโซมจะถูกแบ่งออกเป็นอนุภาคย่อย
ไรโบโซมอาร์เอ็นเอทำหน้าที่เป็นเฟรมเวิร์กสำหรับไรโบโซมและอำนวยความสะดวกในการจับ mRNA เบื้องต้นกับไรโบโซมในระหว่างการสังเคราะห์โปรตีน
คำถามที่ 6 พันธะที่สร้างโครงสร้างหลักและรองของ DNA และ RNA ประเภทของ RNA
รหัสพันธุกรรมเป็นวิธีการเข้ารหัสลำดับกรดอะมิโนของโปรตีนโดยใช้ลำดับของนิวคลีโอไทด์ ซึ่งเป็นลักษณะของสิ่งมีชีวิตทั้งหมด
คุณสมบัติ: 1) รหัสพันธุกรรม แฝดสาม(กรดอะมิโนแต่ละตัวถูกเข้ารหัสโดยนิวคลีโอไทด์สามตัว) 2) ไม่ทับซ้อนกัน(แฝดแฝดเพื่อนบ้านไม่มีนิวคลีโอไทด์ทั่วไป); 3) เสื่อมโทรม(ยกเว้นเมไทโอนีนและทริปโตเฟน กรดอะมิโนทั้งหมดมีโคดอนมากกว่าหนึ่งตัว) 4) สากล(ส่วนใหญ่เหมือนกันสำหรับสิ่งมีชีวิตทั้งหมด); 5) ในโคดอนของกรดอะมิโนหนึ่งตัว นิวคลีโอไทด์สองตัวแรกมักจะเหมือนกัน และตัวที่สามจะแตกต่างกัน 6) มีลำดับการอ่านเชิงเส้นและมีลักษณะโดย ความเชื่อมโยงที
e. ความบังเอิญของลำดับของ codon ใน mRNA กับลำดับของกรดอะมิโนในสายโซ่โพลีเปปไทด์สังเคราะห์
วันที่ตีพิมพ์: 2014-12-08; อ่าน: 11268 | เพจละเมิดลิขสิทธิ์
studopedia.org - Studopedia.Org - 2014-2018. (0.001 น) ...
ไซโตพลาสซึมของเซลล์ประกอบด้วย RNA หน้าที่หลักสามประเภท:
- messenger RNA (mRNA) ที่ทำหน้าที่เป็นแม่แบบสำหรับการสังเคราะห์โปรตีน
- ไรโบโซม RNA (rRNA) ทำหน้าที่เป็นส่วนประกอบโครงสร้างของไรโบโซม
- ถ่ายโอน RNAs (tRNAs) ที่เกี่ยวข้องกับการแปล (การแปล) ของข้อมูล mRNA ไปยังลำดับกรดอะมิโนของโมเลกุลโปรตีน
ในนิวเคลียสของเซลล์พบ RNA นิวเคลียร์ซึ่งประกอบขึ้นจาก 4 ถึง 10% ของ RNA ของเซลล์ทั้งหมด
RNA นิวเคลียร์จำนวนมากแสดงโดยสารตั้งต้นโมเลกุลสูงของไรโบโซมและ RNA การถ่ายโอน สารตั้งต้นของ rRNA ที่มีน้ำหนักโมเลกุลสูง (28 S, 18 S และ 5 S RNA) ส่วนใหญ่จะแปลเป็นภาษาท้องถิ่นในนิวเคลียส
RNA เป็นสารพันธุกรรมหลักในไวรัสในสัตว์และพืชบางชนิด (genomic RNA) ไวรัสอาร์เอ็นเอส่วนใหญ่มีลักษณะเฉพาะจากการถอดรหัสยีนอาร์เอ็นเอแบบย้อนกลับ กำกับโดยรีเวิร์สทรานสคริปเทส
กรดไรโบนิวคลีอิกทั้งหมดเป็นพอลิเมอร์ของไรโบนิวคลีโอไทด์ที่เชื่อมต่อกัน เช่นเดียวกับในโมเลกุลดีเอ็นเอ โดยพันธะ 3′,5′-ฟอสโฟโรไดเอสเทอร์
RNA ต่างจาก DNA ซึ่งมีโครงสร้างเป็นเกลียวคู่ RNA เป็นโมเลกุลโพลีเมอร์เชิงเส้นตรงที่มีสายเดี่ยว
โครงสร้างเอ็มอาร์เอ็นเอ mRNA เป็นคลาส RNA ที่ต่างกันมากที่สุดในแง่ของขนาดและความเสถียร
โครงสร้าง tRNA.
การถ่ายโอน RNA ทำหน้าที่เป็นสื่อกลาง (อะแดปเตอร์) ระหว่างการแปล mRNA พวกมันคิดเป็นประมาณ 15% ของ RNA ของเซลล์ทั้งหมด กรดอะมิโนที่สร้างโปรตีน 20 ชนิดแต่ละชนิดมี tRNA ของตัวเอง สำหรับกรดอะมิโนบางตัวที่เข้ารหัสโดยโคดอนตั้งแต่สองตัวขึ้นไป จะมี tRNA หลายตัว
tRNA เป็นโมเลกุลสายเดี่ยวที่ค่อนข้างเล็กซึ่งประกอบด้วยนิวคลีโอไทด์ 70-93 ตัว น้ำหนักโมเลกุลของมันคือ (2.4-3.1) .104 kDa
โครงสร้างทุติยภูมิของ tRNA เกิดขึ้นจากการก่อตัวของพันธะไฮโดรเจนจำนวนสูงสุดระหว่างคู่เสริมภายในโมเลกุลของเบสไนโตรเจน
อันเป็นผลมาจากการก่อตัวของพันธะเหล่านี้ สายโซ่โพลีนิวคลีโอไทด์ของ tRNA จะบิดตัวด้วยการก่อตัวของกิ่งก้านเกลียวที่สิ้นสุดในลูปของนิวคลีโอไทด์ที่ไม่มีการจับคู่ ภาพเชิงพื้นที่ของโครงสร้างทุติยภูมิของ tRNA ทั้งหมดมีรูปร่างเหมือนใบโคลเวอร์ลีฟ
สี่สาขาบังคับมีความโดดเด่นใน "โคลเวอร์ลีฟ" tRNA ที่ยาวกว่านอกจากนี้ยังมีสาขาสั้นที่ห้า (เพิ่มเติม)
ฟังก์ชันอะแด็ปเตอร์ของ tRNA ถูกจัดเตรียมโดยสาขาของตัวรับ โดยที่ส่วนปลาย 3' ซึ่งจะมีกรดอะมิโนตกค้างติดอยู่ด้วยพันธะอีเทอร์ และกิ่งแอนติโคดอนที่อยู่ตรงข้ามกับกิ่งของตัวรับ ที่ด้านบนสุดมีลูปที่ประกอบด้วย แอนติโคดอน
แอนติโคดอนคือนิวคลีโอไทด์สามเท่าที่เฉพาะเจาะจงซึ่งประกอบกันในทิศทางตรงกันข้ามกับโคดอน mRNA ซึ่งเข้ารหัสกรดอะมิโนที่สอดคล้องกัน
ที-แบรนช์ซึ่งมีห่วงซูโดริดีน (TyC-loop) รับรองการทำงานร่วมกันของ tRNA กับไรโบโซม
D-branch ที่มีลูปดีไฮโดรริดีนทำให้แน่ใจว่าการทำงานร่วมกันของ tRNA กับการสังเคราะห์ aminoacyl-tRNA ที่สอดคล้องกัน

โครงสร้างรองของ tRNA
หน้าที่ของกิ่งเพิ่มเติมที่ห้ายังไม่ค่อยเข้าใจ เป็นไปได้มากว่าจะทำให้ความยาวของโมเลกุล tRNA ต่างกันเท่ากัน
โครงสร้างตติยภูมิของ tRNA มีขนาดกะทัดรัดมากและเกิดขึ้นจากการรวมกิ่งของใบโคลเวอร์แต่ละกิ่งเข้าด้วยกันเนื่องจากพันธะไฮโดรเจนเพิ่มเติมเพื่อสร้างโครงสร้าง "โค้งงอศอก" รูปตัว L
การขนส่ง RNA โครงสร้างและกลไกการทำงาน
ในกรณีนี้ แขนรับซึ่งจับกรดอะมิโนจะอยู่ที่ปลายด้านหนึ่งของโมเลกุล และสารต้านโคดอนอยู่ที่ปลายอีกด้านหนึ่ง

โครงสร้างตติยของ tRNA (ตาม A.S. Spirin)
โครงสร้างของ rRNA และไรโบโซม. ไรโบโซมอาร์เอ็นเอสร้างกระดูกสันหลังที่โปรตีนจำเพาะจับกันเป็นไรโบโซม ไรโบโซมเป็นออร์แกเนลล์ของนิวคลีโอโปรตีนที่ให้การสังเคราะห์โปรตีนจาก mRNA
จำนวนไรโบโซมในเซลล์มีมาก: จาก 104 ในโปรคาริโอตถึง 106 ในยูคาริโอต ไรโบโซมมีการแปลเป็นภาษาท้องถิ่นเป็นหลักในไซโตพลาสซึม ในยูคาริโอต นอกจากนี้ ในนิวเคลียส ในเมทริกซ์ไมโตคอนเดรีย และในสโตรมาของคลอโรพลาสต์ ไรโบโซมประกอบด้วยอนุภาคย่อยสองส่วน: ใหญ่และเล็ก ตามขนาดและน้ำหนักโมเลกุล ไรโบโซมที่ศึกษาทั้งหมดแบ่งออกเป็น 3 กลุ่ม - ไรโบโซม 70S ของโปรคาริโอต (ค่าสัมประสิทธิ์การตกตะกอน S) ประกอบด้วยอนุภาคย่อย 30S ขนาดเล็กและ 50S ขนาดใหญ่ ไรโบโซมยูคาริโอต 80S ประกอบด้วยหน่วยย่อยขนาดเล็ก 40S และหน่วยย่อยขนาดใหญ่ 60S
หน่วยย่อยขนาดเล็กของไรโบโซม 80S ถูกสร้างขึ้นโดยโมเลกุล rRNA (18S) หนึ่งโมเลกุลและ 33 โมเลกุลของโปรตีนต่างๆ
หน่วยย่อยขนาดใหญ่ประกอบด้วยโมเลกุล rRNA สามโมเลกุล (5S, 5.8S และ 28S) และโปรตีนประมาณ 50 ชนิด
โครงสร้างทุติยภูมิของ rRNA เกิดขึ้นเนื่องจากส่วนที่สั้นสองเกลียวของโมเลกุล - กิ๊บติดผม (ประมาณ 2/3 ของ rRNA) 1/3 - แสดงโดยส่วนที่เป็นเกลียวเดี่ยวที่อุดมไปด้วยนิวคลีโอไทด์ purine
ปุ่มโซเชียลสำหรับ Joomla
โปรตีนเป็นพื้นฐานของชีวิต หน้าที่ของพวกมันในเซลล์นั้นมีความหลากหลายมาก อย่างไรก็ตาม โปรตีน "ไม่สามารถ" สืบพันธุ์ได้ และข้อมูลทั้งหมดเกี่ยวกับโครงสร้างของโปรตีนมีอยู่ในยีน (DNA)
ในสิ่งมีชีวิตชั้นสูง โปรตีนจะถูกสังเคราะห์ในไซโตพลาสซึมของเซลล์ และ DNA ถูกซ่อนอยู่หลังเปลือกของนิวเคลียส ดังนั้น DNA จึงไม่สามารถใช้เป็นแม่แบบในการสังเคราะห์โปรตีนได้โดยตรง บทบาทนี้ดำเนินการโดยกรดนิวคลีอิกอื่น - RNA
โมเลกุลอาร์เอ็นเอเป็นพอลินิวคลีโอไทด์ที่ไม่มีการแยกย่อยซึ่งมีโครงสร้างตติยภูมิ
มันถูกสร้างขึ้นโดยสายโซ่พอลินิวคลีโอไทด์หนึ่งสาย และถึงแม้ว่านิวคลีโอไทด์เสริมที่รวมอยู่ในนั้นก็สามารถสร้างพันธะไฮโดรเจนระหว่างกันได้เช่นกัน แต่พันธะเหล่านี้เกิดขึ้นระหว่างนิวคลีโอไทด์ของสายโซ่เดียว RNA chain นั้นสั้นกว่า DNA chain มาก หากเนื้อหาของ DNA ในเซลล์ค่อนข้างคงที่ แสดงว่าเนื้อหาของ RNA จะผันผวนอย่างมาก มีการสังเกตจำนวน RNA ในเซลล์มากที่สุดในระหว่างการสังเคราะห์โปรตีน
RNA มีบทบาทสำคัญในการส่งและการนำข้อมูลทางพันธุกรรมไปใช้
ตามหน้าที่และลักษณะโครงสร้าง RNA มือถือหลายชั้นมีความโดดเด่น
RNA ของเซลล์มีสามประเภทหลัก
- ให้ข้อมูล (mRNA) หรือเมทริกซ์ (mRNA) โมเลกุลของมันมีความหลากหลายมากที่สุดในแง่ของขนาด น้ำหนักโมเลกุล (ตั้งแต่ 0.05x106 ถึง 4x106) และความเสถียร
พวกมันประกอบขึ้นประมาณ 2% ของจำนวน RNA ทั้งหมดในเซลล์ mRNA ทั้งหมดเป็นพาหะของข้อมูลทางพันธุกรรมตั้งแต่นิวเคลียสไปจนถึงไซโตพลาสซึม ไปจนถึงบริเวณที่มีการสังเคราะห์โปรตีน พวกมันทำหน้าที่เป็นเมทริกซ์ (รูปวาดการทำงาน) สำหรับการสังเคราะห์โมเลกุลโปรตีน เนื่องจากพวกมันกำหนดลำดับกรดอะมิโน (โครงสร้างหลัก) ของโมเลกุลโปรตีน

- ไรโบโซม RNA (rRNA)
พวกมันคิดเป็น 80–85% ของเนื้อหา RNA ทั้งหมดในเซลล์
31. โครงสร้างของอาร์เอ็นเอ ประเภท RNA ลักษณะโครงสร้างและหน้าที่ โครงสร้างรองของ tRNA
Ribosomal RNA ประกอบด้วยนิวคลีโอไทด์ 3-5 พันตัว มันถูกสังเคราะห์ในนิวเคลียสของนิวเคลียส ในความซับซ้อนที่มีโปรตีนไรโบโซม rRNA จะสร้างไรโบโซม - ออร์แกเนลล์ที่ประกอบโมเลกุลโปรตีน ความสำคัญหลักของ rRNA คือมันให้การจับเริ่มต้นของ mRNA และไรโบโซม และสร้างศูนย์กลางที่แอคทีฟของไรโบโซม ซึ่งพันธะเปปไทด์จะเกิดขึ้นระหว่างกรดอะมิโนในระหว่างการสังเคราะห์สายโซ่โพลีเปปไทด์
- โอน RNAs (tRNAs)
โมเลกุล tRNA มักจะมีนิวคลีโอไทด์ 75-86 น้ำหนักโมเลกุลของโมเลกุล tRNA อยู่ที่ประมาณ 25,000 โมเลกุล tRNA มีบทบาทเป็นตัวกลางในการสังเคราะห์โปรตีน - พวกมันส่งกรดอะมิโนไปยังบริเวณที่มีการสังเคราะห์โปรตีนซึ่งก็คือไรโบโซม เซลล์ประกอบด้วย tRNA มากกว่า 30 ชนิด tRNA แต่ละประเภทมีลำดับนิวคลีโอไทด์เฉพาะของตัวเอง
อย่างไรก็ตาม โมเลกุลทั้งหมดมีส่วนเสริมภายในโมเลกุลหลายส่วน เนื่องจากมี tRNA ทั้งหมดมีโครงสร้างตติยภูมิที่มีรูปร่างคล้ายใบโคลเวอร์
โครงสร้างรองของ RNA- ลักษณะของ tRNA เป็นเกลียวเดี่ยว มีรูปร่างเหมือน "ใบโคลเวอร์"
รวมถึง:
- เกลียวคู่ที่ค่อนข้างสั้น - ลำต้น
- ส่วนที่เป็นเกลียวเดียว - ลูป
มี 4 ก้าน (ตัวรับ, แอนติโคดอน, ไดไฮโดรริดิล, ซูโดริดิล) และ 3 ลูป

"Stem-loop" - องค์ประกอบของโครงสร้างทุติยภูมิของ RNA แผนผัง

"Pseudoknot" - องค์ประกอบของโครงสร้างรองของ RNA แผนผัง

ก้านตัวรับประกอบด้วยปลายสายโพลีนิวคลีโอไทด์ 3 ฟุตและ 5 นิ้ว ส่วนปลาย 5 นิ้วมีกรดกัวนีลิกตกค้าง ส่วนปลาย 3 นิ้วเป็นทริปเปิลซีเอและทำหน้าที่สร้างพันธะเอสเทอร์กับ AA
ก้านแอนติโคดอนรับรู้โคดอนของมันบน mRNA ในไรโบโซมโดยหลักการของการเติมเต็ม
ก้านเทียมทำหน้าที่ยึดกับไรโบโซม
ก้านไดไฮโดรริดิลทำหน้าที่จับกับอะมิโนอะซิล-tRNA ซินธิเทส
ปุ่มโซเชียลสำหรับ Joomla





