Domača naloga. Infuzorija copat
V primerjavi z drugimi skupinami protozojev imajo ciliati najbolj zapleteno strukturo, kar je povezano z raznolikostjo in kompleksnostjo njihovih funkcij.
Od kod ime "infuzoria copat"? Ne boste presenečeni, če boste pod mikroskopom pogledali živo trepalnico ali celo njeno podobo (slika 85).
Dejansko oblika telesa tega ciliata spominja na eleganten ženski čevelj.
Čevelj za infuzorijo je v neprekinjenem precej hitrem gibanju. Njegova hitrost (pri sobni temperaturi) je približno 2,0-2,5 mm/s. To je velika hitrost za tako majhno žival! Konec koncev to pomeni, da čevelj v sekundi preteče razdaljo, ki za 10-15 krat presega dolžino njegovega telesa. Pot čevlja je precej zapletena. Premakne sprednji del naravnost naprej
INFUSORIJ ČEVELJ (PARAMECIUM CAUDATUM)
Da bi se seznanili z zgradbo in načinom življenja teh zanimivih enoceličnih organizmov, se najprej obrnimo na en značilen primer. Vzemimo ciliate (vrste iz rodu Paramecium), ki so razširjene v plitvih sladkovodnih rezervoarjih. Te ciliate je zelo enostavno vzrejati v majhnih akvarijih, če ribnik napolnite z navadnim travniškim senom. V takih tinkturah se razvijejo številne različne vrste protozojev, skoraj vedno pa se razvijejo čevlji. S pomočjo navadnega izobraževalnega mikroskopa si lahko ogledate marsikaj, o čemer bomo še govorili.
Med najpreprostejšimi ciliates čevlji so dokaj veliki organizmi. Njihova dolžina telesa je približno 1/6-1/3 mm. in se vrti v desno vzdolž vzdolžne osi telesa.
Tako aktivno gibanje čevlja je odvisno od dela velikega števila najfinejših lasu podobnih dodatkov - ciliatov, ki pokrivajo celotno telo trepalk. Število cilijev v enem posamezniku ciliatov čevljev je 10-15 tisoč!
Vsaka cilija naredi zelo pogoste gibe v obliki vesla - pri sobni temperaturi do 30 utripov na sekundo. Med udarcem nazaj se cilij drži v zravnanem položaju. Ko se vrne v prvotni položaj (pri premikanju navzdol), se premika 3-5 krat počasneje in opisuje polkrog.
Ko čevelj plava, se povzamejo gibi številnih cilijev, ki pokrivajo njegovo telo. Delovanje posameznih cilij je usklajeno, kar ima za posledico pravilne valovite vibracije vseh cilij. Nihajni val se začne na sprednjem koncu telesa in se širi nazaj. Hkrati potekajo 2-3 valovi krčenja vzdolž telesa čevlja. Tako je celoten ciliarni aparat ciliata tako rekoč ena sama funkcionalna fiziološka celota, katere dejanja posameznih strukturnih enot (cilije) so med seboj tesno povezana (usklajena).
Struktura vsakega posameznega cilija čevlja, kot kažejo elektronske mikroskopske študije, je zelo zapletena.
Smer in hitrost gibanja čevlja nista stalni in nespremenljivi količini. Čevelj, tako kot vsi živi organizmi (to smo že videli na primeru amebe), reagira na spremembo zunanje okolje sprememba smeri gibanja.
Sprememba smeri gibanja protozojev pod vplivom različnih dražljajev se imenuje taksi. Pri ciliatih je enostavno opazovati različne taksije. Če v kapljico, kjer plavajo čevlji, položite kakšno snov, ki nanje negativno vpliva (na primer kristal namizna sol), potem čevlji odplavajo (kot da bežijo) od tega zanje neugodnega dejavnika (slika 86).

Pred nami je primer negativne taksije na kemični učinek (negativna kemotaksija). Lahko opazujete čevelj in pozitivno kemotaksijo. Če na primer kapljico vode, v kateri plavajo ciliati, pokrijemo s pokrivnim steklom in pod njo spustimo mehurček ogljikovega dioksida (CO2), potem bo večina ciliatov odšla do tega mehurčka in se okoli njega naselila v prstan.
Fenomen taksijev se zelo jasno kaže v čevljih pod vplivom električnega toka. Če skozi tekočino, v kateri plavajo čevlji, preide šibek električni tok, lahko opazimo naslednjo sliko: vsi ciliati usmerijo svojo vzdolžno os vzporedno s trenutno črto in se nato, kot na ukaz, premaknejo proti katodi, na območju katerega tvorijo gosto gručo. Gibanje ciliatov, ki ga določa smer električnega toka, se imenuje galvanotaksija. Pod vplivom najrazličnejših okoljskih dejavnikov je mogoče zaznati različne taksije pri ciliatih.
Celotno citoplazmatsko telo ciliata je jasno razdeljeno na 2 plasti: zunanja je svetlejša (ektoplazma) in notranja je temnejša in zrnata (endoplazma).Pomembna vloga pri ohranjanju konstantnosti oblike telesa trepalk.
V zunanji plasti (v ektoplazmi) telesa živega čevlja so jasno vidne številne kratke palice, ki se nahajajo pravokotno na površino (sl. 85, 7). Te formacije se imenujejo trihociste. Njihova funkcija je zelo zanimiva in je povezana z zaščito najpreprostejših. Ob mehanskem, kemičnem ali kakšnem drugem močnem draženju se trihociste s silo vržejo ven in se spremenijo v tanke dolge niti, ki zadenejo plenilca, ki napada čevelj. Trichocyste so močna obramba. Redno so razporejene med cilije, tako da število trihocist približno ustreza številu cilij. Namesto uporabljenih (»streljenih«) trihocist se v ektoplazmi čevlja razvijejo nove.
Na eni strani, približno na sredini telesa (sl. 85, 5), ima čevelj precej globoko vdolbino. To je ustna votlina ali peristom. Cilije se nahajajo vzdolž sten peristoma, pa tudi vzdolž površine telesa. Tu so razvite veliko močneje kot na preostali površini telesa. Te tesno razporejene cilije so razporejene v dve skupini. Funkcija teh visoko diferenciranih cilij ni povezana z gibanjem, temveč s prehrano (slika 87).

Kako in kaj jedo čevlji, kako se prebavijo?
Čevlji sodijo med ciliate, katerih glavna hrana so bakterije. Poleg bakterij lahko pogoltnejo tudi vse druge delce, suspendirane v vodi, ne glede na njihovo hranilno vrednost. Peroralne cilije ustvarjajo neprekinjen tok vode z suspendiranimi delci v smeri ustne odprtine, ki se nahaja globoko v peristomu. Majhni delci hrane (najpogosteje bakterije) prodrejo skozi usta v majhen cevasti žrelo in se kopičijo na njegovem dnu, na meji z endoplazmo. Odprtina ust je vedno odprta. Morda ne bo napačno reči, da je ciliatni čevelj ena najbolj požrešnih živali: nenehno se hrani. Ta proces se prekine le v določenih trenutkih življenja, povezanih z razmnoževanjem in spolnim procesom.
Gruda hrane, nakopičena na dnu žrela, se nato odcepi od dna žrela in skupaj z majhno količino tekočine vstopi v endoplazmo in tvori prebavno vakuolo. Slednji ne ostane na mestu nastanka, ampak, ko pade v tokove endoplazme, naredi precej zapleteno in pravilno pot v telesu čevlja, imenovano cikloza prebavne vakuole (slika 88). Med tem precej dolgim (pri sobni temperaturi, ki traja približno eno uro) potovanjem prebavne vakuole se v njej zgodijo številne spremembe, povezane s prebavo hrane v njej.
Tu, tako kot pri amebah in nekaterih flagelatih, poteka tipična znotrajcelična prebava. Iz endoplazme, ki obdaja prebavno vakuolo, vanjo vstopajo prebavni encimi, ki delujejo na delce hrane. Prebavni produkti se absorbirajo skozi prebavno vakuolo v endoplazmo.
Med ciklozo prebavne vakuole se v njej zamenja več faz prebave. V prvih trenutkih po nastanku vakuole se tekočina, ki jo polni, malo razlikuje od tekočine iz okolja. Kmalu začnejo prebavni encimi vstopati v vakuolo iz endoplazme in reakcija okolja v njej postane močno kisla. To je enostavno zaznati z dodajanjem nekega indikatorja živilu, katerega barva se spreminja glede na reakcijo (kisla, nevtralna ali alkalna) okolja. V tem kislem okolju potekajo prve faze prebave. Nato se slika spremeni in reakcija v prebavnih vakuolah postane rahlo alkalna. Pod temi pogoji obstajajo Naslednji koraki znotrajcelična prebava. Kisla faza je običajno krajša od alkalne faze; traja približno 1/6-1/4 celotnega bivanja prebavne vakuole v telesu ciliata. Razmerje kisle in alkalne faze pa se lahko razlikuje v precej širokih mejah, odvisno od narave hrane.
Pot prebavne vakuole v endoplazmi se konča z dejstvom, da se približa površini telesa in skozi pelikulo se njegova vsebina, sestavljena iz tekočih in neprebavljenih ostankov hrane, vrže ven - pride do iztrebljanja. Ta proces, za razliko od ameb, pri katerih se iztrebljanje lahko pojavi kjer koli, je v čevljih, tako kot pri drugih ciliatih, strogo omejen na določeno področje telesa, ki se nahaja na ventralni strani (trebuh se običajno imenuje površina živali na kateri je nameščena perioralna vdolbina), približno na sredini med peristomom in zadnjim koncem telesa.
Tako je znotrajcelična prebava zapleten proces, sestavljen iz več faz, ki se zaporedoma zamenjajo.

Izračuni kažejo, da se v približno 30-45 minutah iz čevlja skozi kontraktilne vakuole izloči prostornina tekočine, ki je enaka volumnu ciliatnega telesa. Tako se zaradi delovanja kontraktilnih vakuol skozi telo ciliata izvaja neprekinjen tok vode, ki vstopa od zunaj skozi ustno odprtino (skupaj s prebavnimi vakuolami), pa tudi osmotsko neposredno skozi pelikulo. Kontraktilne vakuole igrajo pomembno vlogo pri uravnavanju pretoka vode, ki prehaja skozi telo ciliata, in pri uravnavanju osmotskega tlaka. Ta proces tukaj poteka načeloma na enak način kot pri amebah, le struktura kontraktilne vakuole je veliko bolj zapletena.
Med znanstveniki, ki so preučevali protozoje, je dolga leta obstajal spor o vprašanju, ali so v citoplazmi kakršne koli strukture, povezane s pojavom kontraktilne vakuole, ali se ta tvori vsakič znova. Na živem ciliatu ni mogoče opaziti posebnih struktur, ki bi bile pred nastankom. Ko pride do krčenja vakuole - sistole - v citoplazmi na mestu nekdanje vakuole niso vidne popolnoma nobene strukture. Nato se ponovno pojavijo prozorni mehurčki ali adduktorski kanali, ki se začnejo povečevati. Vendar pa ni mogoče najti povezave med novo nastalo vakuolo in prej obstoječo vakuolo. Zdi se, da ni kontinuitete med zaporednimi cikli kontraktilne vakuole in vsaka nova kontraktilna vakuola se v citoplazmi tvori na novo. Vendar so posebne raziskovalne metode pokazale, da v resnici ni tako. Uporaba elektronske mikroskopije, ki daje zelo veliko povečavo (do 100 tisoč-krat), je prepričljivo pokazala, da ima ciliat posebej diferencirano citoplazmo na območju, kjer nastajajo kontraktilne vakuole, sestavljene iz prepletanja najtanjših cevk. Tako se je izkazalo, da kontraktilna vakuola ne nastane v citoplazmi na " prazno mesto«, ampak na podlagi prejšnjega posebnega celičnega organoida, katerega funkcija je tvorba kontraktilne vakuole.
Kot vsi protozoji imajo ciliati celično jedro. Vendar pa se po strukturi jedrskega aparata ciliati močno razlikujejo od vseh drugih skupin protozojev.
Za jedrski aparat ciliatov je značilen dualizem. To pomeni, da imajo ciliati dve različni vrsti jeder - velika jedra ali makronukleusi in majhna jedra ali mikronukleusa. Poglejmo, kakšno strukturo ima jedrski aparat v ciliatih čevlja (slika 85).
V središču telesa ciliata (na ravni peristoma) je nameščeno veliko masivno jedro jajčaste ali fižolaste oblike. To je makronukleus. V neposredni bližini nje je drugo jedro, večkrat manjše, običajno precej blizu makronukleusa. To je mikronukleus. Razlika med tema dvema jedroma ni le v velikosti, je pomembnejša in močno vpliva na njihovo strukturo.
Makronukleus je v primerjavi z mikronukleusom veliko bogatejši s posebno jedrsko snovjo (kromatin ali, natančneje, deoksiribonukleinska kislina, skrajšano DNK), ki je del kromosomov.
Nedavne študije so pokazale, da ima makronukleus več deset (in pri nekaterih ciliatih in stotine) krat veliko število kromosomov kot mikronukleusov. Makronukleus je zelo svojevrstna vrsta multikromosomskih (poliploidnih) jeder. Tako razlika med mikro- in makronukleusi vpliva na njihovo kromosomsko sestavo, ki določa večje ali manjše bogastvo njihove jedrske snovi – kromatina.
V eni najpogostejših vrst ciliatov - čevlji(Paramecium caudatum) - ima eno makronukleus (skrajšano Ma) in eno mikronukleus (skrajšano Mi). Ta struktura jedrskega aparata je značilna za številne ciliate. Drugi imajo lahko več Ma in Mi. Toda značilna lastnost vseh ciliatov je diferenciacija jeder v dve kvalitativno različni skupini, v Ma in Mi, ali, z drugimi besedami, fenomen jedrskega dualizma.
Kako se ciliati razmnožujejo? Za primer se spet obrnimo na čevelj iz infuzorije. Če posadite en izvod čevlja v majhno posodo (mikroakvarij), potem bosta čez dan dva, pogosto pa štirje ciliati. Kako se to zgodi? Po določenem obdobju aktivnega plavanja in hranjenja se ciliat nekoliko raztegne v dolžino. Nato se točno na sredini telesa pojavi vedno globlja prečna zožitev (slika 90). Na koncu so ciliati tako rekoč prepleteni na polovico in iz enega posameznika dobimo dva posameznika, sprva nekoliko manjša od matičnega posameznika. Celoten proces cepitve traja približno eno uro pri sobni temperaturi. Študija notranjih procesov kaže, da se še preden se pojavi prečna zožitev, začne proces cepitve jedrskega aparata. Najprej se deli Mi in šele nato Ma. Tukaj se ne bomo ustavili podroben premislek procese jedrske cepitve in opazimo le, da se Mi deli z mitozo, medtem ko delitev Ma po videzu spominja na neposredno jedrsko cepitev - amitozo. Ta proces nespolnega razmnoževanja ciliatov čevljev je, kot vidimo, podoben nespolnemu razmnoževanju ameb in flagelatov. Nasprotno pa se ciliati v procesu nespolnega razmnoževanja vedno delijo počez, medtem ko je pri bičastih ravnina delitve vzporedna z vzdolžno osjo telesa.
Med delitvijo pride do globokega notranjega prestrukturiranja ciliatnega telesa. Nastaneta dva nova peristoma, dva žrela in dve ustni odprtini. Hkrati je časovno določena delitev bazalnih jeder cilij, zaradi česar nastanejo nove cilije. Če se število cilij med razmnoževanjem ne bi povečalo, bi hčerinski posamezniki zaradi vsake delitve prejeli približno polovico števila cilijev materinega posameznika, kar bi privedlo do popolne "plešavosti" trepalk. Pravzaprav se to ne zgodi.

Od časa do časa ima večina ciliatov, vključno s čevlji, posebno in izjemno nenavadno obliko spolnega procesa, ki se imenuje konjugacija. Tukaj ne bomo podrobno analizirali vseh kompleksnih jedrskih sprememb, ki spremljajo ta proces, ampak upoštevamo le najpomembnejše. Konjugacija poteka na naslednji način (slika 91), dve ciliatu se približata drug drugemu, tesno pritrjena drug na drugega s trebušnimi stranicami in v tej obliki precej plavata. dolgo časa skupaj (pri čevlju približno 12 ur pri sobni temperaturi). Konjuganti se nato ločijo. Kaj se zgodi v telesu ciliatov med konjugacijo? Bistvo teh procesov je naslednje (slika 91). Veliko jedro (makronukleus) se zruši in postopoma raztopi v citoplazmi. Mikronukleusi se najprej razdelijo, nekatera jedra, ki nastanejo kot posledica cepitve, so uničena (glej sliko 91). Vsak od konjugantov zadrži dve jedri. Eno od teh jeder ostane na mestu v posamezniku, v katerem je nastalo (stacionarno jedro), medtem ko se drugo aktivno premakne v konjugacijskega partnerja (migrirajoče jedro) in se zlije s svojim stacionarnim jedrom. Tako je v vsakem od konjugantov na tej stopnji eno jedro, ki nastane kot posledica fuzije stacionarnega in migrirajočega jedra. To kompleksno jedro se imenuje sinkarion. Tvorba sinkariona ni nič drugega kot proces oploditve. In pri večceličnih organizmih je bistveni trenutek oploditve zlitje jeder zarodnih celic. Pri ciliatih se zarodne celice ne tvorijo, obstajajo le spolna jedra, ki se zlijejo med seboj. Tako pride do medsebojne navzkrižne oploditve.
Kmalu po nastanku sinkariona se konjuganti ločijo. Glede na zgradbo njihovega jedrskega aparata se v tej fazi še zelo močno razlikujejo od običajnih tako imenovanih nevtralnih (nekonjugiranih) ciliatov, saj imajo le po eno jedro. V prihodnosti se zaradi sinkariona obnovi normalni jedrski aparat. Sinkarij je razdeljen (enkrat ali večkrat). Del produktov te delitve se s kompleksnimi transformacijami, povezanimi s povečanjem števila kromosomov in obogatitvijo s kromatinom, spremeni v makronukleuse. Drugi ohranijo strukturo, značilno za mikronukleuse. Na ta način se obnovi jedrski aparat, ki je značilen in značilen za ciliate, nato pa se trebuščki začnejo nespolno razmnoževati z cepljenjem.
Tako proces konjugacije vključuje dva bistvena biološka momenta: oploditev in obnovo novega makronukleusa zaradi sinkariona.
Kakšen je biološki pomen konjugacije, kakšno vlogo ima v življenju ciliatov? Tega ne moremo imenovati razmnoževanje, saj se število osebkov ne poveča. Zgornja vprašanja so služila kot material za številne eksperimentalne študije, izvedene v mnogih državah. Glavni rezultat teh študij je naslednji. Prvič, konjugacija, tako kot kateri koli drug spolni proces, pri katerem se dva dedna principa (očetovsko in materino) združita v enem organizmu, vodi do povečanja dedne variabilnosti, dedne raznolikosti. Povečanje dedne variabilnosti povečuje prilagoditvene sposobnosti organizma na okoljske razmere. Druga biološko pomembna stran konjugacije je razvoj novega makronukleusa zaradi cepitvenih produktov sinkariona in hkrati uničenje starega. Eksperimentalni podatki kažejo, da ima makronukleus izjemno pomembno vlogo v življenju ciliatov. Nadzira vse glavne življenjske procese in določa najpomembnejše od njih - nastanek (sintezo) beljakovine, ki je glavni del protoplazme žive celice. Pri dolgotrajnem nespolnem razmnoževanju z delitvijo pride do svojevrstnega procesa "staranja" makronukleusa in hkrati celotne celice: aktivnost presnovnega procesa se zmanjša, hitrost delitve se zmanjša. Po konjugaciji (med katero se, kot smo videli, uniči staro makronukleus), se obnovita hitrost presnove in hitrost delitve. Ker do procesa oploditve pride med konjugacijo, ki je pri večini drugih organizmov povezana z razmnoževanjem in pojavom nove generacije, lahko pri ciliatih posameznika, ki nastane po konjugaciji, štejemo tudi za novo spolno generacijo, ki se tukaj pojavlja kot posledica na »pomlajevanje« starega.
Na primeru čevljev ciliatov smo se srečali s tipičnim predstavnikom obsežnega razreda ciliatov. Za ta razred pa je značilna izjemna raznolikost vrst tako v strukturi kot v življenjskem slogu. Oglejmo si podrobneje nekatere najbolj značilne in zanimive oblike.
Pri ciliatih cilije čevlja enakomerno pokrivajo celotno površino telesa. To je značilna lastnost strukture (Holotricha). Za številne cilijate je značilna drugačna narava razvoja ciliarnega pokrova. Dejstvo je, da so cilije ciliatov sposobne, ko so združene, tvoriti bolj zapletene komplekse. Na primer, pogosto opazimo, da se cilije, ki se nahajajo v eni ali dveh vrstah blizu drug drugemu, združijo (lepijo skupaj) in tvorijo ploščo, ki je tako kot cilije sposobna premagati. Takšne lamelne kontraktilne tvorbe imenujemo membranela (če so kratke) ali membrane (če so dolge). V drugih primerih so cilije združene skupaj in se nahajajo v tesnem snopu. Te tvorbe - cirresi - spominjajo na krtačo, katere posamezne dlake so zlepljene skupaj. Različne kompleksne ciliarne tvorbe so značilne za številne ciliate. Zelo pogosto se ciliarni pokrov ne razvije enakomerno, ampak le na nekaterih delih telesa.
trobentač INFUSORIA (SENTOR POLYMORPHIC)
V sladkih vodah pripadajo vrste velikih lepih ciliatov vrste trobentačev(Stentor). To ime je povsem skladno z obliko telesa teh živali, ki res spominja na cev (slika 92), široko odprto na enem koncu. Ob prvem spoznavanju živih trobentačev opazimo eno lastnost, ki ni značilna za čevelj. Ob najmanjšem draženju, tudi mehanskem (na primer tapkanje s svinčnikom po steklu, kjer je kapljica vode s trobentači), se njihovo telo močno in zelo hitro (v delčku sekunde) skrči ter prevzame skoraj redno sferična oblika. Nato se precej počasi (čas se meri v sekundah) trobentač vzravna in prevzame svojo značilno obliko. Ta sposobnost trobentača, da se hitro skrči, je posledica prisotnosti posebnih mišičnih vlaken, ki se nahajajo vzdolž telesa in v ektoplazmi. Tako se lahko mišični sistem razvije tudi v enoceličnem organizmu.

V rodu trobentačev so vrste, za nekatere je značilna precej svetla barva. Zelo pogosta v sladkih vodah modri trobentač(Stentor coeruleus), ki je svetlo modre barve. Ta obarvanost trobentača je posledica dejstva, da se v njegovi ektoplazmi nahajajo najmanjša zrna modrega pigmenta.
Druga vrsta trobentača (Stentor polymorphus) je pogosto obarvana zeleno. Razlog za to obarvanje je precej drugačen. Zelena barva zaradi dejstva, da v endoplazmi ciliata živijo in se razmnožujejo majhne enocelične zelene alge, ki dajejo telesu trobentača značilno barvo. Stentor polymorphus je tipičen primer obojestransko koristnega sobivanja – simbioze. Trobentač in alge sta v medsebojnem simbiotičnem odnosu: trobentač ščiti alge, ki živijo v njegovem telesu, in jih oskrbuje z ogljikovim dioksidom, ki nastane kot posledica dihanja; alge trobentaču zagotavljajo kisik, ki se sprosti v procesu fotosinteze. Očitno del alg prebavijo ciliati, ki so hrana za trobentača.
Trobentači počasi plavajo v vodi s širokim koncem naprej. Lahko pa se začasno pritrdijo na substrat tudi z zadnjim ozkim koncem telesa, na katerem se oblikuje majhen prisesek.
V telesu trobentača je mogoče razlikovati področje trupa, ki se širi od zadaj naprej, in široko perioralno (peristomalno) polje, ki se nahaja skoraj pravokotno nanj. To polje spominja na asimetričen ploščati lijak, na enem robu katerega je vdolbina - žrelo, ki vodi do endoplazme ciliata. Telo trobentača je pokrito z vzdolžnimi vrstami kratkih cilij. Ob robu peristomalnega polja se v krogu nahaja močno razvita blizu ustna (adoralna) cona membranele (slika 92). To območje je sestavljeno iz velikega števila posameznih ciliastih plošč, od katerih je vsaka sestavljena iz številnih cilijev, ki so zlepljeni med seboj, ki se nahajajo v dveh tesno razporejenih vrstah.
V predelu ustne odprtine se perioralne membrane ovijejo proti žrelu in tvorijo levo spiralo. Tok vode, ki ga povzroča nihanje perioralne membrane, je usmerjen proti ustni odprtini (v globino lijaka, ki ga tvori sprednji konec telesa). Skupaj z vodo v žrelo vstopijo tudi delci hrane, suspendirani v vodi. Prehranski predmeti trobentača so bolj raznoliki kot pri copatu. Skupaj z bakterijami poje majhne protozoje (na primer flagellate), enocelične alge itd.
Trobentač ima dobro razvito kontraktilno vakuolo, ki ima svojevrstno strukturo. Osrednji rezervoar se nahaja v sprednji tretjini telesa, nekoliko pod ustno odprtino. Od njega se oddaljita dva dolga aduktivna kanala. Eden od njih poteka od rezervoarja do zadnjega konca telesa, drugi se nahaja v območju peristomalnega polja vzporedno s perioralno cono membranele.
Trobentač infuzorije je priljubljen predmet za eksperimentalne raziskave regeneracije. Številni poskusi so dokazali visoko regenerativno sposobnost trobentačev. Ciliat s tankim skalpelom lahko razrežemo na več delov in vsakega od njih skozi kratek čas(več ur, včasih na dan ali več) se bo spremenil v sorazmerno grajenega, a majhnega trobentača, ki nato zaradi energičnega hranjenja doseže velikost, značilno za to vrsto. Za dokončanje obnovitvenih postopkov mora regeneracijski kos vsebovati vsaj en segment zrnatega makronukleusa.
Trobentač ima, kot smo videli, različne cilije: na eni strani so kratke, pokrivajo celotno telo, na drugi pa je blizu ustna cona membranele. V skladu s to značilnostjo zgradbe je bil poimenovan tudi odred ciliatov, ki mu pripada trobentač ciliated ciliates(Heterotricha).
INFUSORIA BURSARIA (BURSARIA TRUNCATELLA)
Drugi zanimiv predstavnik ciliarnih ciliatov pogosto najdemo v sladkih vodah bursaria(Bursaria truncatella, slika 93). To je velikan med ciliati: njegove dimenzije lahko dosežejo 2 mm, najpogostejše velikosti so 0,5-1,0 mm. Bursaria je jasno vidna s prostim očesom. V skladu s svojim imenom ima bursaria obliko vrečke, široko odprte na sprednjem koncu (bursa je latinska beseda, v ruščini prevedena pomeni "torbica", "vreča") in nekoliko razširjena na zadnjem koncu. Celotno telo ciliata je pokrito z vzdolžnimi vrstami kratkih ciliatov. Njihovo pretepanje povzroči precej počasno premikanje živali naprej. Bursaria plava, kot da se "prevrne" z ene strani na drugo.

Iz sprednjega konca v globino telesa (približno 2/3 njegove dolžine) štrli perioralna depresija - peristom. Na ventralni strani komunicira z zunanjim okoljem skozi ozko režo, na hrbtni strani votlina peristoma ne komunicira z zunanjim okoljem. Če pogledamo na prerez zgornje tretjine telesa bursarije (slika 93, B), lahko vidimo, da peristomska votlina zavzema večji del telesa, medtem ko jo citoplazma obdaja v obliki roba.
Na sprednjem koncu telesa, na levi, izvira iz bursarije zelo močno razvita cona perioralne (adoralne) membranele (slika 93, 4). Spusti se v globino peristomne votline in se obrne v levo. Adoralna cona sega do najglobljega dela peristoma. V peristomalni votlini razen perioralnih membran ni drugih ciliarnih tvorb, razen ciliarnega traku, ki poteka vzdolž ventralne strani peristomne votline (sl. 93, 10). Navznoter zadnja stena peristomalna votlina ima po skoraj celotni dolžini ozko režo (slika 93, 7), katere robovi se običajno tesno prilegajo drug drugemu. To je odpiranje ust. Njegovi robovi se odmaknejo le v času jednja.
Bursarije nimajo ozke prehranske specializacije, so pa večinoma plenilci. Ko se premikajo naprej, naletijo na različne male živali. Zahvaljujoč delu membrane blizu ustne cone se plen s silo potegne v obsežno peristomalno votlino, od koder ne more več izplavati. Prehrambeni predmeti se pritisnejo na hrbtno steno peristomalne votline in prodrejo v endoplazmo skozi razširjeno ustno razpoko. Bursarije so zelo požrešne, lahko pogoltnejo precej velike predmete: na primer, njihova najljubša hrana so ciliati za čevlje. Bursaria lahko pogoltne 6-7 čevljev zapored. Posledično v endoplazmi bursarije nastanejo zelo velike prebavne vakuole.
Jedrski aparat bursarije je precej zapleten. Imajo eno dolgo makronukleus v obliki klobase in veliko (do približno 30) število majhnih mikronukleusov, naključno raztresenih v endoplazmi ciliata.
Bursaria so med redkimi vrstami sladkovodnih ciliatov, ki nimajo kontraktilne vakuole. Kako se izvaja osmoregulacija v tem velikem ciliatu, še vedno ni povsem jasno. Pod ektoplazmo bursarije na različnih delih telesa lahko opazimo različne oblike in velikosti mehurčkov tekočine - vakuole, ki spreminjajo svojo prostornino. Očitno te nepravilne oblike vakuole in po svoji funkciji ustrezajo kontraktilnim vakuolam drugih ciliatov.

Zanimivo je opazovati zaporedne faze nespolnega razmnoževanja Bursaria. Na začetnih stopnjah delitve pride do popolnega zmanjšanja celotne votline peristoma in blizu ustne cone membranele (slika 94). Ohranjen je le zunanji cilij. Infuzorija ima obliko jajca. Po tem je telo zavezano s prečnim utorom na dve polovici. V vsakem od nastalih hčerinskih ciliatov se s precej zapletenimi transformacijami razvijeta tipičen peristom in perioralna cona membranele. Celoten postopek delitve bursarije poteka hitro in traja nekaj več kot eno uro.
Pri bursariji je zelo enostavno opazovati še en pomemben življenjski proces, katerega začetek je povezan s pogoji, ki so neugodni za ciliate, proces nastajanja cist (encistacija). Ta pojav je značilen na primer za amebo. Toda izkazalo se je, da lahko celo tako zapleteno organizirani protozoji, kot so ciliati, preidejo v neaktivno stanje. Če kulture, v kateri živijo bursarije, ne nahranimo ali ohladimo pravočasno, se bo čez nekaj ur začela množična encistacija. Ta postopek poteka na naslednji način. Bursaridi, tako kot pred delitvijo, izgubijo peristome in perioralno cono membran. Nato postanejo popolnoma sferične, nakar ločijo dvojno lupino značilne oblike (slika 94, D).
V stanju cist je bursarija lahko več mesecev. Ko se pojavijo ugodni pogoji, lupina ciste poči, iz nje izstopijo bursarije, razvijejo peristome in preidejo v aktivno življenje.
STYLONICHIA MYTILUS
Zelo zapleten in raznoliko diferenciran ciliarni aparat ima sorodne trepalnice odvajanje prebavil(Hypotricha), katere številne vrste živijo v sladki in morski vodi. Eden najpogostejših, pogosto srečenih predstavnikov tega zanimiva skupina se lahko pokliče stilonychia(Stylonichia mytilus). To je precej velik ciliat (dolžine do 0,3 mm), ki živi na dnu sladkovodnih rezervoarjev, na vodna vegetacija(slika 95). Za razliko od copate, trobentača in bursarije stilonihija nima neprekinjenega ciliarnega pokrova, celoten ciliarni aparat pa predstavlja omejeno število strogo določenih ciliarnih tvorb.

Telo Stilonychia (tako kot večina drugih trebušnih trebušnih trebuščkov) je močno sploščeno v hrbtno-trebušni smeri, njegove hrbtne in ventralne strani, sprednji in zadnji konec pa so jasno razločljive. Telo je spredaj nekoliko razširjeno, zadaj zoženo. Pri pregledu živali s trebušne strani se jasno vidi, da je v sprednji tretjini na levi kompleksno urejena pernata in ustna odprtina.
Na hrbtni strani se precej redko nahajajo cilije, ki niso sposobne premagati. Raje jih lahko imenujemo tanke elastične ščetine. So negibni in nimajo nobene zveze s funkcijo gibanja. Tem cilijam je običajno dodeljena taktilna, občutljiva funkcija.
Vse ciliarne tvorbe, povezane z gibanjem in zajemanjem hrane, so skoncentrirane na ventralni strani živali (slika 95). Obstaja majhno število debelih prstastih formacij, ki se nahajajo v več skupinah. To so trebušni ciri. Vsak od njih je zapletena ciliarna tvorba, ki je rezultat tesne povezave (lepljenja) več deset posameznih cilij. Tako so okrogle kot čopiči, katerih posamezne dlake so tesno združene in povezane med seboj.
S pomočjo ciroza se žival premika precej hitro, "teče" vzdolž substrata. Poleg "plazanja" in "tekanja" po substratu je stilonychia sposobna narediti precej ostre in močne skoke, ki se takoj odlepijo od substrata. Ti ostri gibi se izvajajo s pomočjo dveh močnih repnih krogov (slika 95), ki ne sodelujeta pri običajnem "plazenju".
Dve vrsti obrobnih (obrobnih) cirij se nahajata ob robu telesa na desni in levi strani. Z desnega roba živali potekajo po celem telesu, z levega roba pa le do predela peristoma. Te ciliarne tvorbe služijo za poganjanje živali, ko se loči od substrata in prosto plava v vodi.
Vidimo torej, da raznolik in specializiran ciliarni aparat stilonihije omogoča izvajanje zelo raznolikih gibov, v nasprotju z, na primer, preprostim drsenjem v vodi, kot je čevelj ali trobentač.
Zapleten je tudi ciliarni aparat, povezan s funkcijo prehrane. Videli smo že, da se skoraj ustna vdolbina (peristom), na dnu katere se nahaja ustna odprtina, ki vodi v žrelo, nahaja v sprednji polovici živali na levi strani. Ob levem robu, ki se začne od samega sprednjega konca telesa, je močno razvita cona perioralnih (adoralnih) membranol. S svojim udarcem usmerjajo tok vode proti ustni odprtini. Poleg tega so v predelu peristomalnega vdolbine še tri kontraktilne membrane (membrane), ki segajo v žrelo z notranjimi konci, in številne posebne perioralne cilije (slika 95). Celoten ta kompleksen aparat služi za zajemanje in usmerjanje hrane v ustno odprtino.
Stilonychia je ena od protozojev z zelo širokim naborom prehranskih predmetov. Upravičeno ga lahko imenujemo vsejed. Lahko poje, kot čevelj, bakterije. Med njenimi prehranjevalnimi predmeti so flagelati, enocelične alge (pogosto diatomeje). Končno je Stilonychia lahko tudi plenilec, ki napada druge, manjše vrste ciliatov in jih požre.
Stilonychia ima kontraktilno vakuolo. Sestavljen je iz osrednjega rezervoarja, ki se nahaja na levem zadnjem koncu peristoma in enega adduktorskega kanala, usmerjenega nazaj.
Jedrski aparat, kot vedno pri ciliatih, je sestavljen iz makronukleusa in mikronukleusa.
Makronukleus je sestavljen iz dveh polovic, povezanih s tanko zožitvijo; obstajata dve mikronukleusi, ki se nahajata neposredno blizu obeh polovic Ma.
Stilonychia, delno bursaria, trobentač - vse to so ciliati s široko paleto prehranskih predmetov. Sposobnost vpijanja različnih živil je značilna za večino ciliatov. Vendar pa je med njimi mogoče najti tudi vrste, ki so strogo specializirane glede na naravo hrane.
INFUZORIJSKI PLENILCI
Med ciliati so plenilci, ki so zelo »izbirčni« glede svojega plena. Dober primer je infuzorija. didinija(Didinium nasutum). Didinium je relativno majhen ciliat, v povprečju dolg približno 0,1-0,15 mm. Sprednji konec je podolgovat v obliki proboscisa, na koncu katerega je odprtina za usta. Ciliarni aparat predstavljata dva venčka cilia (slika 96). Didinius hitro plava v vodi, pogosto spreminja smer. Najprimernejša hrana didinije so ciliati za čevlje. V tem primeru je plenilec manjši od plena. Didinius s prtljažnikom prodre v plen, nato pa, postopoma vse bolj in bolj širi odprtino ust, pogoltne čevelj celega! V proboscisu je posebna, tako imenovana palica, aparat. Sestavljen je iz številnih elastičnih močnih palic, ki se nahajajo v citoplazmi vzdolž obrobja proboscisa. Menijo, da ta naprava poveča trdnost sten proboscisa, ki ne poči pri požiranju tako velikega plena v primerjavi z didinijem, kot je čevelj. Didinius - primer zadnje zatočišče plenilstvo med protozoji. Če didinij pogoltne svoj plen – čevlje – primerjamo s plenilcem pri višjih živalih, potem je težko najti podobne primere.

Didinius, požiranje paramecije seveda zelo nabrekne. Proces prebave je zelo hiter, pri sobni temperaturi traja le približno dve uri. Nato neprebavljene ostanke vržejo ven in didinij začne loviti drugo žrtev. Posebne študije so pokazale, da je dnevna "prehrana" didinije 12 čevljev - resnično ogromen apetit! Upoštevati je treba, da se v intervalih med naslednjimi "lovi" didinia včasih razdeli. Ob pomanjkanju hrane didinija zelo zlahka encistira in prav tako zlahka spet izstopi iz cist.
rastlinojedi ciliati
Precej redkeje kot plenjenje, najdemo ga med ciliati "čisto vegetarijanstvo" - prehranjevanje izključno rastlinska hrana. Eden redkih primerov "vegetarijanskih" ciliatov so lahko predstavniki rod pasula(Nassula). Njihov prehranski objekt so nitaste modrozelene alge (slika 97).

Skozi usta, ki se nahajajo ob strani, prodrejo v endoplazmo, nato pa jih infuzorija zvije v tesno spiralo, ki se postopoma prebavi. Pigmenti alg delno vstopijo v citoplazmo ciliatov in jo obarvajo v svetlo temno zeleno barvo.
SUVOIKA (VORTICELLA NEBULIFERA)
Zanimiva in po številu vrst precej velika skupina ciliatov so sedeče oblike, pritrjene na substrat, ki tvorijo odcep okroglih oči(Peritricha). Najpogostejši člani te skupine so suvoyki(vrste iz rodu Vorticella).
Suvojki spominjajo na eleganten cvet, kot je zvonec ali šmarnica, ki sedi na dolgem peclju, ki je s koncem pritrjen na substrat. Suvoyka preživi večino svojega življenja v stanju, pritrjenem na substrat.

Razmislite o strukturi telesa ciliatov. Pri različnih vrstah se njihove velikosti razlikujejo v precej širokem razponu (do približno 150 mikronov). Ustna ploščica (slika 98) se nahaja na razširjenem sprednjem delu telesa, ki je popolnoma brez cilijev. Ciliarni aparat se nahaja le ob robu ustnega (peristomalnega) diska (slika 98) v posebnem utoru, zunaj katerega se oblikuje valj (peristomalna ustnica). Ob robu valja potekajo tri ciliirane membrane, od katerih sta dve nameščeni navpično, ena (zunanja) je vodoravna. Tvorijo nekaj več kot en polni obrat vijačnice. Te membrane so v nenehnem utripajočem gibanju in usmerjajo tok vode v ustno odprtino. Ustni aparat se začne precej globoko z lijakom na robu peristomalnega polja (slika 98), v globini katerega je ustna odprtina, ki vodi v kratko žrelo. Suvoji se, tako kot čevlji, hranijo z bakterijami. Njihova ustna odprtina je nenehno odprta, proti ustjem pa je neprekinjen tok vode.
Ena kontraktilna vakuola brez aduktorskih kanalov se nahaja v bližini ustne odprtine. Makronukleus ima obliko, podobno traku ali klobasi, tesno ob njej je eno majhno mikronukleus.
Suvoyka je sposobna močno skrajšati pecelj, ki ga v delčku sekunde zasukamo z odmašnikom. Hkrati se telo ciliata tudi skrči: peristomalni disk in membrane se potegnejo navznoter in celoten sprednji konec se zapre.

Seveda se postavlja vprašanje: ker so suvoji pritrjeni na substrat, na kakšen način se izvaja njihovo usedanje v rezervoar? To se zgodi z oblikovanjem prosto lebdeče faze, potepuha. Na zadnjem koncu ciliatnega telesa se pojavi venček cilia (slika 99). Istočasno se peristomalni disk umakne navznoter in trepavič se loči od peclja. Nastali potepuh lahko plava več ur. Dogodki se nato odigrajo v obratni vrstni red: ciliat se s svojim zadnjim koncem pritrdi na substrat, pecelj raste, zadnji venček cilia se zmanjša, peristomalni disk se na sprednjem koncu zravna in začnejo delovati adoralne membrane. Nastajanje potepuhov v suvoyki je pogosto povezano s procesom aseksualnega razmnoževanja. Infuzorija na peclju se razdeli in eden od hčerinskih posameznikov (in včasih oba) postane potepuh in odplava.
Številne vrste suvokov so sposobne encistirati v neugodnih pogojih.
Med sedečimi ciliati, ki spadajo v skupino ciliatov, je le sorazmerno malo vrst, kot so zgoraj obravnavane suvoje, samotne žive oblike. Večina tukaj vključenih vrst so kolonialni organizmi.
Običajno je kolonialnost posledica nepopolno dokončanega aseksualnega ali vegetativnega razmnoževanja. Posamezniki, ki so nastali kot posledica razmnoževanja, v večji ali manjši meri ohranjajo medsebojno povezanost in skupaj tvorijo organsko individualnost višjega reda, ki združuje velike količine posamezne posameznike, ki dobijo ime kolonije (s primeri kolonialnih organizmov smo se že srečali v razred flagelatov.

Kolonije okroglih ciliatov nastanejo kot posledica dejstva, da se ločeni posamezniki ne spremenijo v potepuhe, ampak ohranjajo stik med seboj s pomočjo pecljev (slika 100). V tem primeru glavnega stebla kolonije, pa tudi njenih prvih vej, ni mogoče pripisati nobenemu od posameznikov, ampak pripada celotni koloniji kot celoti. Včasih kolonijo sestavlja le majhno število posameznikov, medtem ko lahko pri drugih vrstah ciliatov število posameznih posameznikov kolonije doseže več sto. Vendar pa rast katere koli kolonije ni neomejena. Ko dosežejo dimenzije, značilne za to vrsto, se kolonija preneha povečevati in posamezniki, ki nastanejo kot posledica delitve, razvijejo cilijev venček, postanejo potepuhi in odplavajo, kar povzroči nove kolonije.
Kolonije okroglih ciliatov so dveh vrst. Pri nekaterih je pecelj kolonije nereduciran: ko so razdraženi, se sklenejo le posamezni posamezniki kolonije, ki se vrišejo v pernato, vendar se celotna kolonija kot celota ne spremeni (ta vrsta kolonije vključuje npr. Epistylis, Opercularia). Pri drugih (na primer rod Carchesium) se lahko steblo celotne kolonije skrči, saj citoplazma prehaja skozi vse veje in tako povezuje vse posameznike kolonije med seboj. Ko so takšne kolonije razdražene, se popolnoma skrčijo. Celotna kolonija v tem primeru reagira kot celota, kot organska individualnost.
Med vsemi kolonialnimi okroglimi trepalkami je morda še posebej zanimiv zootamnično(Zoothamnium arbuscula). Kolonije tega ciliata odlikuje posebna pravilnost strukture. Poleg tega je tukaj znotraj kolonije orisan zanimiv biološki pojav polimorfizma.

Kolonija zootamnije je videti kot dežnik. Na enem, glavnem, steblu kolonije so sekundarne veje (slika 101). Velikost odrasle kolonije je 2-3 mm, zato so jasno vidne s prostim očesom. Zootamnii živijo v majhni ribniki od čisto vodo. Njihove kolonije se običajno nahajajo na podvodnih rastlinah, najpogosteje na elodeji (vodna kuga).
Stebla kolonije zootamnije so kontraktilna, saj kontraktilna citoplazma prehaja skozi vse veje kolonije, razen bazalnega dela glavnega peclja. Z zmanjšanjem, ki se zgodi zelo hitro in nenadno, se celotna kolonija zbere v kepo.

Za zootamnijo je značilna strogo pravilna razporeditev vej. Eno glavno steblo je pritrjeno na substrat. Devet glavnih vej kolonije odstopa od njenega zgornjega dela v ravnini, pravokotni na pecelj, ki se strogo redno nahaja ena glede na drugo (slika 102, 6). Od teh vej segajo sekundarne veje, na katerih sedijo posamezni posamezniki kolonije. Vsaka sekundarna veja ima lahko do 50 ciliatov. Skupno število posameznikov v koloniji doseže 2-3 tisoč posameznikov.
Večina posameznikov kolonije po svoji strukturi spominja na majhne posamezne suvoike, velike 40-60 mikronov. Toda poleg majhnih posameznikov, ki jih imenujemo mikrozoidi, se na odraslih kolonijah, približno na sredini glavnih vej, razvijejo posamezniki povsem drugačnega tipa in velikosti (slika 102, 5). To so veliki sferični posamezniki s premerom 200-250 mikronov, ki po masi presegajo prostornino mikrozoida za sto ali več krat. Veliki posamezniki se imenujejo makrozoidi.
Po svoji strukturi se bistveno razlikujejo od majhnih posameznikov kolonije. Njihov peristom ni izražen: potegnjen je navznoter in ne deluje. Makrozoan od samega začetka razvoja iz mikrozoida preneha jemati hrano sam. Manjka prebavne vakuole. Rast makrozoida se očitno izvaja na račun snovi, ki vstopajo skozi citoplazemske mostove, ki povezujejo vse posameznike kolonije med seboj. V delu telesa makrozoja, s katerim je pritrjen na pecelj, se kopičijo posebna zrna (zrnca), ki imajo, kot bomo videli, pomembno vlogo pri njegovi nadaljnji usodi. Kaj so ti veliki sferični makrozoidi, kakšna je njihova biološka vloga v življenju kolonije zootamnije? Opazovanje kaže, da so makrozoidi bodoči potepuhi, iz katerih se razvijejo nove kolonije. Doseganje omejitev velikosti, makrozoid razvije venček cilije, se loči od kolonije in odplava. Hkrati se njegova oblika nekoliko spremeni, iz sferične postane stožčasta. Po določenem času je tramp vedno pritrjen na substrat s stranjo, na kateri se nahaja granularnost. Takoj se začne tvorba in rast peclja, za gradnjo peclja pa se porabijo zrnca, ki so lokalizirana na zadnjem koncu potepuha. Ko steblo raste, zrnatost izgine. Ko pecelj doseže končno dolžino, značilno za zootamnijo, se začne serija hitro zaporednih delitev, ki vodijo v nastanek kolonije. Te delitve so narejene v strogo določenem zaporedju (slika 102).
Ne bomo se zadrževali na podrobnostih tega postopka. Bodimo pozorni le na naslednji zanimiv pojav. Med prvimi delitvami potepuhov zootamnije, med razvojem kolonije pri nastajajočih posameznikih, pernati in ustje ne delujeta. Hranjenje se začne kasneje, ko je mlada kolonija že sestavljena iz 12-16 posameznikov. Tako se vse prve stopnje razvoja kolonije izvajajo izključno na račun tistih rezerv, ki so nastale v telesu makrozoida med njegovo rastjo in razvojem na matični koloniji. Obstaja nesporna podobnost med razvojem zootamnije vagrant in razvojem jajčeca pri večceličnih živalih. Ta podobnost se izraža v tem, da razvoj v obeh primerih poteka na račun predhodno nakopičenih rezerv, brez zaznavanja hrane iz zunanjega okolja.
Pri preučevanju sedečih okroglih ciliatov se postavlja vprašanje: kako se izvaja oblika spolnega procesa, značilna za ciliate - konjugacija? Izkazalo se je, da v povezavi s sedečim načinom življenja doživi nekaj pomembnih sprememb. Do začetka spolnega procesa se na koloniji tvorijo posebni, zelo majhni potepuhi. Aktivno se gibljejo s pomočjo venčka cilijev, nekaj časa plazijo vzdolž kolonije, nato pa pridejo v konjugacijo z velikimi normalnimi sedečimi posamezniki kolonije. Tako pride do diferenciacije konjugantov v dve skupini posameznikov: majhne, mobilne (mikrokonjuganti) in večje, nepremične (makrokonjuganti). Ta diferenciacija konjugantov v dve kategoriji, od katerih je ena (mikrokonjuganti) mobilna, je bila nujna prilagoditev sedečemu življenjskemu slogu. Brez tega normalnega poteka spolnega procesa (konjugacije) očitno ne bi bilo mogoče zagotoviti.
SESANJE CILITARIUSA (SUCTORIA)
Zelo svojevrstno skupino po načinu prehranjevanja predstavljajo sesanje ciliatov(Suctoria). Ti organizmi, tako kot suvoyka in drugi ciliati, so sedeči. Število vrst, ki spadajo v ta red, se meri z več desetinami. Oblika telesa sesalnih ciliatov je zelo raznolika. Nekatere njihove značilne vrste so prikazane na sliki 103. Nekatere sedijo na substratu na bolj ali manj dolgih pecljih, druge nimajo pecljev, pri nekaterih se telo precej močno razveja itd. Kljub raznolikosti oblik pa vsi sesajoči ciliati zanje sta značilni naslednji dve značilnosti:
1) popolna odsotnost (pri odraslih oblikah) ciliarnega aparata,
2) prisotnost posebnih dodatkov - lovk, ki služijo za sesanje plena.

Pri različnih vrstah sesalnih ciliatov število lovk ni enako. Pogosto so zbrani v skupinah. Z veliko povečavo mikroskopa je razvidno, da je na koncu lovka opremljena z majhno zadebelitev v obliki palice.
Kako delujejo lovke? Na to vprašanje ni težko odgovoriti tako, da nekaj časa opazujemo sesanje trepalk. Če se kakšen majhen pražival (bička, infuzorija) dotakne lovke suctoria, se bo takoj prilepil nanjo. Vsi poskusi žrtve, da se umakne, so običajno zaman. Če še naprej opazujete plen, pritrjen na lovke, lahko vidite, da se postopoma začne zmanjševati. Njena vsebina se postopoma »načrpa« skozi lovke v endoplazmo sesajoče trepalnice, dokler od žrtve ne ostane le ena pellik, ki jo zavržemo. Tako so lovke sesalnih ciliatov povsem edinstvene, nikjer drugje v živalskem svetu ni organov za lovljenje in hkrati sesanje hrane (slika 103).
Sesalni ciliati so negibni plenilci, ki plena ne preganjajo, ampak ga takoj ujamejo, če se jih samo nepreviden plen dotakne.

Zakaj te nenavadne organizme uvrščamo v razred ciliatov? Na prvi pogled nimajo nič z njimi. Naslednja dejstva govorijo o pripadnosti suctoria ciliatom. Prvič, imajo jedrski aparat, značilen za ciliate, sestavljen iz makronukleusa in mikronukleusa. Drugič, med razmnoževanjem razvijejo cilije, ki jih pri "odraslih" posameznikih ni. Nespolno razmnoževanje in hkrati preselitev sesalnih ciliatov se izvaja z tvorbo potepuhov, opremljenih z več obročastimi venčki cilijev. Nastajanje potepuhov v suktoriji se lahko pojavi na različne načine. Včasih nastanejo kot posledica ne povsem enotne delitve (brstenja), pri kateri vsaka ledvica, ki se loči navzven, prejme segment makronukleusa in eno mikronukleus (slika 104, L). Na enem materinem posamezniku lahko naenkrat nastane več hčerinskih brstov (slika 104, 5). Pri drugih vrstah (slika 104, D, E) opazimo zelo svojevrsten način "notranjega brstenja". Hkrati se znotraj telesa materinega suktorja oblikuje votlina, v kateri nastane potepuška ledvica. Izstopi skozi posebne luknje, skozi katere se z znano težavo "stisne".
Ta razvoj zarodka v materinem telesu in nato dejanje rojevanja otroka je zanimiva analogija najpreprostejšega s tem, kar se dogaja v višjih večceličnih organizmih.
Na prejšnjih straneh je bilo obravnavanih več tipičnih prostoživečih predstavnikov razreda ciliatov, ki so različno prilagojeni različnim okoljskim razmeram. Zanimivo je pristopiti k vprašanju prilagajanja ciliatov življenjskim razmeram in po drugi strani videti, kakšne so značilnosti skupne značilnosti ciliatov, ki živijo v določenih, natančno določenih okoljskih razmerah.
Za primer vzemimo dva zelo različna habitata: življenje v sestavi planktona in življenje na dnu v debelini peska.
PLANKTONSKA INFUZORIJA
V morskem in sladkovodnem planktonu najdemo precej veliko število ciliatov.
Posebnosti prilagajanja na življenje v vodnem stolpcu so še posebej izrazite pri radiolarjih. Glavna smer prilagajanja na planktonski način življenja je zmanjšana na razvoj takšnih strukturnih značilnosti, ki prispevajo k dvigu organizma v vodnem stolpcu.

Tipična planktonska, poleg tega skoraj izključno morska družina ciliatov je tinidi(Tintinnidae, slika 105, 5). Skupno število doslej znanih vrst tinidov je približno 300. Gre za majhne oblike, za katere je značilno, da je protoplazmatsko telo trepalke postavljeno v prozorno, svetlo in hkrati močno hišo, sestavljeno iz organske snovi. Iz hiše štrli disk, ki nosi venček cilijev, ki so v nenehnem utripajočem gibanju. V stanju lebdeče infuzorije v vodnem stolpcu ga podpira predvsem nenehno aktivno delo ciliarnega aparata. Hiša očitno opravlja funkcijo zaščite spodnjega dela ciliata. V sladki vodi živita le 2 vrsti tintinnidov (ne štejemo 7 vrst, značilnih samo za Bajkalsko jezero).
Sladkovodni ciliati imajo še nekatere druge prilagoditve na življenje v planktonu. Pri mnogih je citoplazma zelo močno vakuolizirana (Loxodes, Condylostoma, Trachelius), tako da je podobna peni. To vodi do znatnega zmanjšanja specifične teže. Vsi našteti ciliati imajo poleg tega cilijarno prevleko, zahvaljujoč kateri je telo ciliatov glede na specifično težo le nekoliko večje od specifična težnost voda, ki se zlahka vzdržuje v "visečem" stanju. Pri nekaterih vrstah oblika telesa prispeva k povečanju specifične površine in olajša lebdenje v vodi. Nekateri planktonski ciliati Bajkalskega jezera so na primer po obliki podobni dežniku ali padalu (Liliomorpha, slika 105, 2). V Bajkalskem jezeru je en planktonski sesalni trebušček (Mucophrya pelagica, slika 105, 4), ki se močno razlikuje od svojih sedečih sorodnikov. Ta vrsta je brez stebla. Njegovo protoplazmatsko telo je obdano s širokim sluzastim plaščem, prilagoditvijo, ki vodi do zmanjšanja telesne teže. Izstopajo dolge tanke lovke, ki poleg svoje neposredne funkcije verjetno opravljajo še eno - povečanje specifične površine, kar prispeva k lebdenju v vodi.
Na koncu je treba omeniti še eno, tako rekoč posredno obliko prilagajanja ciliatov na življenje v planktonu. To je navezanost majhnih ciliatov na druge organizme, ki vodijo planktonski življenjski slog. Ja, med obrezani ciliati(Peritricha) obstajajo precej številne vrste, ki se vežejo na planktonske kopepode. To je normalen in normalen način življenja za te vrste ciliatov.
Skupaj s cirkumciliarji in med sesanje(Suctoria) obstajajo vrste, ki se naselijo na planktonskih organizmih.
CILITICI, KI ŽIVIJO V PESKU
Peščene plaže in plitvine predstavljajo izjemno svojevrsten habitat. Ob obali morja zasedajo velike prostore in zanje je značilna svojevrstna favna.
Izvedeno za Zadnja letaŠtevilne študije v različnih državah so pokazale, da je plast številnih morskih peskov zelo bogata z različnimi mikroskopskimi ali mikroskopskimi živalmi. Med delci peska so številni majhni in drobni prostori, napolnjeni z vodo. Izkazalo se je, da so ti prostori bogato naseljeni z organizmi, ki pripadajo najrazličnejšim skupinam živalskega sveta. Tu živi na desetine vrst rakov, anelidi, okrogli črvi, zlasti številni ploski črvi, nekateri mehkužci, črvi črvi. V velikem številu so tudi protozoji, predvsem ciliati. Po sodobnih podatkih favna ciliatov, ki naseljujejo debelino morskega peska, vključuje približno 250-300 vrst. Če ne upoštevamo le ciliatov, temveč tudi druge skupine organizmov, ki naseljujejo debelino peska, bo skupno število njihovih vrst zelo veliko. Celoten nabor živali, ki naseljujejo debelino peska in živijo v najmanjših vrzeli med zrni peska, se imenuje psamofilna favna.
Bogastvo in vrstno sestavo psamofilne favne določajo številni dejavniki. Med njimi predvsem pomembnosti ima velikost peščenih delcev. Grobozrnati pesek ima slabo favno. Slaba je tudi favna zelo drobnozrnatih meljastih pekov (s premerom delcev manj kot 0,1 mm), kjer so očitno reže med delci premajhne, da bi v njih živele živali. Najbogatejši pesek je srednje in drobnozrnat.
Drugi dejavnik, ki igra pomembno vlogo pri razvoju psamofilne favne, je bogastvo peska v organskih ostankih in razpadajočih organskih snoveh (t. i. stopnja saprobnosti). Pesek brez organskih snovi je reven v življenju. Po drugi strani pa je pesek tudi skoraj brez življenja in je zelo bogat z organskimi snovmi, saj razpad organske snovi vodi v pomanjkanje kisika. Pogosto se temu doda anaerobna fermentacija vodikovega sulfida.
Prisotnost prostega vodikovega sulfida je izjemno negativen dejavnik, ki vpliva na razvoj favne.
V površinskih plasteh peska se včasih razvije dokaj bogata flora enoceličnih alg (diatomeji, peridinij). To je dejavnik, ki spodbuja razvoj psamofilne favne, saj se veliko majhnih živali (vključno z ciliati) prehranjuje z algami.
Končno, dejavnik, ki zelo negativno vpliva na psamofilno favno, je surf. To je povsem razumljivo, saj surf, ki preplavi zgornje plasti peska, ubije tukaj vsa živa bitja. Psamofilna favna je najbogatejša v zaščitenih, dobro ogrevanih zalivih. Plivanje in oseka ne preprečujeta razvoja psamofilne favne. Ko voda ob oseki začasno odide in izpostavi pesek, potem v debelini peska, v presledkih med zrni peska, ostane, in to ne preprečuje obstoja živali.
Pri ciliatih, ki so del psamofilne favne in spadajo v različne sistematične skupine (redove, družine), se v procesu evolucije razvijejo številne skupne značilnosti, ki so prilagoditve na posebne pogoje obstoja med peščenimi delci.

Slika 106 prikazuje nekatere vrste psamofilne favne ciliatov različnih vrst in družin. Med njima je veliko podobnosti. Telo večine je po dolžini bolj ali manj močno podolgovate, črvasto. Tako se zlahka »stisne« v najmanjše luknje med zrni peska. Pri zelo številnih vrstah (slika 106) se raztezanje telesa kombinira z njegovo sploščenostjo. Ciliarni aparat je vedno dobro razvit, kar omogoča, da se aktivno, z določeno silo, premika v ozkih vrzeli. Pogosto se na eni strani črvičastega sploščenega telesa razvijejo cilije, nasprotna stran je gola. Ta lastnost je verjetno povezana s sposobnostjo, ki je izrazita pri večini psamofilnih vrst, da se zelo tesno in zelo trdno oprime (pritrdi) na substrat skozi ciliarni aparat (pojav, imenovan tigmotaksija). Ta lastnost omogoča živalim, da ostanejo na mestu v primerih, ko v ozkih vrzelih, kjer živijo, nastanejo tokovi vode. V tem primeru je verjetno bolj ugodno, da je stran, nasprotna tisti, na kateri se je žival pritrdila na substrat, gladka.
Kaj jedo psamofilni ciliati? Pomemben del "prehrane" mnogih vrst so alge, zlasti diatomeje. Bakterije jim v manjši meri služijo kot hrana. V veliki meri je odvisno tudi od tega, da je v pesku malo bakterij, ki niso močno onesnažene. Nazadnje, zlasti med največjimi psamofilnimi ciliati, obstaja precejšnje število plenilskih oblik, ki jedo druge ciliate, ki pripadajo manjšim vrstam. Psamofilni ciliati so očitno razširjeni povsod.
APOSTOMATI INFUSORIA

ciliatov spirofrija(Spirophrya subparasitica) v encistiranem stanju lahko pogosto najdemo sedečega na majhnem peclju na majhnih planktonskih morskih rakih (zlasti pogosto na rakih iz rodu Idia). Medtem ko rak aktivno plava v morski vodi, se spirofrija, ki sedi na njem, ne spreminja. Za nadaljnji razvoj ciliatov je potrebno, da rak poje morski hidroidni polip, kar se zgodi precej pogosto (slika 107). Takoj, ko ciste spirofrije skupaj z rakom vstopijo v prebavno votlino, iz njih takoj izstopijo majhni ciliati, ki se začnejo energijsko hraniti s prehransko kašo, ki nastane kot posledica prebave pogoltnjenega raka. V eni uri se velikost infuzorije poveča za 3-4 krat. Vendar se v tej fazi razmnoževanje ne zgodi. Pred nami je tipična stopnja rasti ciliatov, ki se imenuje trofont. Čez nekaj časa, skupaj z neprebavljenimi ostanki hrane, trofont vrže polip ven v morsko vodo. Tukaj, aktivno plava, se spusti vzdolž telesa polipa do njegovega podplata, kjer se pritrdi, obkrožen s cisto. Ta stopnja velikega ciliata, ki se nahaja na polipu, se imenuje tomont. To je faza razmnoževanja. Tomont se ne hrani, ampak se hitro večkrat zaporedoma deli (slika 107, 7). Rezultat je cela skupina zelo majhnih ciliatov. Njihovo število je odvisno od velikosti tomonta, ki pa je odvisna od velikosti trofonta, ki mu je dal izvor. Majhni ciliati, ki nastanejo kot posledica delitve tomonta (imenujejo se tomiti ali potepuhi), predstavljajo stopnjo poselitve.
Zapustijo cisto, hitro plavajo (ne da bi jedli hkrati, ampak z uporabo rezerv, ki jih imajo v citoplazmi). Če imajo »srečo«, da naletijo na kopepoda, se nanj takoj pritrdijo in encistirajo. To je faza, s katere smo začeli obravnavati cikel.
V pregledanem življenski krog spirofrija opozarja na ostro razmejitev stopenj, ki imajo različen biološki pomen. Trofon je stopnja rasti. Zaradi močne in hitre prehrane samo raste, kopiči veliko količino citoplazme in vseh vrst rezervnih snovi. Trofont ni sposoben razmnoževanja. Pri tomontu opazimo obraten pojav - nezmožnost hranjenja in močno hitro razmnoževanje. Po vsaki delitvi ni rasti, zato se razmnoževanje tomonta zmanjša na hitro propadanje na številne potepuhe. Končno, potepuhi opravljajo svojo posebno in edino značilno funkcijo: so posamezniki - naseljenci in distributerji vrste. Ne morejo jesti ali razmnoževati.
ŽIVLJENJSKI CIKLUS Ichthyophthiriusa

Do konca obdobja rasti ichthyophthirius doseže zelo veliko velikost v primerjavi s potepuhi: 0,5-1 mm v premeru. Ko dosežejo mejno vrednost, se ciliati aktivno premikajo iz ribjih tkiv v vodo in nekaj časa počasi plavajo s pomočjo ciliarnega aparata, ki pokriva celotno telo. Kmalu se veliki ihtioftirije naselijo na nekem podvodnem predmetu in izločijo cisto. Takoj po encistiranju se začnejo zaporedne delitve ciliatov: najprej na polovico, nato se vsak hčerinski posameznik ponovno razdeli na dva in tako naprej do 10-11 krat. Posledično se znotraj ciste oblikuje do 2000 majhnih, skoraj zaobljenih posameznikov, pokritih z cilijami. V notranjosti ciste se potepuhi aktivno gibljejo. Preluknjajo lupino in pridejo ven. Aktivno plavajoči potepuhi okužijo nove ribe.
Hitrost delitve ihtioftirije v cistah, pa tudi hitrost njegove rasti v ribjih tkivih, je v veliki meri odvisna od temperature. Glede na študije različnih avtorjev so podane naslednje številke: pri 26-27°C proces razvoja potepuhov v cisti traja 10-12 ur, pri 15-16°C traja 28-30 ur, pri 4 -5°C, traja 6 -7 dni.
Boj proti ihtioftiriju predstavlja velike težave. Pri tem so najpomembnejši preventivni ukrepi, katerih cilj je preprečiti, da bi prosto plavajoči potepuhi prodrli v tkiva rib. Za to je koristno izvajati pogosto presaditev bolnih rib v nove rezervoarje ali akvarije, da ustvarite pretočne pogoje, kar je še posebej učinkovito v boju proti ihtioftiriju.
ciliates trichodynes

Celoten sistem prilagajanja trihodinov na življenje na površini gostitelja je usmerjen v to, da se ne odlepi od gostiteljevega telesa (kar je skoraj vedno enako smrti), hkrati pa ohranja mobilnost. Te naprave so zelo popolne. Telo večine trihodin je v obliki precej ravnega diska, včasih pokrovčka. Stran, ki je obrnjena proti telesu gostitelja, je rahlo konkavna, tvori nastavek za pritrditev. Ob zunanjem robu sesala je venček dobro razvitih ciliatov, s pomočjo katerih se premikanje (plazenje) trepalk v glavnem dogaja po površini telesa ribe. Ta venček ustreza venčku, ki ga najdemo pri sedečih okroglih ciliastih potepuh, o katerih smo govorili zgoraj. Tako lahko trihodino primerjamo s potepuhom. Na trebušni površini (na sesalniku) imajo trihodini zelo zapleteno podporno in pritrdilno napravo, ki pomaga zadržati ciliate na gostitelju. Ne da bi se spuščali v podrobnosti njegove strukture, ugotavljamo, da je njegova osnova obroč kompleksne konfiguracije, sestavljen iz ločenih segmentov, ki nosijo zunanje in notranje zobe (slika 109, B). Ta obroč tvori elastično in hkrati močno osnovo trebušne površine, ki deluje kot sesalo. Različne vrste trihodina se med seboj razlikujejo po številu segmentov, ki tvorijo obroč, ter po konfiguraciji zunanjih in notranjih kavljev.
Na strani telesa trihodine nasproti diska je pernati in ustni aparat. Njegova struktura je bolj ali manj značilna za obrezani ciliati. Adoralne membrane, zavite v smeri urinega kazalca, vodijo do vdolbine, na dnu katere so usta. Jedrski aparat trihodina je običajno urejen za ciliate: eno trakasto makronukleus in eno mikronukleus, ki se nahajata poleg njega. Obstaja ena kontraktilna vakuola.
Trihodini so široko razširjeni v rezervoarjih vseh vrst. Še posebej pogosto jih najdemo na mladičih različnih vrst rib. Pri množičnem razmnoževanju se uporabljajo trihodini velika škoda ribe, še posebej, če mase pokrivajo škrge. To moti normalno dihanje rib.
Za čiščenje rib trihodina je priporočljivo narediti terapevtske kopeli iz 2% raztopine natrijevega klorida ali 0,01% raztopine kalijevega permanganata (za mladice - 10-20 minut).
INFUZORIJ ČREVEGA TRAKTA UNGATES
Iz brazgotine skozi mrežico se hrana podriguje v ustno votlino, kjer jo dodatno žveči (žvečilni gumi). Ponovno pogoltna prežvečena živilska masa skozi posebno cevko, ki jo tvorijo gube požiralnika, ne gre več v brazgotino, ampak v knjigo in od tam v sihod, kjer je izpostavljena prebavnim sokom prežvekovalca. V sirilu v pogojih kisle reakcije in prisotnosti prebavnih encimov ciliati umrejo. Pride tja z žvečilnim gumijem, se prebavijo.
Število protozojev v vampu (pa tudi v mreži) lahko doseže ogromne vrednosti. Če vzamete kapljico vsebine brazgotine in jo pregledate pod mikroskopom (pri segrevanju, saj se ciliati ustavijo pri sobni temperaturi), potem se trebuščki dobesedno rojijo v vidnem polju. Tudi v kulturi je težko pridobiti tako maso ciliatov. Število ciliatov v 1 cm3 vsebine brazgotine doseže milijon in pogosto več. Glede na celotno količino brazgotine to daje resnično astronomske številke! Bogatost vsebine vampa z ciliati je v veliki meri odvisna od narave hrane prežvekovalcev. Če je hrana bogata z vlakninami, revna pa z ogljikovimi hidrati in beljakovinami (trava, slama), potem je v vampu relativno malo ciliatov. Ko se prehrani dodajo ogljikovi hidrati in beljakovine (otrobi), se število ciliatov močno poveča in doseže ogromno. Upoštevati je treba, da obstaja stalen odtok ciliatov. Če se z žvečilnim gumijem združijo v sibič, umrejo. Visoka stopnjaštevilo ciliatov se vzdržuje z njihovim močnim razmnoževanjem.
V prebavnem traktu imajo tudi lihoprsti kopitarji (konj, osel, zebra) veliko število ciliatov, vendar je njihova lokalizacija v gostitelju drugačna. Lihoprsti kopitarji nimajo zapletenega želodca, zaradi česar ni možnosti za razvoj protozojev v sprednjih predelih prebavnega trakta. Toda pri kopitarjih sta zelo dobro razvita velika in slepo kost, ki sta običajno zamašena s prehranskimi masami in igrata bistveno vlogo pri prebavi. V tem delu črevesja se, tako kot v vampu in mreži prežvekovalcev, razvija zelo bogata favna praživali, predvsem ciliatov, ki večinoma spadajo tudi v red endodinomorfov. Po vrstni sestavi pa favna vampa prežvekovalcev in favna debelega črevesa kopitarjev ne sovpadata.
CILITICI ČREVECA PREŽVEKOVALCEV
Najbolj zanimivi so ciliati družina ofrioskolecidov(Ophryoscolecidae), sorodna z red endodiniomorf. Značilna lastnost tega odcepa je odsotnost neprekinjenega ciliarnega pokrova. Kompleksne ciliarne tvorbe - cirresi - se nahajajo na sprednjem koncu telesa ciliatov v predelu ustnih odprtin. Tem osnovnim elementom ciliarnega aparata lahko dodamo dodatne skupine cire, ki se nahajajo bodisi na sprednjem ali zadnjem koncu telesa. Skupno število vrst ciliatov iz družine ofrioscolecid je približno 120.

Slika 110 prikazuje nekaj najbolj značilnih ofrioskolecidov iz vampa prežvekovalcev. Ciliati iz rodu Entodinium (Entodinium, slika 110, L) so najbolj preprosto razporejeni. Na sprednjem koncu njihovega telesa je ena perioralna cona cirusa. Sprednji del telesa, na katerem je odprtina za usta, se lahko potegne navznoter. Ektoplazma in endoplazma sta močno razmejeni. Na zadnjem koncu je jasno vidna analna cev, ki služi za odstranjevanje neprebavljenih ostankov hrane. Nekoliko bolj zapletena struktura anoplodinija(Anoplodinium, slika 110, B). Imajo dve coni ciliarnega aparata - perioralni ciri in dorzalni ciri. Oba se nahajata na sprednjem koncu. Na zadnjem koncu telesa vrste, prikazane na sliki, so dolgi ostri izrastki - to je precej značilno za številne vrste ofrioskolecidov. Domnevajo, da ti izrastki prispevajo k "potiskanju" trepalk med rastlinskimi delci, ki zapolnijo brazgotino.
Vrste Rod Eudiplodynia(Eudiplodinium, sl. 110, B) so podobni anoplodinija, vendar imajo za razliko od njih skeletno osnovno ploščo, ki se nahaja na desnem robu vzdolž žrela. Ta skeletna plošča je sestavljena iz snovi, podobne v kemična narava na vlakna, torej na snov, ki sestavlja lupine rastlinskih celic.
Pri rod poliplastron(Polyplastron, slika 110, D, E) pride do nadaljnjega zapleta okostja. Struktura teh ciliatov je blizu eudiplodinije. Razlike se nanašajo predvsem na dejstvo, da imajo ti ciliati namesto ene skeletne plošče pet. Dva od njih, največja, se nahajata na desni strani, trije, manjše, pa so na levi strani ciliata. Druga značilnost poliplastrona je povečanje števila kontraktilnih vakuol. Entodinija ima eno kontraktilno vakuolo, anoplodinija in eudiplodinija imata dve kontraktilni vakuoli, poliplastron pa jih ima približno ducat.
Pri epidinium(Epidinium, slika 110), ki imajo dobro razvit ogljikohidratni skelet, ki se nahaja na desni strani telesa, se hrbtna cona cirusa premakne s sprednjega konca na hrbtno stran. Bodice se pogosto razvijejo na zadnjem koncu ciliatov tega rodu.
Najkompleksnejša struktura razkriva rod Ofrioscolex(Ophryoscolex), po katerem je poimenovana celotna družina ciliatov (slika 110, E). Imajo dobro razvito hrbtno cono cirusa, ki pokriva približno 2/3 oboda telesa in skeletnih plošč. Na zadnjem koncu se oblikujejo številne bodice, od katerih je ena običajno še posebej dolga.
Spoznavanje nekaterih tipičnih predstavnikov ofrioskolecid kaže, da je znotraj te družine prišlo do pomembnega zapleta organizacije (od entodinije do ofrioskolexa).
Poleg ciliatov družina ofrioskolecidov, v vampu prežvekovalcev se v majhnih količinah nahajajo predstavniki nam že znanih. odcepitev izociliarnih ciliatov. Zastopane so z majhnim številom vrst. Njihovo telo je enakomerno pokrito z vzdolžnimi vrstami cilij, skeletni elementi so odsotni. V skupni masi populacije ciliatov vampa nimajo pomembne vloge, zato se o njih tukaj ne bomo zadrževali.
Kaj in kako jedo ciliati ofrioscolecides? To vprašanje so podrobno preučevali številni znanstveniki, zlasti profesor V. A. Dogel.

Ofrioskolecidna hrana je precej raznolika, pri različnih vrstah pa opazimo določeno specializacijo. Najmanjše vrste iz rodu Entodynia se prehranjujejo z bakterijami, škrobnimi zrni, glivami in drugimi majhnimi delci. Zelo veliko srednjih in velikih ofrioskolecidov absorbira delce rastlinskih tkiv, ki predstavljajo večino vsebine vampa. Endoplazma nekaterih vrst je dobesedno zamašena z rastlinskimi delci. Vidite lahko, kako se ciliati naslanjajo na ostanke rastlinskih tkiv, jih dobesedno raztrgajo na koščke in jih nato pogoltnejo, pri čemer jih v telesu pogosto zvijajo v spiralo (slika 111, 4). Včasih je treba opazovati takšne slike (slika 111, 2), ko je telo samega ciliata deformirano zaradi pogoltnjenih velikih delcev.
Pri ofrioskolecidu včasih opazimo plenjenje. Večje vrste požrejo manjše. Plenilstvo (slika 112) je združeno s sposobnostjo iste vrste, da se prehranjuje z rastlinskimi delci.

Kako ciliati pridejo v vamp prežvekovalca? Kakšne so poti okužbe z ofrioskolecidi? Izkazalo se je, da novorojeni prežvekovalci še nimajo ciliatov v vampu. Prav tako so odsotni, medtem ko se žival hrani z mlekom. Toda takoj, ko prežvekovalec preide na rastlinsko hrano, se v brazgotini in mrežici takoj pojavijo ciliati, katerih število hitro narašča. od kod prihajajo? Dolgo časa se je domnevalo, da ciliati vampa tvorijo nekakšne faze mirovanja (najverjetneje ciste), ki so po naravi zelo razpršene in ob zaužitju povzročijo aktivne faze ciliatov. Nadaljnje študije so pokazale, da ciliati prežvekovalcev nimajo faz mirovanja. Možno je bilo dokazati, da pride do okužbe z aktivnimi gibljivimi ciliati, ki prodrejo v ustno votlino pri rigiranju žlička. Če pregledate pod mikroskopom, vzeto iz ustne votlinežvečilni gumi, vendar ima vedno veliko število aktivno plavajočih ciliatov. Te aktivne oblike zlahka prodrejo v usta in naprej v vamp drugih prežvekovalcev iz skupne posode za pitje, skupaj s travo, senom (ki lahko vsebuje slino z ciliati) itd. Ta pot okužbe je eksperimentalno dokazana.
Če pri ofrioskolecidih ni faz mirovanja, je očitno enostavno pridobiti živali "brez infuzorja" tako, da jih izoliramo, ko se še hranijo z mlekom. Če se izognemo neposrednemu stiku med rastočimi mladiči in ciliastimi prežvekovalci, lahko mlade živali ostanejo brez ciliatov v vampu. Takšne poskuse je izvedlo več znanstvenikov v različnih državah. Rezultat je bil jasen. V odsotnosti stika med mladiči (odvzetimi od matere v obdobju hranjenja z mlekom) in prežvekovalci z ciliati v vampu, živali odraščajo sterilne v primerjavi z ciliati. Vendar pa je že kratkotrajen stik z živalmi z ciliati (skupna krmnica, navadno vedro za pitje, skupni pašnik) dovolj, da se trebuščki pojavijo v vampu sterilnih živali.
Zgoraj so bili rezultati poskusov o vsebnosti prežvekovalcev, popolnoma brez ciliatov v vampu in mreži. To dosežemo, kot smo videli, z zgodnjo izolacijo mladih. Poskusi so bili izvedeni na ovcah in kozah.
Na ta način je bilo mogoče izvajati opazovanja živali "brez infuzorjev" v daljšem časovnem obdobju (več kot eno leto). Kako odsotnost ciliatov v vampu vpliva na življenje gostitelja? Ali odsotnost ciliatov vpliva na gostitelja negativno ali pozitivno? Za odgovor na to vprašanje so bili na kozah izvedeni naslednji poskusi. Dvojčka (istega legla in istega spola) smo vzeli, da bi imeli več podobnega materiala. Nato je bil eden od dvojčkov tega para vzgojen brez ciliatov v vampu (zgodnja izolacija), drugi pa je bil že od samega začetka hranjenja z rastlinsko hrano obilno okužen s številnimi vrstami ciliatov. Oba sta imela popolnoma enako prehrano in sta bila vzgojena v enakih razmerah. Edina razlika med njimi je bila prisotnost ali odsotnost ciliatov. Pri več parih tako preučenih kozličkov ni bilo ugotovljenih razlik v poteku razvoja obeh članov vsakega para (»infuzor« in »neinfuzor«). Tako je mogoče trditi, da ciliati, ki živijo v vampu in mreži, nimajo ostrega vpliva na vitalne funkcije živali gostiteljice.
Zgornji rezultati poskusov pa ne dovoljujejo trditve, da so ciliati vampa popolnoma brezbrižni do lastnika. Ti poskusi so bili izvedeni z običajno prehrano gostitelja. Možno je, da bo v drugih pogojih z drugačno prehrano (na primer z nezadostno prehrano) mogoče razkriti učinek na gostitelja favne protozojev, ki naseljujejo vamp.
V literaturi so podani različni predlogi o možnem pozitivnem vplivu protozojske favne vampa na prebavne procese gostitelja. Poudarjeno je bilo, da številni milijoni ciliatov, ki aktivno plavajo v vampu in drobijo rastlinska tkiva, prispevajo k fermentaciji in prebavi živilskih mas, ki se nahajajo v sprednjih delih prebavnega trakta. Znatno število ciliatov, ki vstopijo v sibuh skupaj z žvečilnim gumijem, se prebavi in absorbira beljakovine, ki sestavljajo pomemben del telesa ciliatov. Infuzorija je torej lahko dodaten vir beljakovin za gostitelja. Predlagano je tudi, da ciliati prispevajo k prebavi vlaknin, ki predstavljajo večino hrane za prežvekovalce, in njeni prenos v bolj prebavljivo stanje.
Vse te domneve niso dokazane, nekaterim pa so bili podani ugovori. Poudarjeno je bilo na primer, da ciliati gradijo protoplazmo svojega telesa iz beljakovin, ki vstopijo v vamp s hrano gostitelja. Absorbirajo rastlinske beljakovine, jih pretvorijo v živalske beljakovine svojega telesa, ki se nato prebavijo v sibuhu. Ali to prinaša kakršno koli korist gostitelju, ostaja nejasno. Vsa ta vprašanja so zelo praktična, saj govorimo o prebavi prežvekovalcev - glavnih predmetov živinoreje. Nadaljnje raziskave o vlogi vampnih ciliatov pri prebavi prežvekovalcev so zelo zaželene.
Ofrioskolecidi prežvekovalcev imajo praviloma široko specifičnost. Po vrstah sta si populacija brazgotine in mreže goveda, ovac in koz zelo blizu. Če primerjamo vrstno sestavo vampa afriških antilop z govedom, potem bo tudi tukaj približno 40 % skupnega števila vrst običajnih. Vendar pa obstaja veliko vrst ofrioskolecidov, ki jih najdemo le v antilopah ali samo pri jelenih. Tako lahko v ozadju splošne široke specifičnosti ofrioskolecidov govorimo o njihovih ločenih, ožje specifičnih vrstah.
Infuzorije črevesja kopitarjev
Zdaj pa se obrnimo na kratko seznanitev s trepalkami, ki naseljujejo veliko in slepo črevo kopitarjev.
Tudi ta živalski svet je vrstno, tako kot favna vampa prežvekovalcev, zelo raznolik. Trenutno je opisanih približno 100 vrst ciliatov, ki živijo v debelem črevesu živali iz družine kopitarjev. Tu najdeni ciliati so v smislu pripadnosti različnim sistematičnim skupinam bolj raznoliki od trepalk v vampu prežvekovalcev.

V črevesju konj je kar nekaj vrst ciliatov, ki sodijo v red izociliarnih, torej ciliatov, pri katerih ciliarni aparat ne tvori membranele ali ciroze v bližini ustne cone (slika 113, 1).
Red entodyniomorph(Entodiniomorpha) je bogato zastopan tudi v črevesju konja. Medtem ko v vampu prežvekovalcev najdemo samo eno družino endodiniomorfov (družina ofrioscolecid), v črevesju konja živijo predstavniki treh družin, na katerih značilnostih se tukaj ne bomo zadrževali in se omejili le na nekaj risb tipične vrste konj (slika 113) .
Podrobne študije A. Strelkova so pokazale, da različne vrste ciliatov še zdaleč niso enakomerno razporejene po debelem črevesu konja. Obstajata dve različni skupini vrst, dve favni, tako rekoč. Eden od njih naseljuje slepo črevo in trebušni del debelega črevesa (začetni odseki debelega črevesa), drugi pa dorzalni del debelega in tankega črevesa. Ta dva kompleksa vrst sta precej ostro razmejena. Za ta dva oddelka je nekaj skupnih vrst - manj kot ducat.
, 
Zanimivo je, da med številnimi vrstami ciliatov, ki naseljujejo debelo črevo kopitarjev, obstajajo predstavniki istega rodu, ki so povezani s sesajočimi trepalkami. Kot smo videli zgoraj, sesanje ciliatov(Suctoria) so tipični prostoživeči sedeči organizmi s prav posebnim načinom prehranjevanja s pomočjo lovk (slika 103). Eden od porodni suktor prilagojeno tako na videz nenavadnemu habitatu, kot je debelo črevo konja, na primer več vrst alantoza(Alantosom). Te zelo svojevrstne živali (slika 114) nimajo peclja, cilije so odsotne, paličaste lovke, ki so odebeljene na koncih, so dobro razvite.
S pomočjo lovk se alantosomi prilepijo na različne vrste ciliatov in jih izsesajo. Pogosto je plen večkrat večji od plenilca.
Vprašanje narave razmerja med ciliati debelega črevesa kopitarjev in njihovimi gostitelji je še vedno nejasno. Število ciliatov je lahko tako veliko, včasih celo več kot v vampu prežvekovalcev. Obstajajo podatki, ki kažejo, da lahko količina ciliatov v debelem črevesu konja doseže 3 milijone v 1 cm3. Simbiotični pomen, ki ga predlagajo nekateri znanstveniki, je še manj verjeten kot pri vampnih trepalkah.
Najverjetneje je mnenje, da povzročajo nekaj škode gostitelju, saj absorbirajo veliko količino hrane. Del ciliatov se izloči s fekalnimi masami in tako organske snovi (vključno z beljakovinami), ki sestavljajo njihovo telo, ostanejo neuporabljene s strani gostitelja.
Vprašanje o načinih okužbe kopitarjev z ciliati, ki naseljujejo debelo črevo, še ni rešeno.

Balantidia zajame različne delce hrane iz vsebine debelega črevesa. Še posebej voljno se prehranjuje s škrobnimi zrni. Če balantidija živi v lumnu človeškega debelega črevesa, se prehranjuje z vsebino črevesja in nima nobenega škodljivega učinka. To je tipičen "kočija", ki smo ga srečali že pri obravnavi dizenterijske amebe. Vendar je manj verjetno, da bo balantidija kot dizenterična ameba ostala tako "neškodljiv najemnik".
Trenutno so se strokovnjaki dobro razvili različne metode, ki omogoča gojenje balantidije v umetnem okolju - zunaj gostiteljskega organizma.

Kot je razvidno iz slike, je troglodptella eden od kompleksnih endodiniomorfov. Poleg perioralne cone ciroze (na sprednjem koncu telesa) ima še tri cone dobro razvitega ciroze, ki obročasto pokrivajo telo ciliata. Trogloditeli imajo dobro razvit skeletni aparat, sestavljen iz ogljikovih hidratov, ki pokriva skoraj celoten sprednji konec telesa. Velikosti teh nenavadnih ciliatov so precej pomembne. V dolžino dosežejo 200-280 mikronov.
Ciliates brez ust ASTOMATS

Podporne skeletne tvorbe se razvijejo predvsem na sprednjem koncu telesa, ki mora doživeti mehanske obremenitve in premagovati ovire, ki se prebijajo skozi črevesni lumen med delci hrane. Vrste rod radiofrija(Radiophrya) na sprednjem koncu na eni strani telesa (ki se običajno šteje za trebušno stran) so zelo močna elastična rebra (spikule), ki ležijo v površinski plasti ektoplazme (slika 117, B, D, E) . Vrste Rod Menilella(Mesnilella) obstajajo tudi podporni žarki (spikule), ki večino svoje dolžine ležijo v globljih plasteh citoplazme (v endoplazmi, sl. 117, A). Podobno urejene nosilne formacije so razvite tudi pri vrstah nekaterih drugih rodov Astomat.
Nespolno razmnoževanje pri nekaterih astomatih ciliatov poteka na svojstven način. Namesto prečne delitve na dva, ki je značilna za večino ciliatov, ima veliko astomatov neenakomerno delitev (brstenje). Hkrati ledvice, ki se ločijo na zadnjem koncu, še nekaj časa ostanejo povezane z materinim posameznikom (slika 117, B). Kot rezultat dobimo verige, ki jih sestavljajo sprednji veliki in zadnji manjši posamezniki (ledvice). V prihodnosti se ledvice postopoma ločijo od verige in preidejo v samostojen obstoj. Ta posebna oblika razmnoževanja je razširjena, na primer, pri nam že znani radiofriji. Verige nekaterih astomatov, ki so posledica brstenja, so podobne videz verige trakulj. Tu se spet srečamo s pojavom konvergence.
Jedrski aparat astomata ima strukturo, značilno za ciliate: makronukleus, najpogosteje trakaste oblike (slika 117), in eno mikronukleus. Kontraktilne vakuole so običajno dobro razvite. Večina vrst ima več (včasih več kot ducat) kontraktilnih vakuol, razporejenih v eni vzdolžni vrsti.
Študija porazdelitve vrst Astomat po različnih vrstah gostiteljev kaže, da je večina vrst Astomat omejena na strogo določene vrste gostiteljev. Za večino stomatov je značilna ozka specifičnost: le ena živalska vrsta jim lahko služi kot gostitelj.
Kljub velikemu številu študij, posvečenih preučevanju astomatskih ciliatov, ostaja en zelo pomemben vidik njihove biologije popolnoma nejasen: kako poteka prenos teh ciliatov z enega gostiteljskega posameznika na drugega? Nikoli ni bilo mogoče opaziti nastanka cist pri teh ciliatih.
Zato se predlaga, da se okužba pojavlja aktivno - mobilne faze.
INFUZORIJ ČREVEVA MORSKEGA JEŽKA
Morski ježki so zelo številni v obalnem območju naših severnih (Barents) in Daljnega vzhoda (Japonsko morje, pacifiška obala Kurilskih otokov). večina morski ježki prehranjuje se z rastlinsko hrano, predvsem z algami, ki jih s posebnimi ostrimi »zobčki«, ki obdajajo ustno odprtino, postrgajo s podvodnih predmetov. V črevesju teh rastlinojedih ježev je bogata favna ciliatov. Pogosto se tukaj razvijejo v množičnih količinah, vsebina črevesja morskega ježka pod mikroskopom pa je skoraj tako »kipiva« z infuzorijami kot vsebina vampa prežvekovalcev. Povedati je treba, da poleg globokih razlik v življenjskih razmerah ciliatov črevesja morskega ježka in vampa prežvekovalcev obstaja nekaj podobnosti. Ležijo v tem, da tako tu in tam ciliati živijo v okolju, ki je zelo bogato z rastlinskimi ostanki. Trenutno je znano, da v črevesju morskih ježkov živi več kot 50 vrst ciliatov, ki jih najdemo le v obalnem območju, kjer se ježki hranijo z algami. Na velikih globinah, kjer alge ne rastejo več, v morskih ježkih ni ciliatov.

Glede na način življenja in naravo prehrane je večina ciliatov v črevesju morskih ježkov rastlinojedih. Hranijo se z algami, ki v velikih količinah napolnijo črevesje gostitelja. Nekatere vrste so pri izbiri hrane precej "izbirčne". na primer strobilidij(Strobilidium, sl. 118, A) se prehranjuje skoraj izključno z velikimi diatomeji. Obstajajo tudi plenilci, ki jedo predstavnike drugih, več majhne vrste ciliatov.
Pri ciliatih iz črevesja morskih ježkov, za razliko od astomata, ni stroge omejitve na določene vrste gostiteljev. Živijo v najrazličnejših morskih ježkih, ki se hranijo z algami.
Načini okužbe morskih ježkov z ciliati niso raziskani. Vendar pa je tukaj z visoko stopnjo verjetnosti mogoče domnevati, da se pojavlja v aktivnih prosto plavajočih oblikah. Dejstvo je, da lahko ciliati iz črevesja morskih ježkov živijo dolgo (več ur) v morski vodi. Vendar so se že tako zelo prilagodili življenju v črevesju ježkov, da zunaj njihovega telesa, v morski vodi, prej ali slej poginejo.
Ob zaključku seznanitve z ciliati je treba še enkrat poudariti, da predstavljajo z vrstami bogato, obsežno in uspešno skupino (razred) živalskega sveta. Infuzorije, ki so ostale na ravni celične organizacije, so v primerjavi z drugimi razredi protozojev dosegle največjo kompleksnost strukture in funkcij.
Posebno pomembno vlogo pri tem progresivnem razvoju (evoluciji) sta verjetno odigrala transformacija jedrskega aparata in nastanek jedrskega dualizma (kvalitativne neekvivalence jeder). Bogastvo makronukleusa z nukleinskimi snovmi je povezano z aktivnimi presnovnimi procesi, z živahnimi procesi sinteze beljakovin v citoplazmi in jedrih.
Zaključek
Prišli smo do konca našega pregleda strukture in življenjskega sloga ekstenzivne vrste živalskega sveta - protozoji. Njihova značilnost, kot je bilo že večkrat poudarjeno zgoraj, je enoceličnost. Po svoji strukturi so protozoji celice. Vendar pa so neprimerljive s celicami, ki sestavljajo telo večceličnih organizmov, saj so sami organizmi. Tako so protozoji organizmi na celični ravni organizacije. Zdi se, da nekatere visoko organizirane protozoje, ki imajo veliko jeder, že presegajo morfološke meje celične strukture, kar nekaterim znanstvenikom daje razlog, da takšne protozoje imenujejo "nadcelične". To malo spremeni bistvo zadeve, saj je enocelična organizacija še vedno značilna za protozoe.
V mejah enoceličnosti so protozoji prehodili dolgo pot evolucijskega razvoja in dali ogromno različnih oblik, prilagojenih najrazličnejšim življenjskim razmeram. V središču rodovniškega stebla protozojev sta dva razreda: sarcodes in flagellats. O vprašanju, kateri od teh razredov je bolj primitiven, se v znanosti še vedno razpravlja. Po eni strani imajo nižji predstavniki Sarcodidae (ameba) najbolj primitivno strukturo. Toda flagelati kažejo največjo plastičnost vrste metabolizma in stojijo tako rekoč na meji med živalskim in rastlinskim svetom. V življenjskem ciklu nekaterih sarkodov (na primer foraminifer) obstajajo stopnje biča (gamete), kar kaže na njihovo razmerje z bičevci. Očitno je, da niti moderni sarkodi niti moderni bičevci ne morejo biti začetna skupina evolucije živalskega sveta, saj so sami prehodili dolgo pot v zgodovinskem razvoju in razvili številne prilagoditve sodobnim življenjskim razmeram na Zemlji. Verjetno je treba oba razreda sodobnih protozojev obravnavati kot dva debla v evoluciji, ki izvirata iz starodavnih oblik, ki niso preživele do danes, ki so živele na zori razvoja življenja na našem planetu.
V nadaljnji evoluciji protozojev je prišlo do sprememb drugačne narave. Nekatere so privedle do splošnega dviga nivoja organiziranosti, povečanja aktivnosti in intenzivnosti življenjskih procesov. Med takšne filogenske (evolucijske) preobrazbe je treba na primer uvrstiti razvoj organelov gibanja in zajemanja hrane, ki je v razredu ciliatov dosegel visoko stopnjo popolnosti. Nesporno je, da so cilije organele, ki ustrezajo (homolognim) flagelam. Medtem ko je pri bičačih, z nekaj izjemami, število bičk malo, je pri ciliatih število cilijev doseglo več tisoč. Razvoj ciliarnega aparata je močno povečal aktivnost protozojev, naredil je oblike njihovega odnosa z okoljem bolj raznolike in zapletene, oblike reakcij na zunanje dražljaje. Prisotnost diferenciranega ciliarnega aparata je bil nedvomno eden od glavnih razlogov za postopno evolucijo v razredu ciliatov, kjer je nastala široka paleta oblik, prilagojenih različnim habitatom.
Razvoj ciliarnega aparata ciliatov je primer tovrstne evolucijske spremembe, ki jo je poimenoval akad. Severcove aromorfoze. Za aromorfozo je značilno splošno povečanje organizacije, razvoj prilagoditev širokega pomena. Povečanje organizacije se razume kot spremembe, ki povzročajo povečanje vitalne aktivnosti organizma; povezani so s funkcionalno diferenciacijo njegovih delov, kar vodi do bolj raznolikih oblik komunikacije med organizmom in okoljem. Razvoj ciliarnega aparata ciliatov se nanaša prav na tovrstne strukturne preobrazbe v procesu evolucije. To je tipična aromorfoza.
Pri protozojih, kot je poudaril V. A. Dogel, so spremembe v vrsti aromorfoz običajno povezane s povečanjem števila organelov. Pojavi se polimerizacija organelov. Razvoj ciliarnega aparata pri ciliatih je tipičen primer takšnih sprememb. Drugi primer aromorfoze pri evoluciji ciliatov je njihov jedrski aparat. Zgoraj smo preučili strukturne značilnosti jedra ciliatov. Jedrski dualizem ciliatov (prisotnost mikronukleusa in makronukleusa) je spremljalo povečanje števila kromosomov v makronukleusu (fenomen poliploidije). Ker so kromosomi povezani z glavnimi sintetičnimi procesi v celici, predvsem s sintezo beljakovin, je ta proces privedel do splošnega povečanja intenzivnosti glavnih vitalnih funkcij. Tudi tu je potekala polimerizacija, ki je vplivala na kromosomske komplekse jedra.
ciliatov- ena najštevilčnejših in naprednih skupin protozojev, izvira iz bičačev. To dokazuje popolna morfološka podobnost njihovih organoidov gibanja. Ta stopnja evolucije je bila povezana z dvema velikima aromorfozama: ena od njih je vplivala na organele gibanja, druga - na jedrski aparat. Obe vrsti sprememb sta medsebojno povezani, saj obe vodita k povečanju vitalne aktivnosti in zapletu oblik medsebojnih odnosov z zunanjim okoljem.
Poleg aromorfoza obstaja še ena vrsta evolucijske spremembe, ki se izraža v razvoju prilagoditev (prilagajanja) na določene, strogo določene pogoje obstoja. To vrsto evolucijske spremembe je Severtsov poimenoval idioadaptacija. V evoluciji protozojev je ta vrsta sprememb igrala zelo pomembno vlogo. Zgoraj, ko razmišljate različni razredi protozoa, podani so številni primeri idioadaptivnih sprememb. Prilagoditve na planktonski način življenja v različnih skupinah protozojev, prilagoditve na življenje v pesku pri ciliatih, nastanek zaščitnih lupin oocist v kokcidijah in še veliko več - vse to so idioadaptacije, ki so imele veliko vlogo pri nastanku in razvoju. posameznih skupin, niso pa povezane s splošnimi progresivnimi spremembami v organizaciji.
Prilagoditve na različne specifične habitate pri protozojih so zelo raznolike. Zagotovili so široko razširjenost te vrste v najrazličnejših habitatih, kar je bilo podrobneje obravnavano zgoraj, pri opisu posameznih razredov.
Velika medicinska enciklopedija
Infuzoria Tetrahymena thermophila ... Wikipedia
- (Infusoria) razred najbolj razvitih protozojev (Protozoa). Glavne značilnosti I.: prisotnost cilij (za gibanje in prehrano), dve vrsti jeder (poliploidni makronukleus in diploidni mikronukleus, različni po strukturi in ... ... Velika sovjetska enciklopedija
Ali tekoče živali (Infusoria) razred protozojskih živali, opremljenih s svetlečimi pletenicami ali cilijami, ki jih v odrasli dobi včasih nadomestijo cilindrični sesalni izrastki, z določeno obliko telesa, večinoma z ... ... enciklopedični slovar F. Brockhaus in I.A. Efron
Taksonomija ifuzorijev razvrstitev in sistematizacija organizmov, ki pripadajo vrsti Ciliates, v skladu s splošnim konceptom taksonomije zlasti protozojev in živali na splošno Vsebina 1 Sistematika 1.1 Klasična ... Wikipedia
Ali tekoče živali (Infusoria) razred protozojskih živali, opremljenih s svetlečimi pletenicami ali cilijami, ki jih v odraslem stanju včasih nadomestijo valjasti sesalni izrastki, z določeno obliko telesa, večinoma z ... ... Enciklopedija Brockhausa in Efrona
Vrsta Ciliates se običajno imenuje tudi Ciliated - organi gibanja teh protozojev so cilije. Ciliatna celica ima dve jedri, imenujemo jih majhna in velika. Prvi uravnava proces razmnoževanja, drugi pa je odgovoren za procese prehrane, gibanja in dihanja.
Značilnosti življenja te vrste je treba upoštevati na primeru ciliates-čevljev.
Gibanje in dihanje
Infuzorija-čevelj, katerega dolžina je približno 0,5 mm, si za svoj življenjski prostor izbere vodna telesa. Obliko telesa najpreprostejših je enostavno uganiti po imenu - spominja na čevelj. Hitrost gibanja je približno 2,5 mm na sekundo.
Prisotnost zunanje elastične lupine zagotavlja stabilno obliko telesa.
V citoplazmi, ki meji na lupino, so podporna vlakna, njihov razvoj je zagotovilo za ohranitev stalne oblike ciliata.
Na površini ciliatov je 15 tisoč ciliatov, na njihovem dnu je bazalno telo. Gibanje poteka s pomočjo tresljajev cilijev: proizvedejo približno 30 udarcev na sekundo, s čimer potisnejo ciliat-čevelj naprej.
Diha skozi površino telesa.
Prehrana
Značilnost ciliatov je prisotnost celična usta, v bližini katerega so še posebej dolge in goste cilije. Celična usta se nadaljujejo s celičnim žrelom: cilije potiskajo vodo in ciliat hrano - bakterije vanj.
Infuzorija zaznava kemikalije, kar sprošča kopico bakterij. Tako išče plen.
Hrana nato konča v prebavni vakuoli, kjer se prebavi. Od tu sledi že v citoplazmo.
Izbor
Izbira se izvaja z uporabo dve kontraktilni vakuoli, eden se nahaja na sprednjem koncu, drugi pa na zadnji strani. Vakuole so sestavljene iz rezervoarja in kanalov.
Tekočina napolni kanale, nato sledi osrednjemu rezervoarju, nato pa zapusti infuzorijo. Proces krčenja vakuol traja 10-20 sekund.
razmnoževanje
Infuzorija se razmnožuje nespolno - razdeljena je na dva dela. Njegova značilnost je razdelitev po telesu.
Jedra ciliatov so razdeljena na dva dela: novonastali ciliati imajo majhno in veliko jedro. Hčerinski ciliati imajo dele organoidov, manjkajoči pa se oblikujejo samostojno. Razmnoževanje se pojavi večkrat na dan.
Za trebušne čevlje je možno tudi spolno razmnoževanje, vendar se v tem primeru število posameznikov ne poveča. Začasno so protozoji povezani in tvorijo povezovalni most iz citoplazme.
Pri vsakem posamezniku veliko jedro izgine, majhna pa se dvakrat razdelijo - pojavijo se štiri jedra. Od teh ostane samo eno jedro, ki se tudi deli. Posameznik vsebuje dve jedri, nato pride do izmenjave jeder - eno od jeder se premakne k drugemu posamezniku.
Tam se zlije z jedrom, ki ostane, in tako v vsakem od posameznikov nastane manjše in večje jedro. Ta proces, imenovan konjugacija, je nujen za obnovo genskega materiala med posamezniki.
Vrste ciliatov
Ciliati so zapleteno organizirane protozoje, jih je približno 7000 vrst.
1. Raznolikost sarkodov.
2. Raznolikost flagel.
Predstavniki drugega podtipa se premikajo s pomočjo flagel, kar se odraža v imenu te skupine.
- Flagellate. Euglena zelena je tipičen predstavnik te skupine. Ta protozoan ima veretano obliko, prekrito z gosto elastično membrano. Na sprednjem koncu telesa je biček, blizu dna katerega je oko, občutljivo na svetlobo - stigma in kontraktilna vakuola. Bližje zadnjemu koncu, v debelini citoplazme, je jedro, ki nadzoruje vse vitalne procese telesa. Zelena barva euglene je posledica številnih kloroplastov. Na svetlobi ta flagelata fotosintezira, vendar se dolgo časa na neosvetljenih območjih rezervoarja preklopi na hranjenje s produkti razpada kompleksnih organskih snovi in jih izloči iz okolja. Tako lahko zelena euglena združuje tako avtotrofno vrsto presnove kot heterotrofno. Obstoj organizmov s tako mešano vrsto prehrane kaže na razmerje med živalskim in rastlinskim svetom. Večina flagel
3. Raznolikost sporozojev in ciliatov.
Tip Ciliates vključuje protozoje, ki jih odlikuje najbolj zapletena organizacija med enoceličnimi živalmi. Tipičen predstavnik te skupine je infuzorija-čevelj - običajen prebivalec sladke vode. Njeno telo je pokrito z gosto lupino - pelikulo in ima zato relativno konstantno obliko. Značilna lastnost strukture je prisotnost cilij, ki enakomerno pokrivajo celotno telo čevlja. Gibanje cilij je usklajeno zaradi mreže kontraktilnih vlaken, ki se nahajajo v površinski plasti citoplazme. Druga značilnost je prisotnost dveh jeder, velikega (makronukleus) in majhnega (mikronukleus). Jedra se razlikujejo tudi funkcionalno: veliko uravnava presnovo, majhno pa sodeluje pri spolnem procesu (konjugacija). Infuzorija-čevelj se prehranjuje z bakterijami, enoceličnimi algami, ki jih cilije prilagodijo celičnim ustjem, ki ležijo na dnu predustne votline. Po prehodu skozi celično žrelo, ki se konča v citoplazmi, se delci hrane zaprejo v prebavne vakuole, kjer se pod delovanjem encimov razgradijo. Neprebavljeni ostanki se vržejo ven skozi prašek. V telesu čevlja sta dve izmenično pulzirajoči kompleksni kontraktilni vakuoli. Čevlji se razmnožujejo nespolno, z delitvijo na polovico. Večkratno nespolno razmnoževanje se v čevlju nadomesti s spolnim procesom – konjugacijo, med katero se dva čevlja združita in izmenjujeta genetski material. Po tem se čevlji razhajajo in kmalu spet začnejo nespolno razmnoževanje. Biološki pomen konjugacije je v kombinaciji v enem organizmu dednih lastnosti dveh posameznikov. S tem se poveča njegova sposobnost preživetja, ki se izraža v boljši prilagodljivosti okoljskim razmeram.
Niste našli tistega, kar ste iskali? Uporabite iskanje
Na tej strani gradivo o temah:
- na kratko o bičačih, psevdopodih, sporozojih in cilijatih
- sarkodi in sporozoji
- sorta flagel
- povzetek poročila o copatih infuzoria
- kakšna je razlika med ciliati in vrstami sarcode
Eden najbolj tipičnih znanih predstavnikov ciliarja je ciliatni čevelj. Praviloma živi v vodi stoječe smeri, pa tudi v sladkovodnih rezervoarjih, kjer tok odlikuje izjema samozavest. Njegov habitat mora nujno vsebovati razpadajočo organsko snov. Priporočljivo bi bilo podrobno preučiti vse vidike življenja tega predstavnika favne.
Predstavniki trepalnic

Treba je opozoriti, da so Ciliati vrsta, katere ime izvira iz besede "tinktura" (prevedeno iz latinščina). To je mogoče razložiti z dejstvom, da so bili prvi predstavniki protozojev najdeni prav v zeliščnih tinktrah. Sčasoma je razvoj te vrste začel hitro pridobivati zagon. Tako je že danes v biologiji znanih približno 6-7 tisoč vrst, ki vključujejo vrsto Ciliates. Če se zanašamo na podatke iz osemdesetih let prejšnjega stoletja, potem lahko trdimo, da zadevni tip vsebuje dva razreda v svoji strukturi: ciliates ciliates (ima tri nadrede) in sesalni ciliates. V zvezi s temi podatki lahko sklepamo, da je pestrost živih organizmov zelo široka, kar je resnično zanimivo.
Vrsta Infuzorije: predstavniki

Pomembni predstavniki te vrste so ciliates-balantidia in ciliates-čevelj. Posebnosti Te živali so prevleka pelikula z cilijami, ki se uporabljajo za gibanje, zaščito ciliatov s pomočjo posebej zasnovanih organov, trihociste (ki se nahajajo v ektoplazmi lupine), kot tudi prisotnost v celici dveh jeder (vegetativnih). in generativni). Poleg tega ustna votlina na telesu ciliata tvori ustni lijak, ki teži k prehajanju v celična usta, ki vodijo v žrelo. Tam nastanejo prebavne vakuole, ki služijo neposredno za prebavo hrane. Toda neprebavljene komponente se odstranijo iz telesa skozi prašek. Značilnosti vrste ciliatov zelo vsestranski, vendar so glavne točke obravnavane zgoraj. Edino, kar je treba dodati, je, da se ciliata nahajata na nasprotnih delih telesa. Z njihovim delovanjem se iz telesa izloča odvečna voda ali presnovni produkti.
Infuzoria čevelj

Da bi kvalitativno preučili strukturo in način življenja tako zanimivih organizmov enocelične strukture, bi se bilo priporočljivo sklicevati na ustrezen primer. To zahteva čevlje za infuzorije, ki so razširjene v sladkovodnih rezervoarjih. Z lahkoto jih je mogoče gojiti v navadnih posodah (na primer v akvarijih), najlažji je zaliv travniškega sena sladko vodo, ker se v tovrstnih tinkturah praviloma razvije veliko vrst protozojev, vključno z ciliati-čevlji. Tako lahko skozi mikroskop praktično preučite vse informacije, ki so navedene v članku.
Značilnosti ciliates-čevljev
Kot je navedeno zgoraj, so Ciliates vrsta, ki vključuje številne elemente, med katerimi je najbolj zanimiv ciliatni čevelj. To je dolžina, ki je pol milimetra, obdarjena z vretenasto obliko. Treba je opozoriti, da je ta organizem vizualno podoben čevlju, zato ima tako zanimivo ime. Infuzorija-čevelj je nenehno v gibanju in plava s topim koncem naprej. Zanimivo je, da hitrost njegovega gibanja pogosto doseže 2,5 mm na sekundo, kar je zelo dobro za predstavnika te vrste. Na površini telesa ciliatov-čevljev lahko opazimo cilije, ki služijo kot motorični organeli. Kot vsi ciliati ima zadevni organizem v svoji strukturi dve jedri: veliko je odgovorno za prehranske, dihalne, motorične in presnovne procese, majhno pa sodeluje pri spolnem vidiku.
Telo ciliatov-čevljev

Struktura telesa ciliatov-čevljev je zelo zapletena. Zunanja prevleka tega predstavnika je tanka elastična lupina. Sposoben je vzdrževati pravilno obliko telesa telesa skozi vse življenje. Zvesti pomočniki pri tem so odlično razvita podporna vlakna, ki se nahajajo v citoplazmatski plasti, ki je tesno ob membrani. Površina telesa ciliatnega čevlja je obdarjena z ogromnim številom (približno 15.000) cilij, ki nihajo ne glede na zunanje okoliščine. Na dnu vsakega od njih je bazalno telo. Cilije se premikajo približno 30-krat na sekundo, da potisnejo telo naprej. Pomembno je omeniti, da so valovna gibanja teh instrumentov zelo usklajena, kar omogoča, da se infuzorija med gibanjem počasi in lepo vrti okoli vzdolžne osi svojega telesa.
Ciliates - vrsta določenega zanimanja
Za popolno razumevanje vseh značilnosti ciliates-čevljev je priporočljivo upoštevati glavne procese njegovega življenja. Torej gre za uživanje bakterij in alg. Telo organizma je obdarjeno z vdolbino, imenovano celična usta in prehaja v žrelo, na dnu katerega hrana vstopa neposredno v vakuolo. Tam se prebavi približno eno uro, pri čemer pride do prehoda iz kislega v alkalno okolje. Vakuole se premikajo v telesu ciliata skozi tok citoplazme, neprebavljeni ostanki pa gredo v zadnji del telesa skozi prah.
Dihanje ciliatov-čevljev poteka s pomočjo kisika, ki vstopa v citoplazmo skozi kožo telesa. In izločevalni procesi potekajo skozi dve kontraktilni vakuoli. Kar zadeva razdražljivost organizmov, se ciliati-čevlji nagibajo k združevanju v bakterijske komplekse kot odgovor na delovanje snovi, ki jih izločajo bakterije. In odplavajo stran od takega dražilnega sredstva, kot je kuhinjska sol.
razmnoževanje

Ciliat čevlja se lahko razmnožuje na enega od dveh načinov. Bolj razširjeno je nespolno razmnoževanje, po katerem so jedra razdeljena na dva dela. Kot rezultat te operacije vsak ciliat vsebuje 2 jedri (velika in majhna). Spolno razmnoževanje je primerno, če pride do pomanjkanja hranil ali spremembe temperaturnega režima živalskega telesa. Treba je opozoriti, da se po tem lahko infuzorija spremeni v cisto. Toda pri spolnem tipu razmnoževanja je povečanje števila posameznikov izključeno. Torej sta dva ciliata med seboj povezana za določeno časovno obdobje, zaradi česar se lupina raztopi in med živalmi nastane povezovalni most. Pomembno je, da veliko jedro vsakega od njih izgine brez sledu, majhno pa gre skozi proces cepitve dvakrat. Tako v vsakem ciliatu nastanejo 4 hčerinska jedra, po katerih se tri uničijo, četrta pa se ponovno razdeli. Ta spolni proces se imenuje konjugacija. In njegovo trajanje je lahko do 12 ur.
Infuzorija-čevelj živi v majhnih stoječih rezervoarjih. Ta enocelična žival, dolga 0,5 mm, ima vretenasto telo, ki je nejasno podobno čevlju. Ciliati so nenehno v gibanju, plavajo s topim koncem naprej. Hitrost gibanja te živali doseže 2,5 mm na sekundo. Na površini telesa imajo organele gibanja - cilije. V celici sta dve jedri: veliko jedro je odgovorno za prehrano, dihanje, gibanje, presnovo; majhno jedro je vključeno v spolni proces.
Struktura infuzorije za čevlje
Organizem ciliatov je bolj zapleten. Tanka elastična lupina, ki pokriva zunanjost trepalnice, ohranja konstantno obliko svojega telesa. To olajšajo tudi dobro razvite nosilne fibrile, ki se nahajajo v plasti citoplazme, ki meji na lupino. Približno 15.000 nihajočih ciliatov se nahaja na površini telesa ciliata. Na dnu vsake cilije leži bazalno telo. Gibanje vsake trepalnice je sestavljeno iz ostrega giba v eno smer in počasnejšega, bolj gladkega vračanja v prvotni položaj. Cilije vibrirajo približno 30-krat na sekundo in kot vesla potiskajo infuzorijo naprej. Valovito gibanje cilij je usklajeno. Ko ciliat-čevelj plava, se počasi vrti okoli vzdolžne osi telesa.
Življenjski procesi
Prehrana
Slipper in nekateri drugi prostoživeči ciliati se hranijo z bakterijami in algami.
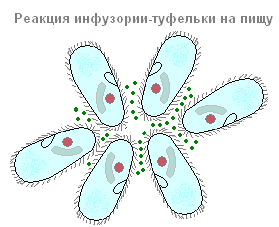
Reakcija ciliates-čevljev na hrano
Tanka elastična lupina, ( celična membrana), ki od zunaj prekriva ciliat, ohranja stalno obliko telesa. Približno 15 tisoč cilij se nahaja na površini telesa. Na telesu je vdolbina - celična usta, ki prehaja v celično žrelo. Na dnu žrela hrana vstopi v prebavno vakuolo. V prebavni vakuoli se hrana prebavi v eni uri, najprej s kislo in nato z alkalno reakcijo. Prebavne vakuole se premikajo v telesu ciliatov s tokom citoplazme. Neprebavljeni ostanki se izvržejo na zadnji del telesa skozi posebno strukturo - prah, ki se nahaja za ustno odprtino.
Dih
Dihanje poteka skozi prevleko telesa. Kisik vstopa v citoplazmo skozi celotno površino telesa in oksidira kompleksne organske snovi, zaradi česar se spremenijo v vodo, ogljikov dioksid in nekatere druge spojine. Hkrati se sprosti energija, ki je potrebna za življenje živali. Ogljikov dioksid se med dihanjem odstrani skozi celotno površino telesa.
Izbor
V telesu ciliatov-čevljev sta dve kontraktilni vakuoli, ki se nahajata na sprednjem in zadnjem koncu telesa. Zbirajo vodo z raztopljenimi snovmi, ki nastanejo pri oksidaciji kompleksnih organskih snovi. Ko dosežejo mejno vrednost, se kontraktilne vakuole približajo površini telesa in njihova vsebina se izlije. Pri sladkovodnih enoceličnih živalih se preko kontraktilnih vakuol odstrani odvečna voda, ki nenehno prihaja v njihovo telo iz okolja.
Razdražljivost
Ciliates-čevlji se zberejo v gruče bakterij kot odgovor na delovanje snovi, ki jih izločajo, vendar plavajo stran od takšnega dražilnega sredstva, kot je kuhinjska sol.
Razdražljivost je lastnost vseh živih organizmov, da se odzivajo na delovanje dražljajev – svetlobe, toplote, vlage, kemikalij, mehanskih vplivov. Zaradi razdražljivosti se enocelične živali izogibajo neugodnim razmeram, najdejo hrano, posameznike svojega leta.
razmnoževanje
aseksualni
Ciliati se običajno razmnožujejo nespolno z delitvijo na dva dela. Jedra so razdeljena na dva dela, vsak nov ciliat pa vsebuje eno veliko in eno majhno jedro. Vsaka od obeh hčera prejme del organelov, druge pa nastanejo na novo.
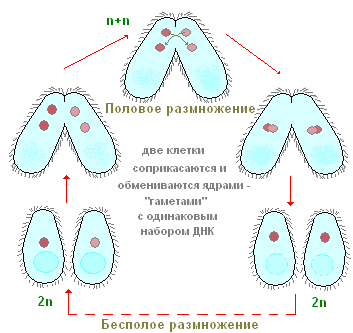
Reprodukcija ciliatov-čevljev
spolne
Ob pomanjkanju hrane ali spremembi temperature ciliati gredo na spolno razmnoževanje, nato pa se lahko spremenijo v cisto.
Med spolnim procesom se število posameznikov ne poveča. Dva ciliata sta med seboj začasno povezana. Na mestu stika se lupina raztopi in med živalmi nastane povezovalni most. Veliko jedro vsakega ciliata izgine. Majhno jedro se dvakrat razdeli. V vsakem ciliatu nastanejo štiri hčerinska jedra. Trije so uničeni, četrti pa je spet razdeljen. Posledično v vsakem ostaneta dve jedri. Jedrska izmenjava poteka vzdolž citoplazemskega mostu in se tam zlije s preostalim jedrom. Novonastala jedra tvorijo veliko in majhno jedro, ciliati pa se razhajajo. Ta spolni proces se imenuje konjugacija. Traja približno 12 ur. Spolni proces vodi v obnovo, izmenjavo med posamezniki in prerazporeditev dednega (genetskega) materiala, kar poveča sposobnost preživetja organizmov.
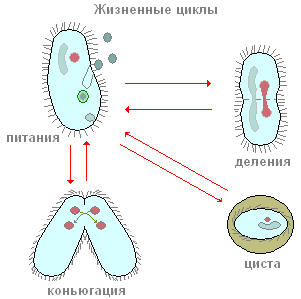
Življenjski cikel ciliatov-čevljev
Priporočamo tudi
 Stikalno napajanje: popravilo in izboljšanje
Stikalno napajanje: popravilo in izboljšanje
 Daljinski nadzor svetlobe
Daljinski nadzor svetlobe
 Učne ure plavanja za predšolske otroke
Učne ure plavanja za predšolske otroke
 Opombe za mojstra - domači gospodinjski alarmi
Opombe za mojstra - domači gospodinjski alarmi
 Propeler ure na Atmega8
Propeler ure na Atmega8
 Primeri uporabe naprav in relejev, kako izbrati in pravilno povezati rele Mikrokrmilnik in rele preprosta preklopna vezja
Primeri uporabe naprav in relejev, kako izbrati in pravilno povezati rele Mikrokrmilnik in rele preprosta preklopna vezja