Daerah fungsional tRNA. Struktur dan fungsi tRNA, fitur aktivasi asam amino
70-90N | halaman kedua - daun semanggi | CCA 3" const untuk semua tRNA |
kehadiran timin, pseudouridine-psi, digirouridine DGU di D-loop - perlindungan terhadap ribonuklease? berumur panjang | Berbagai struktur primer tRNA - 61 + 1 - dengan jumlah kodon + formilmetionin tRNA, antikodon kucing sama dengan metionin tRNA. Ragam struktur tersier - 20 (sesuai dengan jumlah asam amino) | pengenalan - pembentukan ikatan kovalen m-y tRNA dan aksi | sintetase aminoasil-tRNA menempel pada tRNA
Fungsi tRNA adalah untuk mentransfer asam amino dari sitoplasma ke ribosom, di mana sintesis protein terjadi.
tRNA yang mengikat satu asam amino disebut isoakseptor.
Secara total, 64 tRNA berbeda secara bersamaan ada dalam sel.
Setiap tRNA hanya berpasangan dengan kodonnya sendiri.
Setiap tRNA mengenali kodonnya sendiri tanpa melibatkan asam amino. Asam amino yang terikat pada tRNA dimodifikasi secara kimia, setelah itu polipeptida yang dihasilkan, yang mengandung asam amino yang dimodifikasi, dianalisis. Sisteinil-tRNACys (R=CH2-SH) direduksi menjadi alanil-tRNACys (R=CH3).
Kebanyakan tRNA, terlepas dari urutan nukleotidanya, memiliki struktur sekunder berbentuk daun semanggi karena adanya tiga jepit rambut di dalamnya.
Fitur struktural tRNA
Selalu ada empat nukleotida yang tidak berpasangan di ujung 3 "molekul, dan tiga di antaranya pasti CCA. Ujung rantai RNA 5" dan 3" membentuk batang akseptor. Rantai disatukan karena pasangan komplementer dari tujuh nukleotida 5" - ujung dengan tujuh nukleotida yang terletak di dekat ujung 3 ". 2. Semua molekul memiliki jepit rambut T? C, dinamakan demikian karena mengandung dua residu yang tidak biasa: ribothymidine (T) dan pseudouridine (? Jepit rambut terdiri dari rangkap dua -batang beruntai dari lima basa berpasangan, termasuk pasangan G-C, dan loop dengan tujuh nukleotida panjangnya.
pada titik yang sama dalam loop. 3. Dalam jepit rambut antikodon, batang selalu diwakili oleh keluarga berpasangan
alasan. Triplet komplementer dengan kodon terkait, antikodon, terletak di loop.
le, terdiri dari tujuh nukleotida. Sebuah ura invarian
cyl dan sitosin yang dimodifikasi, dan purin yang dimodifikasi berdampingan dengan ujung 3 ", sebagai aturan
adenin. 4. Jepit rambut lainnya terdiri dari tangkai tiga sampai empat pasang nukleotida panjang dan loop variabel
ukuran, sering mengandung urasil dalam bentuk tereduksi - dihydrouracil (DU). Urutan nukleotida batang, jumlah nukleotida antara batang antikodon dan batang T?C (loop variabel), serta ukuran loop dan lokalisasi residu dihydrouracil di loop DU sangat bervariasi.
[Penyanyi, 1998].
Struktur tersier tRNA
Struktur berbentuk L.
Pengikatan asam amino pada tRNA
Agar asam amino dapat membentuk rantai polipeptida, asam amino harus diikat ke tRNA oleh enzim aminoasil-tRNA sintetase. Enzim ini membentuk ikatan kovalen antara gugus karboksil asam amino dan gugus hidroksil ribosa pada ujung 3' tRNA dengan partisipasi ATP. Aminoasil-tRNA sintetase mengenali kodon spesifik bukan karena adanya antikodon pada tRNA, tetapi dengan adanya situs pengenalan spesifik pada tRNA.
Secara total, ada 21 sintetase aminoasil-tRNA yang berbeda di dalam sel.
Bergabung terjadi dalam dua tahap:
1. Gugus karboksil dari asam amino terikat pada ATP a-fosfat. Aminoasil adenilat yang tidak stabil yang dihasilkan distabilkan dengan mengikat enzim.
2. Transfer gugus aminoasil dari aminoasil adenilat ke gugus 2' atau 3'-OH dari ribosa terminal tRNA
Beberapa sintetase aminoasil-tRNA terdiri dari rantai polipeptida tunggal, sementara yang lain terdiri dari dua atau empat rantai identik, masing-masing dengan berat molekul 35 hingga 115 kDa. Beberapa enzim dimer dan tetramerik terdiri dari dua jenis subunit. Tidak ada korelasi yang jelas antara ukuran molekul enzim atau sifat struktur subunit dan spesifisitasnya.
Spesifisitas suatu enzim ditentukan oleh ikatannya yang kuat pada ujung akseptor tRNA, wilayah DU, dan loop variabel. Beberapa enzim tampaknya tidak mengenali triplet antikodon dan mengkatalisis reaksi aminoasetilasi bahkan ketika antikodon diubah. Namun, beberapa enzim menunjukkan aktivitas yang berkurang sehubungan dengan tRNA yang dimodifikasi tersebut dan menambahkan asam amino yang salah saat mengganti antikodon.
70-90n | halaman kedua - daun semanggi | CCA 3" const untuk semua tRNA |
kehadiran timin, pseudouridine-psi, digirouridine DGU di D-loop - perlindungan terhadap ribonuklease? berumur panjang | Berbagai struktur primer tRNA - 61 + 1 - dengan jumlah kodon + formilmetionin tRNA, antikodon kucing sama dengan metionin tRNA. Ragam struktur tersier - 20 (sesuai dengan jumlah asam amino)
Ada dua jenis tRNA yang mengikat metionin tRNAFMet dan tRNAMMet pada prokariota dan tRNAIMet dan tRNAMMet pada eukariota. Metionin ditambahkan ke setiap tRNA menggunakan sintesis aminoasil-tRNA yang sesuai. metionin melekat pada tRNAFMet dan tRNAIMet dibentuk oleh enzim methionyl-tRNA-transformylase menjadi Fmet-tRNAFMet. tRNA yang sarat dengan formilmetionin mengenali kodon inisiasi AUG.
Literatur:
Sayangnya, tidak ada daftar pustaka.
Buku pelajaran. Terlepas dari kenyataan bahwa tRNA jauh lebih kecil, cerita tentang struktur, fitur, dan fungsinya layak mendapat bab terpisah.
Jadi, tRNA adalah "adaptor", yang mengenali urutan tiga huruf dari kode genetik di satu ujung, mencocokkannya dengan satu-satunya asam amino yang sesuai yang dipasang di ujung lain tRNA. Pada ujung RNA transfer yang menyentuh RNA pembawa pesan, terdapat 3 nukleotida yang terbentuk antikodon. Hanya jika antikodon melengkapi wilayah mRNA, RNA transfer dapat bergabung dengannya. Tetapi bahkan dalam kasus ini, tRNA tidak dapat bergabung dengan mRNA sendiri; ia membutuhkan bantuan ribosom, yang merupakan tempat interaksi mereka, serta peserta aktif dalam terjemahan. Misalnya, ribosom yang menciptakan ikatan antara asam amino yang dibawa oleh tRNA, membentuk rantai protein.
Fitur struktural tRNA ditentukan oleh kode genetik, yaitu aturan untuk membangun protein menurut gen yang dibaca oleh RNA transfer. Kode ini bekerja di setiap makhluk hidup di Bumi: penciptaan virus ditulis dengan kodon tiga huruf yang sama yang digunakan untuk menulis "instruksi perakitan" lumba-lumba. Telah diverifikasi secara eksperimental bahwa gen dari satu makhluk hidup, yang ditempatkan di sel makhluk lain, disalin dengan sempurna dan diterjemahkan ke dalam protein yang tidak dapat dibedakan dari gen yang bereproduksi dalam sel inangnya. Keseragaman kode genetik adalah dasar untuk produksi E. coli yang dimodifikasi oleh koloni insulin dan banyak enzim manusia lainnya yang digunakan sebagai obat untuk orang yang tubuhnya tidak mampu memproduksinya, atau produksinya tidak mencukupi. Terlepas dari perbedaan yang jelas antara manusia dan E. coli, protein manusia dengan mudah dibuat dari cetak biru manusia menggunakan mesin fotokopi E. coli. Tidak mengherankan, RNA transfer dari makhluk yang berbeda sangat sedikit berbeda.

Setiap kodon dari daftar ini, kecuali tiga hentikan kodon, menandakan selesainya terjemahan, harus dikenali oleh RNA transfer. Pengenalan dilakukan dengan menempelkan antikodon pada messenger RNA, yang hanya dapat mengikat satu kodon dari daftar, sehingga tRNA hanya dapat mengenali satu kodon. Ini berarti setidaknya ada 61 jenis molekul ini di dalam sel. Bahkan, ada lebih banyak dari mereka, karena dalam beberapa situasi untuk membaca RNA pembawa pesan tidak cukup hanya memiliki antikodon yang tepat: diperlukan kondisi lain, yang dengannya tRNA khusus yang dimodifikasi dibuat.
Sepintas, keragaman tRNA seperti itu seharusnya secara signifikan memperumit proses translasi: lagi pula, masing-masing molekul ini akan memeriksa kodon RNA matriks yang digantikan oleh ribosom untuk kesesuaian dengan antikodonnya - tampaknya begitu banyak pekerjaan mekanis yang sia-sia. , begitu banyak membuang waktu dan tenaga. Tetapi sebagai hasil evolusi, mekanisme seluler juga telah terbentuk yang mencegah masalah ini. Misalnya, jumlah tRNA dari setiap spesies dalam sel sesuai dengan seberapa sering asam amino yang dibawa oleh spesies itu ditemukan dalam protein yang sedang dibangun. Ada asam amino yang jarang digunakan oleh sel, dan ada yang sering digunakan, dan jika jumlah tRNA yang membawanya sama, ini akan sangat mempersulit perakitan protein. Oleh karena itu, ada beberapa asam amino "langka" dan tRNA yang sesuai di dalam sel, sementara asam amino yang sering muncul diproduksi dalam jumlah besar.

Dengan berbagai macam molekul tRNA, mereka semua sangat mirip, oleh karena itu, dengan mempertimbangkan struktur dan fungsinya, kami terutama akan mempelajari fitur-fitur umum untuk semua spesies. Ketika Anda melihat tata letak 3D tRNA, itu terlihat seperti tumpukan atom yang padat. Tampaknya luar biasa bahwa molekul melingkar yang rumit ini adalah hasil dari pelipatan rantai panjang nukleotida, tetapi begitulah cara pembentukannya.
Dimungkinkan untuk melacak tahapan pembentukannya, mulai dari yang pertama: penyusunan urutan nukleotida oleh RNA polimerase sesuai dengan gen yang mengandung informasi tentang RNA transfer ini. Urutan nukleotida-nukleotida ini saling mengikuti dan jumlahnya disebut struktur utama tRNA. Ternyata itu adalah struktur utama tRNA yang dikodekan dalam gen yang dibaca oleh RNA polimerase. Secara umum, struktur primer adalah urutan molekul yang relatif sederhana dari jenis yang sama, yang terdiri dari molekul polimer terlipat yang lebih kompleks. Misalnya, struktur utama molekul protein adalah urutan sederhana dari asam amino penyusunnya.

Rantai nukleotida apa pun tidak dapat berada dalam keadaan tidak terlipat di dalam sel, hanya direntangkan dalam satu garis. Ada terlalu banyak bagian bermuatan positif dan negatif di tepi nukleotida, yang dengan mudah membentuk ikatan hidrogen satu sama lain. Bagaimana ikatan yang sama terbentuk antara nukleotida dari dua molekul DNA, menghubungkannya menjadi heliks ganda, dijelaskan dalam, dan Anda dapat masuk ke detail tentang ikatan hidrogen. Ikatan hidrogen kurang kuat dibandingkan ikatan antar atom dalam molekul, tetapi ikatan tersebut cukup untuk memutar benang tRNA secara rumit dan mempertahankannya pada posisi tersebut. Pada awalnya, ikatan ini hanya terbentuk di antara beberapa nukleotida, melipat tRNA menjadi bentuk daun semanggi. Hasil dari pelipatan awal ini disebut struktur sekunder tRNA. Diagram di sebelah kiri menunjukkan bahwa hanya beberapa nukleotida yang dihubungkan oleh ikatan hidrogen, sementara yang lain tetap tidak berpasangan, membentuk cincin dan loop. Perbedaan antara struktur sekunder dari berbagai jenis tRNA disebabkan oleh perbedaan struktur primernya. Ini memanifestasikan dirinya dalam panjang yang berbeda dari "daun semanggi" atau "tangkai" karena panjang yang berbeda dari rantai awal nukleotida.
Perbedaan lain dalam struktur utama tRNA yang berbeda adalah bahwa hanya di beberapa posisi mereka memiliki nukleotida yang sama (dalam diagram di atas mereka ditandai dengan huruf pertama dari nama mereka), sementara sebagian besar nukleotida dalam tRNA yang berbeda berbeda satu sama lain. Skema di atas umum untuk semua tRNA, jadi nukleotida yang berbeda ditandai dengan angka.
Bagian fungsional utama tRNA adalah:
=) antikodon, yaitu urutan nukleotida yang komplementer dengan kodon tunggal RNA messenger yang terletak di jepit rambut antikodon
=) ujung akseptor dimana asam amino dapat dilampirkan. Itu terletak di sisi berlawanan dari jepit rambut antikodon.
Pada kenyataannya, tidak ada satu tRNA pun yang terlihat seperti pada diagram struktur sekunder, karena hanya beberapa nukleotida yang bergabung bersama untuk membentuknya, sedangkan sisanya tetap tidak berpasangan. Karena pembentukan ikatan hidrogen antara nukleotida dari berbagai bagian daun semanggi, lipatannya lebih jauh menjadi lebih kompleks. struktur tersier dalam bentuk L. Anda dapat memahami dengan tepat bagaimana bagian-bagian berbeda dari struktur sekunder melengkung untuk membentuk struktur tersier dengan mencocokkan warna dalam diagram di bawah ini. Jepit rambut antikodon, ditandai dengan warna biru dan abu-abu, tetap berada di bawah (perlu diingat bahwa "bawah" ini bersyarat: lebih mudah untuk menggambarkan tRNA dalam orientasi spasial ini dalam skema terjemahan protein), dan ujung akseptor (kuning) ditekuk ke samping.

Ini adalah tampilan tRNA ketika siap untuk melampirkan asam amino. tRNA tidak dapat bergabung dengan asam amino sendiri, ini membutuhkan partisipasi enzim khusus: sintetase aminoasil-tRNA. Jumlah jenis sintetase dalam sel bertepatan dengan jumlah jenis tRNA.
Keseragaman bentuk semua jenis tRNA diperlukan agar ribosom dapat mengenali salah satu dari mereka, memfasilitasi docking mereka dengan mRNA, dan bergerak di dalam dirinya sendiri dari satu situs ke situs lainnya. Jika berbagai jenis tRNA berbeda secara signifikan satu sama lain, ini akan membuat pekerjaan ribosom menjadi sangat sulit, secara kritis mengurangi laju sintesis protein. Seleksi alam dengan demikian bertujuan untuk membuat tRNA serupa satu sama lain. Tetapi pada saat yang sama, ada faktor lain yang mengharuskan adanya perbedaan nyata antara berbagai jenis tRNA: bagaimanapun, perlu untuk mengenali setiap jenis dan menempelkannya satu-satunya asam amino yang sesuai. Jelas, perbedaan ini harus terlihat, tetapi tidak terlalu signifikan, sehingga pekerjaan mengenali spesies tRNA berubah menjadi proses perhiasan. Dan justru inilah yang dilakukan oleh sintetase aminoasil-tRNA: masing-masing hanya dapat mengikat satu dari 20 asam amino dan menempelkannya tepat pada jenis tRNA yang sesuai dengan asam amino ini. Dari tabel dengan kode genetik dapat dilihat bahwa setiap asam amino dikodekan oleh beberapa urutan nukleotida, oleh karena itu, misalnya, keempat tRNA dengan antikodon CGA, CGG, CGU dan CGC akan dikenali oleh sintetase yang sama yang mengikat alanin. ke mereka. TRNA semacam itu yang diproses oleh satu sintetase disebut terkait.
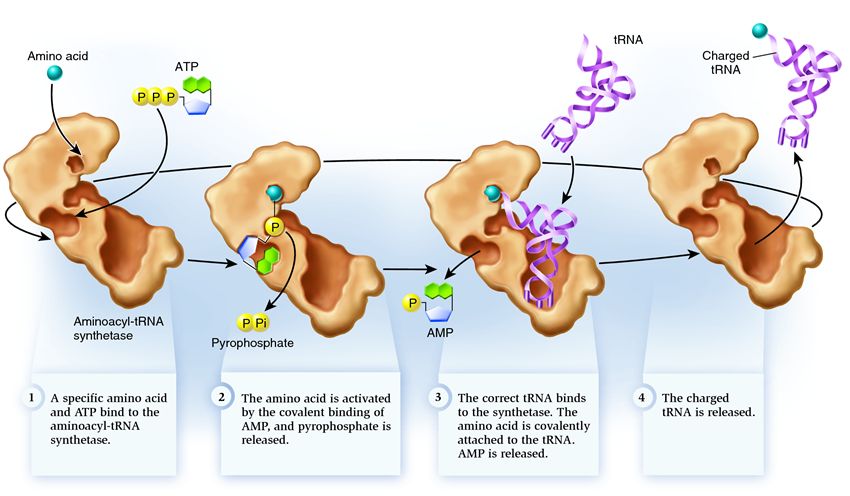
Sintetase termasuk dalam kelompok enzim yang fungsinya mengikat molekul yang ada secara terpisah dan menggabungkannya menjadi satu:
1 . sintetase menghubungkan asam amino dan molekul ATP. Dua gugus fosfat melepaskan diri dari ATP, melepaskan energi yang dibutuhkan untuk aktivitas berikut. Adenosin monofosfat (AMP) yang tersisa dari molekul yang dihancurkan menempel pada asam amino, mempersiapkannya untuk dihubungkan dengan jepit rambut akseptor.
2 . sintetase menempel pada dirinya sendiri salah satu tRNA terkait yang sesuai dengan asam amino ini.

Pada tahap ini, kesesuaian RNA transfer ke sintetase diperiksa. Ada beberapa cara pengenalan, dan setiap sintetase memiliki kombinasi unik di antaranya. Setidaknya satu nukleotida antikodon terlibat dalam interaksi antara sintetase dan tRNA. Jepit rambut akseptor juga perlu diperiksa: keberadaan nukleotida spesifik di atasnya yang umum untuk tRNA terkait yang sesuai dengan asam amino yang diinginkan ditentukan. Nukleotida dari bagian lain tRNA juga dapat berpartisipasi dalam pencocokan dengan mengikat situs sintetase tertentu. TRNA yang salah mungkin cocok dengan yang diinginkan dalam beberapa cara, tetapi karena pencocokan yang tidak lengkap, tRNA akan bergabung dengan sintetase secara perlahan dan longgar, dengan mudah jatuh. Dan tRNA yang benar akan menempel pada sintetase dengan cepat dan kuat, akibatnya struktur sintetase berubah, memulai proses aminoasilasi , yaitu perlekatan asam amino pada tRNA.

3 . aminoasilasi terdiri dari penggantian molekul AMP yang terikat pada asam amino dengan molekul tRNA. Setelah penggantian ini, AMP meninggalkan sintetase dan tRNA ditahan untuk pemeriksaan asam amino terakhir. Jika asam amino yang melekat diakui sebagai salah, itu akan terlepas dari tRNA, tempat asam amino di sintetase akan kosong, dan molekul lain dapat bergabung di sana. Asam amino baru akan melalui tahapan koneksi dengan ATP dan tRNA, dan juga akan diuji. Jika tidak ada kesalahan yang dibuat, tRNA yang bermuatan asam amino dilepaskan: ia siap memainkan perannya dalam translasi protein. Dan sintetase siap untuk melampirkan asam amino dan tRNA baru, dan siklus akan dimulai lagi.
Banyak tergantung pada operasi yang benar dari sintetase aminoasil-tRNA: jika kegagalan terjadi pada tahap ini, maka asam amino yang salah akan melekat pada tRNA. Dan itu akan dibangun ke dalam protein yang tumbuh di ribosom, karena tRNA dan ribosom tidak memiliki fungsi untuk memeriksa kesesuaian kodon dan asam amino. Konsekuensi dari kesalahan bisa kecil atau bencana, dan melalui seleksi alam, makhluk dengan enzim yang tidak memiliki fungsi pemeriksaan tersebut telah digantikan oleh yang lebih mudah beradaptasi dengan berbagai pilihan untuk pencocokan antara asam amino dan tRNA. Oleh karena itu, dalam sel modern, sintetase bergabung dengan asam amino yang salah rata-rata dalam satu kasus dari 50 ribu, dan dengan tRNA yang salah hanya sekali dalam 100 ribu perlekatan.

Beberapa asam amino berbeda satu sama lain hanya dengan beberapa atom. Jika Anda melihat skema mereka, menjadi jelas bahwa kemungkinan membingungkan arginin dengan alanin jauh lebih sedikit daripada membingungkan isoleusin untuk leusin atau valin. Oleh karena itu, setiap sintetase yang mengikat salah satu asam amino yang mirip satu sama lain memiliki mekanisme verifikasi tambahan. Berikut adalah contoh adaptasi seperti itu dalam sintetase isoleusin-tRNA:

Setiap sintase memiliki pusat sintetis dimana asam amino terikat pada tRNA. Jepit rambut akseptor tRNA yang ditangkap oleh sintetase pergi ke sana, seperti halnya asam amino yang siap bereaksi dengannya. Pekerjaan beberapa sintetase berakhir segera setelah hubungan asam amino dan tRNA. Tetapi Ile-tRNA sintetase memiliki peluang yang lebih besar untuk membuat kesalahan karena adanya asam amino mirip isoleusin lainnya. Oleh karena itu, dia juga memiliki lembaga pemasyarakatan: dari namanya sudah jelas apa perannya dalam proses penyambungan tRNA dan asam amino. Gambar di sebelah kanan menunjukkan bahwa posisi ujung jepit rambut akseptor tRNA di pusat sintetik Ile-tRNA sintetase membuat jepit rambut ini melengkung tidak wajar. Namun, sintetase menahan tRNA pada posisi ini sampai asam amino melekat padanya. Setelah hubungan ini terjadi, kebutuhan untuk menemukan jepit rambut akseptor di pusat sintetis habis, dan tRNA diluruskan, mendapatkan ujungnya dengan asam amino yang melekat padanya ke pusat koreksi.

Tentu saja, pusat sintetik juga berperan dalam menyaring asam amino yang tidak cocok untuk sintetase. Untuk masuk ke dalamnya, molekul tersebut harus memenuhi sejumlah syarat, termasuk memiliki ukuran yang pas. Terlepas dari kenyataan bahwa leusin dan isoleusin mengandung jumlah atom yang sama, karena perbedaan struktur spasial, leusin lebih besar. Oleh karena itu, ia tidak dapat menembus ke pusat sintetik, yang ukurannya sesuai dengan isoleusin yang lebih kompak, dan hanya memantul dari sintetase Ile-tRNA.
Tetapi valin, yang merupakan molekul terkecil dari ketiga molekul ini dengan struktur atom yang serupa, dengan mudah menggantikan isoleusin di pusat sintetik, dan sintetase menempelkannya pada tRNA. Dalam hal ini pusat pemasyarakatan sintetase ikut bermain. Jika jepit rambut akseptor pelurus diisi dengan benar dan membawa isoleusin, maka ia tidak dapat masuk ke dalam pusat koreksi: ia terlalu kecil untuk molekul ini. Dengan demikian, tRNA yang diluruskan tidak lagi dipegang oleh apa pun, dan terlepas dari sintetase. Tetapi jika valin dilekatkan pada tRNA, ia akan masuk ke pusat koreksi, sehingga tRNA tetap terhubung dengannya di sintetase. Tinggal terlalu lama tRNA di dalam adalah sinyal kesalahan untuk sintetase, mengubah konfigurasi spasialnya. Hasil dari:
=) valin terlepas dari tRNA dan dikeluarkan dari sintetase
=) jepit rambut akseptor kembali ke situs sintetis, menunggu perlekatan ke asam amino
=) sintetase mengikat asam amino baru, "mengisi" tRNA dengannya dan sekali lagi memeriksa apakah isoleusin digunakan untuk ini.
Mekanisme pengenalan ganda yang serupa digunakan oleh sintetase lain.
Sifat fisik dan kimia DNA
Berbagai faktor yang memutuskan ikatan hidrogen (peningkatan suhu di atas 80 C, perubahan pH dan kekuatan ionik, kerja urea, dll.) menyebabkan denaturasi DNA, mis. perubahan susunan spasial rantai DNA tanpa memutuskan ikatan kovalen. Heliks ganda DNA selama denaturasi sepenuhnya atau sebagian dibagi menjadi rantai komponennya. Denaturasi DNA disertai dengan peningkatan penyerapan optik di wilayah UV basa purin dan pirimidin. Fenomena ini disebut efek hiperkromik . Denaturasi juga mengurangi viskositas tinggi yang melekat pada larutan DNA asli. Ketika struktur DNA untai ganda asli dipulihkan, sebagai akibat dari renaturasi, penyerapan pada 260 nm oleh basa nitrogen berkurang karena "pelindung" mereka. Fenomena ini disebut efek hipokromik .
Penguraian setiap DNA ke dalam rantai komponennya dilakukan dalam kisaran suhu tertentu. Titik tengah interval ini disebut titik leleh. Suhu leleh DNA tergantung pada kondisi standar (pH tertentu dan kekuatan ionik) pada rasio basa nitrogen. Pasangan G-C yang mengandung tiga ikatan hidrogen lebih kuat, oleh karena itu, semakin tinggi kandungan pasangan G-C dalam DNA, semakin tinggi titik lelehnya.
Fungsi DNA. Dalam urutan nukleotida dalam molekul DNA, informasi genetik dikodekan. Fungsi utama DNA adalah, pertama, untuk memastikan reproduksi dirinya sendiri dalam serangkaian generasi sel dan generasi organisme, dan kedua, untuk memastikan sintesis protein. Fungsi-fungsi ini disebabkan oleh fakta bahwa molekul DNA berfungsi sebagai matriks dalam kasus pertama untuk replikasi, yaitu. menyalin informasi dalam molekul DNA anak, di detik - untuk transkripsi, mis. untuk mengkode ulang informasi ke dalam struktur RNA.
Beras. 5 Kurva leleh (denaturasi DNA)
Untaian DNA komplementer yang dipisahkan selama denaturasi, dalam kondisi tertentu, dapat terhubung kembali menjadi heliks ganda. Proses ini disebut RENATURASI. Jika denaturasi belum terjadi sepenuhnya dan setidaknya beberapa basa tidak kehilangan interaksi oleh ikatan hidrogen, renaturasi berlangsung sangat cepat.
Sitoplasma sel mengandung tiga jenis fungsional utama RNA. Ini adalah messenger RNA - mRNA yang bertindak sebagai template untuk sintesis protein, RNA ribosom - rRNA yang bertindak sebagai komponen struktural ribosom, dan transfer RNA - tRNA yang terlibat dalam translasi (translasi) informasi mRNA ke dalam urutan asam amino dalam protein.
Tabel 2 menunjukkan perbedaan antara DNA dan RNA dalam hal struktur, lokalisasi dalam sel, dan fungsi.
Tabel 2 Perbedaan antara DNA dan RNA
Transfer RNA, tRNA-asam ribonukleat, yang fungsinya untuk mengangkut AA ke tempat sintesis protein. Ini memiliki panjang khas 73-93 nukleotida dan ukuran sekitar 5 nm. tRNA juga terlibat langsung dalam pertumbuhan rantai polipeptida, bergabung - berada dalam kompleks dengan asam amino - ke kodon mRNA dan menyediakan konformasi kompleks yang diperlukan untuk pembentukan ikatan peptida baru. Setiap asam amino memiliki tRNA sendiri. tRNA adalah RNA untai tunggal, tetapi dalam bentuk fungsionalnya memiliki konformasi daun semanggi. AA terikat secara kovalen pada ujung 3" molekul menggunakan enzim aminoasil-tRNA sintetase, spesifik untuk setiap jenis tRNA. Di situs C, ada antikodon yang sesuai dengan AA-te. tRNA disintesis oleh RNA polimerase biasa di kasus prokariota dan oleh RNA polimerase III dalam kasus eukariota Transkrip gen tRNA menjalani pemrosesan bertingkat, yang mengarah pada pembentukan struktur spasial khas tRNA.
Pemrosesan tRNA melibatkan 5 langkah utama:
penghapusan urutan nukleotida pemimpin 5";
penghapusan urutan terminal 3';
menambahkan urutan CCA di ujung 3";
eksisi intron (pada eukariota dan archaea);
modifikasi nukleotida individu.
Transpor tRNA dilakukan di sepanjang jalur yang bergantung pada Ran dengan partisipasi eksport faktor transpor, yang mengenali karakteristik str-ru sekunder dan tersier dari tRNA matang: bagian untai ganda pendek dan diproses dengan benar 5 "- dan 3" berakhir. Mekanisme ini memastikan bahwa hanya tRNA matang yang diekspor dari nukleus.
62. Terjemahan - pengenalan kodon mRNA
Translasi adalah sintesis protein yang dilakukan oleh ribosom dari asam amino pada template mRNA (atau dan RNA). Unsur-unsur penyusun proses translasi: asam amino, tRNA, ribosom, mRNA, enzim untuk aminoasilasi tRNA, faktor translasi protein (faktor protein inisiasi, elongasi, terminasi - protein ekstraribosomal spesifik yang diperlukan untuk proses translasi), sumber energi ATP dan GTP , ion magnesium (menstabilkan struktur ribosom). 20 asam amino terlibat dalam sintesis protein. Agar asam amino "mengenali" tempatnya di rantai polipeptida masa depan, ia harus mengikat RNA transfer (tRNA) yang melakukan fungsi adaptor. TRNA yang mengikat asam amino kemudian mengenali kodon yang sesuai pada mRNA. pengenalan kodon mRNA:
Interaksi kodon-antikodon didasarkan pada prinsip saling melengkapi dan antiparalelisme:
3'----C - G-A*------5' antikodon tRNA
5'-----G-C-Y*------3' kodon mRNA
Hipotesis goyangan diajukan oleh F. Crick:
Basa 3' dari kodon mRNA memiliki pasangan yang tidak ketat dengan basa 5' dari antikodon tRNA: misalnya, Y (mRNA) dapat berinteraksi dengan A dan G (tRNA)
Beberapa tRNA dapat berpasangan dengan lebih dari satu kodon.
63. Karakteristik unsur-unsur penyusun proses penerjemahan. Translasi (translatio-translation) adalah proses sintesis protein dari asam amino pada matriks informational (matriks) RNA (mRNA, mRNA) yang dilakukan oleh ribosom.
Sintesis protein adalah dasar dari kehidupan sel. Untuk melakukan proses ini dalam sel semua organisme ada organel khusus - ribosom- Kompleks ribonukleoprotein yang dibangun dari 2 subunit: besar dan kecil. Fungsi ribosom adalah untuk mengenali tiga huruf (tiga nukleotida) kodon mRNA, membandingkannya dengan antikodon tRNA yang sesuai yang membawa asam amino, dan penambahan asam amino ini ke rantai protein yang sedang tumbuh. Bergerak di sepanjang molekul mRNA, ribosom mensintesis protein sesuai dengan informasi yang terkandung dalam molekul mRNA.
Untuk pengenalan AK-t di dalam sel, ada "adaptor" khusus, mentransfer molekul RNA(tRNA). Molekul berbentuk daun semanggi ini memiliki situs (antikodon) yang melengkapi kodon mRNA, serta situs lain tempat asam amino yang sesuai dengan kodon tersebut terpasang. Pengikatan asam amino ke tRNA dilakukan dalam reaksi yang bergantung pada energi oleh enzim aminoasil-tRNA sintetase, dan molekul yang dihasilkan disebut aminoasil-tRNA. Dengan demikian, spesifisitas translasi ditentukan oleh interaksi antara kodon mRNA dan antikodon tRNA, serta spesifisitas sintetase aminoasil-tRNA yang mengikat asam amino secara ketat pada tRNA yang sesuai (misalnya, kodon GGU akan sesuai dengan a tRNA yang mengandung antikodon CCA, dan hanya AK glisin).
ribosom prokariotik
5S dan 23S rRNA 16S rRNA
34 tupai 21 tupai
Ribosom prokariotik memiliki konstanta sedimentasi 70S, itulah sebabnya mereka disebut partikel 70S. Mereka dibangun dari dua subunit yang berbeda: subunit 30S dan 50S. Setiap subunit adalah kompleks rRNA dan protein ribosom.
Partikel 30S mengandung satu molekul 16S rRNA dan dalam banyak kasus satu molekul protein dari lebih dari 20 spesies (21) . Subunit 50S terdiri dari dua molekul rRNA (23S dan 5S). Ini terdiri dari lebih dari 30 protein berbeda (34), juga diwakili, sebagai suatu peraturan, oleh satu salinan. Sebagian besar protein ribosom melakukan fungsi struktural.
ribosom eukariotik
5S; 5.8S dan 28S rRNA 18S rRNA
setidaknya 50 protein setidaknya 33 protein
Ribosom terdiri dari subunit besar dan kecil. Dasar dari struktur setiap subunit adalah rRNA yang terlipat secara kompleks. Protein ribosom melekat pada perancah rRNA.
Koefisien sedimentasi ribosom eukariotik lengkap adalah sekitar 80 unit Svedberg (80S), dan koefisien sedimentasi subpartikelnya adalah 40S dan 60S.
Subunit 40S yang lebih kecil terdiri dari satu molekul 18S rRNA dan 30-40 molekul protein. Subunit 60S besar berisi tiga jenis rRNA dengan koefisien sedimentasi protein 5S, 5.8S, dan 28S dan 40-50 (misalnya, ribosom hepatosit tikus termasuk 49 protein).
Daerah fungsional ribosom
P - situs peptidil untuk peptidil tRNA
A - situs aminoasil untuk aminoasil tRNA
E - situs untuk pelepasan tRNA dari ribosom
Ribosom mengandung 2 situs fungsional untuk interaksi dengan tRNA: aminoasil (akseptor) dan peptidil (donor). Aminoasil-tRNA memasuki situs akseptor ribosom dan berinteraksi untuk membentuk ikatan hidrogen antara triplet kodon dan antikodon. Setelah pembentukan ikatan hidrogen, sistem memajukan 1 kodon dan berakhir di situs donor. Pada saat yang sama, sebuah kodon baru muncul di situs akseptor yang dikosongkan, dan aminoasil-t-RNA yang sesuai melekat padanya.
Ribosom: struktur, fungsi
Ribosom adalah pusat sitoplasma biosintesis protein. Mereka terdiri dari subunit besar dan kecil, berbeda dalam koefisien sedimentasi (laju sedimentasi selama sentrifugasi), dinyatakan dalam satuan Svedberg - S.
Ribosom hadir dalam sel eukariotik dan prokariotik, karena mereka melakukan fungsi penting dalam: biosintesis protein. Di setiap sel ada puluhan, ratusan ribu (sampai beberapa juta) organel bulat kecil ini. Ini adalah partikel ribonukleoprotein bulat. Diameternya 20-30 nm. Ribosom terdiri dari subunit besar dan kecil, berbeda dalam koefisien sedimentasi (laju sedimentasi selama sentrifugasi), dinyatakan dalam unit Svedberg - S. Subunit ini digabungkan dengan adanya untaian m-RNA (matriks, atau informasional, RNA). Kompleks sekelompok ribosom yang disatukan oleh molekul mRNA tunggal seperti untaian manik-manik disebut polisom. Struktur-struktur ini terletak secara bebas di sitoplasma atau melekat pada membran RE granular (dalam kedua kasus, sintesis protein berlangsung secara aktif pada mereka).
Polisom ER granular membentuk protein yang dikeluarkan dari sel dan digunakan untuk kebutuhan seluruh organisme (misalnya, enzim pencernaan, protein ASI). Selain itu, ribosom hadir di permukaan bagian dalam membran mitokondria, di mana mereka juga mengambil bagian aktif dalam sintesis molekul protein.
Transfer RNA (tRNA) berperan penting dalam proses penggunaan informasi herediter oleh sel. Mengantarkan asam amino yang diperlukan ke tempat perakitan rantai peptida, tRNA bertindak sebagai mediator translasi.
Molekul tRNA adalah rantai polinukleotida yang disintesis pada sekuens DNA spesifik. Mereka terdiri dari sejumlah kecil nukleotida -75-95. Sebagai hasil dari hubungan komplementer basa yang terletak di berbagai bagian rantai polinukleotida tRNA, ia memperoleh struktur yang menyerupai bentuk daun semanggi (Gbr. 3.26).
Beras. 3.26. Struktur molekul tRNA yang khas.
Ini memiliki empat bagian utama yang melakukan fungsi yang berbeda. akseptor"Tangkai" dibentuk oleh dua bagian terminal terhubung yang saling melengkapi dari tRNA. Ini terdiri dari tujuh pasangan basa. Ujung 3' dari batang ini agak lebih panjang dan membentuk daerah untai tunggal yang diakhiri dengan rangkaian CCA dengan gugus OH bebas. Asam amino yang dapat diangkut melekat pada ujung ini. Tiga cabang yang tersisa adalah urutan nukleotida berpasangan komplementer yang berakhir pada bagian tidak berpasangan yang membentuk loop. Bagian tengah cabang ini - antikodon - terdiri dari lima pasang nukleotida dan mengandung antikodon di tengah lingkarannya. Antikodon adalah tiga nukleotida yang melengkapi kodon mRNA, yang mengkode asam amino yang diangkut oleh tRNA ini ke tempat sintesis peptida.
Antara cabang akseptor dan antikodon adalah dua cabang samping. Dalam loopnya, mengandung basa yang dimodifikasi - dihydrouridine (D-loop) dan triplet TψC, di mana \y adalah pseudouriain (T^C-loop).
Antara cabang aiticodone dan T^C ada loop tambahan, yang mencakup 3-5 hingga 13-21 nukleotida.
Secara umum, berbagai jenis tRNA dicirikan oleh keteguhan tertentu dari urutan nukleotida, yang paling sering terdiri dari 76 nukleotida. Variasi jumlah mereka terutama disebabkan oleh perubahan jumlah nukleotida dalam loop tambahan. Daerah komplementer yang mendukung struktur tRNA biasanya dilestarikan. Struktur primer tRNA, ditentukan oleh urutan nukleotida, membentuk struktur sekunder tRNA, yang berbentuk daun semanggi. Pada gilirannya, struktur sekunder menyebabkan struktur tersier tiga dimensi, yang dicirikan oleh pembentukan dua heliks ganda tegak lurus (Gbr. 3.27). Salah satunya dibentuk oleh cabang akseptor dan TψC, yang lain oleh antikodon dan cabang D.
Di ujung salah satu heliks ganda adalah asam amino yang diangkut, di ujung yang lain adalah antikodon. Daerah-daerah ini adalah yang paling terpencil satu sama lain. Stabilitas struktur tersier tRNA dipertahankan karena munculnya ikatan hidrogen tambahan antara basa rantai polinukleotida, yang terletak di bagian yang berbeda, tetapi secara spasial dekat dalam struktur tersier.
Berbagai jenis tRNA memiliki struktur tersier yang serupa, meskipun dengan beberapa variasi.
Beras. 3.27. Organisasi spasial tRNA:
I - struktur sekunder tRNA dalam bentuk "daun semanggi", ditentukan oleh struktur primernya (urutan nukleotida dalam rantai);
II - proyeksi dua dimensi dari struktur tersier tRNA;
III - tata letak molekul tRNA di luar angkasa
LAMPIRAN (jika ada yang tidak mengerti)
Gigi petir - nukleotida (Adenin-Timin / Urasil /, Guanin-Sitazin). Semua petir adalah DNA.

Untuk mentransfer informasi dari DNA, Anda perlu mematahkan 2 untai. Ikatan antara A-T dan G-C adalah hidrogen, oleh karena itu mudah diputus oleh enzim Helicase:

Untuk mencegah terbentuknya simpul (Sebagai contoh, saya memutar handuk):



Topoisomerase memotong satu untai DNA di tempat asal replikasi sehingga rantai tidak terpuntir.

Ketika satu utas bebas, yang kedua dapat dengan mudah berputar di sekitar porosnya, sehingga menghilangkan ketegangan selama "melepas". Node tidak muncul, energi dihemat.
Kemudian, primer RNA diperlukan untuk mulai mengumpulkan RNA. Protein yang merakit mRNA tidak bisa begitu saja merakit nukleotida pertama, ia membutuhkan sepotong RNA untuk memulai (tertulis secara rinci di sana, saya akan menulisnya nanti). Bagian ini disebut primer RNA. Dan protein ini sudah menempelkan nukleotida pertama padanya.


Ketika menggambarkan struktur asam nukleat, tingkat organisasi makromolekul yang berbeda diperhitungkan: struktur primer dan sekunder.
Struktur utama asam nukleat adalah komposisi nukleotida dan urutan unit nukleotida tertentu dalam rantai polimer.
Struktur sekunder RNA. Molekul asam ribonukleat dibangun dari rantai polinukleotida tunggal.
Struktur sekunder RNA
Bagian terpisah dari rantai RNA membentuk loop spiral - "jepit rambut", karena ikatan hidrogen antara basa nitrogen komplementer A-U dan G-C. Bagian rantai RNA dalam struktur heliks seperti itu antiparalel, tetapi tidak selalu saling melengkapi; mereka mengandung residu nukleotida yang tidak berpasangan atau bahkan loop beruntai tunggal yang tidak sesuai dengan heliks ganda. Kehadiran daerah spiral adalah karakteristik dari semua jenis RNA.
Peran utama RNA adalah partisipasi langsung dalam biosintesis protein.
Tiga jenis RNA seluler diketahui, yang berbeda dalam lokasinya di dalam sel, komposisi, ukuran dan sifat yang menentukan peran spesifiknya dalam pembentukan makromolekul protein:
- RNA informasi (matriks) mengirimkan informasi yang dikodekan dalam DNA tentang struktur protein dari inti sel ke ribosom, di mana sintesis protein dilakukan; struktur utama semua mRNA, terlepas dari keunikan urutan pengkodeannya, memiliki struktur ujung 5'- dan 3' yang sama.
Jadi, pada ujung 5' terdapat nukleotida 7-metilguanosin-5'-trifosfat yang dimodifikasi (tutup). Beberapa puluh nukleotida memisahkan tutup dari kodon inisiasi, biasanya triplet -AUG-. Daerah pengkode diikuti oleh salah satu kodon terminasi -UGA-, -UUA-, -UAG-. Pada ujung 3' sebagian besar mRNA, terdapat urutan nukleotida dari 100-200 residu adenosin monofosfat.
- RNA transfer mengumpulkan asam amino dalam sitoplasma sel dan mentransfernya ke ribosom; Molekul RNA jenis ini "belajar" dari bagian yang sesuai dari rantai RNA pembawa pesan yang asam amino harus berpartisipasi dalam sintesis protein.
Struktur spasial tRNA apa pun, terlepas dari perbedaan urutan nukleotida, dijelaskan oleh model daun semanggi universal. Setiap molekul tRNA memiliki bagian rantai yang tidak terlibat dalam pembentukan ikatan hidrogen antara residu nukleotida.
Ini termasuk, khususnya, situs yang bertanggung jawab untuk mengikat asam amino pada ujung 3' molekul dan antikodon, triplet nukleotida spesifik yang berinteraksi komplementer dengan kodon mRNA.
- RNA ribosom menyediakan sintesis protein dari struktur tertentu, membaca informasi dari informasi (matriks) RNA. rRNA membentuk kompleks dengan protein yang disebut ribosom.
Setiap ribosom terdiri dari dua subunit - kecil (40S) dan besar (60S). Subunit ribosom berbeda tidak hanya dalam set rRNA, tetapi juga dalam jumlah dan struktur protein.
Tanggal publikasi: 02-03-2015; Baca: 2729 | Pelanggaran hak cipta halaman

RNA adalah polimer yang monomernya adalah ribonukleotida.
Tidak seperti DNA, RNA dibentuk bukan oleh dua, tetapi oleh satu rantai polinukleotida (pengecualian - beberapa virus yang mengandung RNA memiliki RNA untai ganda). Nukleotida RNA mampu membentuk ikatan hidrogen satu sama lain. Rantai RNA jauh lebih pendek daripada rantai DNA.
Monomer RNA - nukleotida (ribonukleotida) - terdiri dari residu tiga zat: 1) basa nitrogen, 2) monosakarida lima karbon (pentosa) dan 3) asam fosfat. Basa nitrogen RNA juga termasuk dalam kelas pirimidin dan purin.
Basa pirimidin RNA - urasil, sitosin, basa purin - adenin dan guanin.
31. Jenis RNA dan fitur strukturnya
Monosakarida nukleotida RNA diwakili oleh ribosa.
Ada tiga jenis RNA: 1) informasi (matriks) RNA - mRNA (mRNA), 2) RNA transfer - tRNA, 3) RNA ribosom - rRNA.
Semua jenis RNA adalah polinukleotida tidak bercabang, memiliki konformasi spasial tertentu dan mengambil bagian dalam proses sintesis protein.
Informasi tentang struktur semua jenis RNA disimpan dalam DNA. Proses sintesis RNA pada cetakan DNA biasa disebut transkripsi.

RNA transfer biasanya mengandung 76 (dari 75 hingga 95) nukleotida; berat molekul - 25.000–30.000.
tRNA menyumbang sekitar 10% dari total konten RNA dalam sel. Fungsi tRNA: 1) transpor asam amino ke tempat sintesis protein, ke ribosom, 2) mediator translasi. Sekitar 40 jenis tRNA ditemukan di dalam sel, masing-masing memiliki karakteristik urutan nukleotida hanya untuk itu. Pada saat yang sama, semua tRNA memiliki beberapa daerah komplementer intramolekul, karena itu tRNA memperoleh konformasi yang menyerupai bentuk daun semanggi.
Setiap tRNA memiliki loop untuk kontak dengan ribosom (1), loop antikodon (2), loop untuk kontak dengan enzim (3), batang akseptor (4), dan antikodon (5). Asam amino melekat pada ujung 3' batang akseptor. Antikodon - tiga nukleotida yang "mengenali" kodon mRNA.
Harus ditekankan bahwa tRNA tertentu dapat mengangkut asam amino yang ditentukan secara ketat sesuai dengan antikodonnya. Kekhususan koneksi asam amino dan tRNA dicapai karena sifat-sifat enzim aminoasil-tRNA sintetase.
RNA ribosom mengandung 3000–5000 nukleotida; berat molekul - 1.000.000-1.500.000.
rRNA menyumbang 80-85% dari total konten RNA dalam sel. Dalam kompleks dengan protein ribosom, rRNA membentuk ribosom - organel yang melakukan sintesis protein. Pada sel eukariotik, sintesis rRNA terjadi di nukleolus. Fungsi rRNA: 1) komponen struktural penting dari ribosom dan, dengan demikian, memastikan fungsi ribosom; 2) memastikan interaksi ribosom dan tRNA; 3) pengikatan awal ribosom dan kodon inisiator mRNA dan penentuan kerangka baca, 4) pembentukan pusat aktif ribosom.
Messenger RNA memiliki kandungan nukleotida dan berat molekul yang beragam (dari 50.000 hingga 4.000.000).
Bagian mRNA menyumbang hingga 5% dari total konten RNA dalam sel. Fungsi mRNA: 1) transfer informasi genetik dari DNA ke ribosom, 2) matriks untuk sintesis molekul protein, 3) penentuan urutan asam amino dari struktur primer molekul protein.
Baca juga
RNA adalah polimer yang monomernya adalah nukleotida.
Tiga basa nitrogen sama seperti pada DNA (adenin, guanin, sitosin); keempat - urasil - hadir dalam molekul RNA bukan timin. Nukleotida RNA mengandung ribosa, bukan deoksiribosa. Dalam rantai RNA...
tiga jenis utama RNA: informasional(mRNA), atau matriks(mRNA), ribosom(rRNA), dan mengangkut(tRNA). Mereka berbeda dalam ukuran molekul dan fungsi. Semua jenis RNA disintesis pada DNA dengan partisipasi enzim - RNA polimerase. Messenger RNA membentuk 2-3% dari semua RNA seluler, ribosom - 80-85, transportasi - sekitar 15%.
mRNA.
ia membaca informasi herediter dari segmen DNA dan, dalam bentuk salinan urutan basa nitrogen, mentransfernya ke ribosom, tempat protein tertentu disintesis. Setiap molekul mRNA dalam urutan nukleotida dan ukurannya sesuai dengan gen dalam DNA dari mana ia ditranskripsi. Rata-rata, mRNA mengandung 1500 nukleotida (75-3000). Setiap triplet (tiga nukleotida) pada mRNA disebut kodon. Itu tergantung pada kodon asam amino mana yang akan muncul di tempat tertentu selama sintesis protein.
(tRNA) memiliki berat molekul yang relatif rendah sekitar 24-29 ribu.
D dan mengandung 75 hingga 90 nukleotida dalam molekulnya. Hingga 10% dari semua nukleotida tRNA adalah basa minor, yang tampaknya melindunginya dari aksi enzim hidrolitik.Peran tRNA adalah mentransfer asam amino ke ribosom dan berpartisipasi dalam proses sintesis protein. Setiap asam amino menempel pada tRNA tertentu. Sejumlah asam amino memiliki lebih dari satu tRNA. Sampai saat ini, lebih dari 60 tRNA telah ditemukan yang berbeda dalam struktur primernya (urutan basa).
Struktur sekunder semua tRNA disajikan dalam bentuk daun semanggi dengan batang beruntai ganda dan tiga beruntai tunggal). Di ujung salah satu rantai ada situs akseptor - triplet CCA, ke adenin di mana asam amino spesifik terpasang.
(rRNA). Mereka mengandung 120-3100 nukleotida. RNA ribosom terakumulasi dalam nukleus, di nukleolus.
Protein ribosom diangkut ke nukleolus dari sitoplasma, dan pembentukan spontan subpartikel ribosom terjadi di sana dengan menggabungkan protein dengan rRNA yang sesuai. Subpartikel ribosom diangkut bersama-sama atau secara terpisah melalui pori-pori membran nukleus ke dalam sitoplasma. Ribosom adalah organel berukuran 20-30 nm.
Mereka dibangun dari dua subpartikel dengan ukuran dan bentuk yang berbeda. Pada tahap tertentu sintesis protein dalam sel, ribosom dibagi menjadi subpartikel.
RNA ribosom berfungsi sebagai kerangka kerja untuk ribosom dan memfasilitasi pengikatan awal mRNA ke ribosom selama biosintesis protein.
Soal 6 Ikatan yang membentuk struktur primer dan sekunder DNA dan RNA. Jenis RNA
Kode genetik adalah cara pengkodean urutan asam amino protein menggunakan urutan nukleotida, karakteristik semua organisme hidup.
Properti: 1) kode genetik tiga serangkai(setiap asam amino dikodekan oleh tiga nukleotida); 2) tidak tumpang tindih(kembar tiga tetangga tidak memiliki nukleotida yang sama); 3) merosot(dengan pengecualian metionin dan triptofan, semua asam amino memiliki lebih dari satu kodon); 4) universal(kebanyakan sama untuk semua organisme hidup); 5) dalam kodon untuk satu asam amino, dua nukleotida pertama biasanya sama, dan yang ketiga bervariasi; 6) memiliki urutan bacaan linier dan dicirikan oleh kolinearitas, t.
e.kesesuaian urutan kodon pada mRNA dengan urutan asam amino pada rantai polipeptida yang disintesis.
Tanggal publikasi: 2014-12-08; Baca: 11268 | Pelanggaran hak cipta halaman
studopedia.org - Studopedia.Org - 2014-2018 (0,001 dtk) ...
Sitoplasma sel mengandung tiga jenis fungsional utama RNA:
- messenger RNA (mRNA) yang bertindak sebagai cetakan untuk sintesis protein;
- RNA ribosom (rRNA) bertindak sebagai komponen struktural ribosom;
- mentransfer RNA (tRNA) yang terlibat dalam translasi (translasi) informasi mRNA ke dalam urutan asam amino dari molekul protein.
Dalam nukleus sel, RNA inti ditemukan, yang terdiri dari 4 hingga 10% dari total RNA seluler.
Sebagian besar RNA nuklir diwakili oleh prekursor molekul tinggi dari RNA ribosom dan transfer. Prekursor rRNA dengan berat molekul tinggi (28 S, 18 S dan 5 S RNA) terutama terlokalisasi di nukleolus.
RNA adalah materi genetik utama pada beberapa virus hewan dan tumbuhan (RNA genomik). Sebagian besar virus RNA dicirikan oleh transkripsi balik genom RNA mereka, yang diarahkan oleh reverse transcriptase.
Semua asam ribonukleat adalah polimer dari ribonukleotida yang terhubung, seperti dalam molekul DNA, dengan ikatan 3′,5′-fosforodiester.
Tidak seperti DNA, yang memiliki struktur untai ganda, RNA adalah molekul polimer linier beruntai tunggal.
struktur mRNA. mRNA adalah kelas RNA yang paling heterogen dalam hal ukuran dan stabilitas.
struktur tRNA.
Transfer RNA bertindak sebagai mediator (adaptor) selama translasi mRNA. Mereka menyumbang sekitar 15% dari total RNA seluler. Masing-masing dari 20 asam amino proteinogenik memiliki tRNA sendiri. Untuk beberapa asam amino yang dikodekan oleh dua atau lebih kodon, ada beberapa tRNA.
tRNA adalah molekul untai tunggal yang relatif kecil yang terdiri dari 70-93 nukleotida. Berat molekulnya adalah (2.4-3.1) .104 kDa.
Struktur sekunder tRNA terbentuk karena pembentukan jumlah maksimum ikatan hidrogen antara pasangan basa nitrogen komplementer intramolekul.
Sebagai hasil dari pembentukan ikatan ini, rantai polinukleotida tRNA berputar dengan pembentukan cabang spiral yang berakhir pada loop nukleotida yang tidak berpasangan. Gambar spasial struktur sekunder semua tRNA memiliki bentuk daun semanggi.
Empat cabang wajib dibedakan dalam "daun semanggi", tRNA yang lebih panjang, di samping itu, mengandung cabang kelima (tambahan) yang pendek.
Fungsi adaptor tRNA disediakan oleh cabang akseptor, pada ujung 3' di mana residu asam amino dilekatkan oleh ikatan eter, dan cabang antikodon di seberang cabang akseptor, di atasnya terdapat loop yang mengandung sebuah antikodon.
Antikodon adalah triplet nukleotida spesifik yang komplementer dalam arah antiparalel dengan kodon mRNA yang mengkode asam amino yang sesuai.
Cabang-T yang membawa loop pseudouridine (TyC-loop) memastikan interaksi tRNA dengan ribosom.
Cabang D, membawa loop dehydrouridine, memastikan interaksi tRNA dengan sintetase aminoasil-tRNA yang sesuai.

Struktur sekunder tRNA
Fungsi cabang tambahan kelima masih kurang dipahami; kemungkinan besar, itu menyamakan panjang molekul tRNA yang berbeda.
Struktur tersier tRNA sangat kompak dan dibentuk dengan menyatukan cabang individu daun semanggi karena ikatan hidrogen tambahan untuk membentuk struktur "siku" berbentuk L.
Transport RNA, struktur dan mekanisme fungsional.
Dalam hal ini, lengan akseptor yang mengikat asam amino terletak di salah satu ujung molekul, dan anti-kodon di ujung lainnya.

Struktur tersier tRNA (menurut A.S. Spirin)
Struktur rRNA dan ribosom. RNA ribosom membentuk tulang punggung yang mengikat protein spesifik untuk membentuk ribosom. Ribosom adalah organel nukleoprotein yang menyediakan sintesis protein dari mRNA.
Jumlah ribosom dalam sel sangat besar: dari 104 pada prokariota hingga 106 pada eukariota. Ribosom terlokalisasi terutama di sitoplasma, di eukariota, di samping itu, di nukleolus, di matriks mitokondria dan di stroma kloroplas. Ribosom terdiri dari dua subpartikel: besar dan kecil. Berdasarkan ukuran dan berat molekul, semua ribosom yang dipelajari dibagi menjadi 3 kelompok - ribosom 70S prokariota (koefisien sedimentasi S), yang terdiri dari subpartikel kecil 30S dan 50S besar; Ribosom eukariotik 80S, terdiri dari subunit kecil 40S dan subunit besar 60S.
Subunit kecil ribosom 80S dibentuk oleh satu molekul rRNA (18S) dan 33 molekul berbagai protein.
Subunit besar dibentuk oleh tiga molekul rRNA (5S, 5.8S, dan 28S) dan sekitar 50 protein.
Struktur sekunder rRNA terbentuk karena bagian molekul beruntai ganda pendek - jepit rambut (sekitar 2/3 dari rRNA), 1/3 - diwakili oleh bagian beruntai tunggal yang kaya akan nukleotida purin.
Tombol sosial untuk Joomla
Protein membentuk dasar kehidupan. Fungsi mereka di dalam sel sangat beragam. Namun, protein "tidak bisa" bereproduksi. Dan semua informasi tentang struktur protein terkandung dalam gen (DNA).
Pada organisme yang lebih tinggi, protein disintesis dalam sitoplasma sel, dan DNA tersembunyi di balik cangkang nukleus. Oleh karena itu, DNA tidak dapat langsung berfungsi sebagai cetakan untuk sintesis protein. Peran ini dilakukan oleh asam nukleat lain - RNA.
Molekul RNA adalah polinukleotida tidak bercabang dengan struktur tersier.
Ini dibentuk oleh satu rantai polinukleotida, dan meskipun nukleotida komplementer yang termasuk di dalamnya juga mampu membentuk ikatan hidrogen di antara mereka, ikatan ini terjadi antara nukleotida dari satu rantai. Rantai RNA jauh lebih pendek daripada rantai DNA. Jika kandungan DNA dalam sel relatif konstan, maka kandungan RNA sangat berfluktuasi. Jumlah terbesar RNA dalam sel diamati selama sintesis protein.
RNA memainkan peran utama dalam transmisi dan implementasi informasi herediter.
Sesuai dengan fungsi dan fitur struktural, beberapa kelas RNA seluler dibedakan.
Ada tiga kelas utama RNA seluler.
- Informasional (mRNA), atau matriks (mRNA). Molekulnya paling beragam dalam hal ukuran, berat molekul (dari 0,05x106 hingga 4x106) dan stabilitas.
Mereka membuat sekitar 2% dari jumlah total RNA dalam sel. Semua mRNA adalah pembawa informasi genetik dari nukleus ke sitoplasma, ke tempat sintesis protein. Mereka berfungsi sebagai matriks (gambar kerja) untuk sintesis molekul protein, karena mereka menentukan urutan asam amino (struktur utama) dari molekul protein.

- RNA ribosom (rRNA).
Mereka membuat 80-85% dari total konten RNA dalam sel.
31. Struktur RNA. Jenis RNA, fitur struktural dan fungsi. Struktur sekunder tRNA
RNA ribosom terdiri dari 3-5 ribu nukleotida. Ini disintesis dalam nukleolus nukleus. Dalam kompleks dengan protein ribosom, rRNA membentuk ribosom - organel tempat molekul protein dirakit. Signifikansi utama rRNA adalah menyediakan pengikatan awal mRNA dan ribosom dan membentuk pusat aktif ribosom, di mana ikatan peptida terbentuk antara asam amino selama sintesis rantai polipeptida.
- Transfer RNA (tRNA).
Molekul tRNA biasanya mengandung 75-86 nukleotida. Berat molekul molekul tRNA adalah sekitar 25 ribu. Molekul tRNA memainkan peran perantara dalam biosintesis protein - mereka mengirimkan asam amino ke tempat sintesis protein, yaitu ke ribosom. Sel mengandung lebih dari 30 jenis tRNA. Setiap jenis tRNA memiliki urutan nukleotida yang unik.
Namun, semua molekul memiliki beberapa daerah komplementer intramolekul, karena kehadirannya semua tRNA memiliki struktur tersier yang menyerupai bentuk daun semanggi.
Struktur sekunder RNA- ciri tRNA, beruntai tunggal, berbentuk seperti "daun semanggi".
Termasuk:
- heliks ganda yang relatif pendek - batang,
- bagian beruntai tunggal - loop.
Ada 4 batang (akseptor, antikodon, dihydrouridyl, pseudouridyl) dan 3 loop.

"Stem-loop" - elemen struktur sekunder RNA, secara skematis

"Pseudoknot" - elemen struktur sekunder RNA, secara skematis

Batang akseptor mengandung ujung 3'- dan 5' dari rantai polinukleotida, ujung 5' berakhir dengan residu asam guanylic, ujung 3' adalah triplet CCA dan berfungsi untuk membentuk ikatan ester dengan AA.
Batang antikodon mengenali kodonnya pada mRNA di ribosom dengan prinsip saling melengkapi.
Batang pseudouridyl berfungsi untuk menempel pada ribosom.
Batang dihydrouridyl berfungsi untuk mengikat sintetase aminoasil-tRNA.
Tombol sosial untuk Joomla
Kami juga merekomendasikan
 Cara membuat smoothie pisang yang sehat
Cara membuat smoothie pisang yang sehat
 Memanen asparagus untuk resep musim dingin untuk memasak di rumah
Memanen asparagus untuk resep musim dingin untuk memasak di rumah
 Pai ayam dengan zucchini dan keju cottage Resep Dukan pai zucchini dengan keju cottage
Pai ayam dengan zucchini dan keju cottage Resep Dukan pai zucchini dengan keju cottage
 Roti jahe dengan icing
Roti jahe dengan icing
 Cara memasak salad dengan tongkat kepiting dan wortel
Cara memasak salad dengan tongkat kepiting dan wortel
 Salad kubis dengan paprika - resep terbaik
Salad kubis dengan paprika - resep terbaik