숙제. 인퓨소리아 슬리퍼
다른 원생 동물 그룹과 비교할 때 섬모는 기능의 다양성과 복잡성과 관련된 가장 복잡한 구조를 가지고 있습니다.
"인퓨소리아 슬리퍼"라는 이름은 어디에서 왔습니까? 현미경으로 살아있는 섬모나 그 이미지를 보면 놀라지 않을 것입니다(그림 85).
실제로이 섬모의 몸 모양은 우아한 숙녀 신발과 비슷합니다.
인퓨소리아 신발은 계속해서 빠른 속도로 움직입니다. 속도(실온에서)는 약 2.0-2.5mm/초입니다. 그런 작은 동물에게는 엄청난 속도입니다! 결국, 이것은 신발이 1초에 몸 길이를 10-15배 초과하는 거리를 달린다는 것을 의미합니다. 신발의 궤적은 다소 복잡합니다. 그녀는 프론트 엔드를 똑바로 앞으로 움직입니다.
INFUSORIUM 신발 (PARAMECIUM CAUDATUM)
이 흥미로운 단세포 유기체의 구조와 생활 방식에 대해 알아보기 위해 먼저 한 가지 특징적인 예를 살펴보겠습니다. 얕은 담수 저수지에 널리 퍼져있는 신발 섬모류(파라메시움 속의 종)를 살펴보겠습니다. 이 섬모는 연못을 일반 초원 건초로 채우면 작은 수족관에서 번식하기가 매우 쉽습니다. 이러한 팅크에서는 다양한 유형의 원생동물이 발생하고 신발 섬모가 거의 항상 발생합니다. 일반 교육용 현미경을 사용하면 더 자세히 설명할 내용을 많이 볼 수 있습니다.
가장 단순한 것 중에서 섬모 신발상당히 큰 유기체입니다. 몸 길이는 약 1/6-1/3mm입니다. 몸체의 세로축을 따라 오른쪽으로 회전합니다.
이러한 신발의 적극적인 움직임은 섬모의 몸 전체를 덮는 섬모와 같은 가장 미세한 머리카락과 같은 부속기의 작업에 달려 있습니다. 신발 infusoria의 한 개인에있는 섬모의 수는 10-15,000입니다!
각 섬모는 상온에서 초당 최대 30회까지 노와 같은 움직임을 매우 자주 합니다. 블로우 백 동안 섬모는 곧게 펴진 위치에 유지됩니다. 원래 위치로 돌아갈 때(아래로 이동할 때) 3~5배 느리게 이동하며 반원을 그립니다.
신발이 헤엄칠 때 몸을 덮고 있는 수많은 섬모의 움직임이 집약된다. 개별 섬모의 작용이 조정되어 모든 섬모의 올바른 파동 진동이 발생합니다. 진동파는 몸체의 앞쪽 끝에서 시작하여 뒤로 전파됩니다. 동시에 2-3개의 수축파가 신발의 몸체를 따라 전달됩니다. 따라서 섬모의 전체 모양체 장치는 말하자면 단일 기능적 생리적 전체이며, 개별 구조 단위의 작용(섬모)이 서로 밀접하게 연결(조정)됩니다.
전자현미경 연구에서 알 수 있듯이 각 개별 신발 섬모의 구조는 매우 복잡합니다.
신발의 이동 방향과 속도는 일정하지 않고 불변하는 양이 아닙니다. 모든 살아있는 유기체와 마찬가지로 신발(아메바의 예에서 이미 보았음)은 변화에 반응합니다. 외부 환경이동 방향의 변화.
다양한 자극의 영향으로 원생동물의 이동 방향이 바뀌는 것을 택시라고 합니다. 섬모류에서는 다양한 택시를 관찰하기 쉽습니다. 신발이 뜨는 방울에 신발에 악영향을 미치는 물질(예: 수정 식탁용 소금) 그런 다음 신발은 이 요소에서 멀어지는 것처럼(도망가는 것처럼) 그들에게 불리합니다(그림 86).

우리 앞에 화학적 효과에 대한 부정적인 택시의 예가 있습니다(네거티브 화학주성). 신발과 양성 주화성을 관찰할 수 있습니다. 예를 들어 섬모가 헤엄치는 물 한 방울이 덮개 유리로 덮여 있고 이산화탄소(CO2) 거품이 그 아래에 놓이면 대부분의 섬모가 이 거품으로 이동하여 일정 시간 안에 그 주위에 정착합니다. 반지.
택시 현상은 전류의 영향을받는 신발에서 매우 명확하게 나타납니다. 신발이 뜨는 액체에 약한 전류가 흐르면 다음 그림이 관찰될 수 있습니다. 모든 섬모는 세로축을 전류선과 평행한 방향으로 향하게 한 다음 명령에 따라 음극 쪽으로 이동합니다. 밀도가 높은 클러스터를 형성하는 영역. 전류의 방향에 의해 결정되는 섬모의 움직임을 갈바노탁시라고 합니다. 섬모의 다양한 택시는 다양한 환경 요인의 영향으로 감지될 수 있습니다.
섬모체의 전체 세포질체는 2개의 층으로 명확하게 구분됩니다: 바깥쪽은 더 밝고(엑토플라즘) 내부는 더 어둡고 세분화되어 있습니다(소포체).섬모체의 몸 모양을 일정하게 유지하는 데 중요한 역할을 합니다.
살아있는 신발 몸체의 외층 (외골격)에는 표면에 수직으로 위치한 수많은 짧은 막대가 명확하게 보입니다 (그림 85, 7). 이러한 형성을 트리코포스트라고 합니다. 그들의 기능은 매우 흥미롭고 가장 단순한 것의 보호와 관련이 있습니다. 기계적, 화학적 또는 기타 강한 자극으로 삼차 포낭은 힘으로 버려져 신발을 공격하는 포식자를 때리는 얇은 긴 실로 변합니다. Trichocysts는 강력한 방어입니다. 그들은 섬모 사이에 규칙적으로 배열되어 트리코 포스트의 수가 섬모의 수와 거의 일치합니다. 사용된("샷") 삼모낭 대신에 신발의 세포질에서 새로운 삼모낭이 발생합니다.
한쪽에는 대략 몸의 중앙(그림 85, 5)에 신발이 다소 깊게 함몰되어 있습니다. 이것은 구강 또는 peristome입니다. 섬모는 신체의 표면뿐만 아니라 peristome의 벽을 따라 위치합니다. 그들은 신체 표면의 나머지 부분보다 여기에서 훨씬 더 강력하게 발달합니다. 이 밀접하게 배치된 섬모는 두 그룹으로 배열됩니다. 이러한 고도로 분화된 섬모의 기능은 움직임이 아니라 영양과 관련이 있습니다(그림 87).

신발은 어떻게 그리고 무엇을 먹고 어떻게 소화합니까?
신발은 섬모류 중 하나이며 주요 음식은 박테리아입니다. 박테리아와 함께 영양가와 상관없이 물에 떠 있는 다른 입자도 삼킬 수 있습니다. Perioral cilia는 peristome 깊숙이 위치한 구강 입구 방향으로 부유 입자로 연속적인 물의 흐름을 만듭니다. 작은 음식 입자(대부분 박테리아)는 입을 통해 작은 관 모양의 인두로 침투하여 소포체와의 경계에 있는 바닥에 축적됩니다. 입이 항상 열려있습니다. 섬모 신발이 가장 탐욕스러운 동물 중 하나라고 말하는 것은 실수가 아닐 것입니다. 계속해서 먹이를 먹습니다. 이 과정은 생식 및 성 과정과 관련된 삶의 특정 순간에만 중단됩니다.
인두 바닥에 축적된 음식물 덩어리는 인두 바닥에서 떨어져 나와 소량의 액체와 함께 소포체로 들어가 소화액포를 형성합니다. 후자는 형성 장소에 남아 있지 않지만 소포체의 흐름에 빠지면서 소화 액포의 순환이라고 불리는 신발 몸체에서 다소 복잡하고 규칙적인 경로를 만듭니다 (그림 88). 이 다소 긴(실온에서 약 1시간이 소요되는) 소화 액포의 여행 동안, 그 안에 있는 음식의 소화와 관련하여 그 내부에서 많은 변화가 발생합니다.
여기에서 아메바 및 일부 편모류에서와 같이 전형적인 세포 내 소화가 발생합니다. 소화 액포를 둘러싼 소포체에서 소화 효소가 들어가 음식물 입자에 작용합니다. 소화 생성물은 소화 액포를 통해 소포체로 흡수됩니다.
소화 액포의 순환 과정에서 여러 소화 단계가 대체됩니다. 액포 형성 후 첫 순간에 액포를 채우는 액체는 환경의 액체와 거의 다르지 않습니다. 곧 소화 효소가 소포체에서 액포로 들어가기 시작하고 내부 환경의 반응이 급격히 산성화됩니다. 이것은 환경의 반응(산성, 중성 또는 알칼리성)에 따라 색상이 변하는 일부 지시약을 식품에 추가하면 쉽게 감지할 수 있습니다. 이 산성 환경에서 소화의 첫 번째 단계가 발생합니다. 그러면 그림이 바뀌고 소화액포 내부의 반응이 약간 알칼리성으로 변합니다. 이러한 조건하에서 다음 단계세포 내 소화. 산상은 일반적으로 알칼리상보다 짧습니다. 섬모체의 소화액포 전체 체류 기간의 약 1/6-1/4 동안 지속됩니다. 그러나 산성과 알칼리성의 비율은 식품의 특성에 따라 상당히 넓은 범위 내에서 달라질 수 있습니다.
소포체의 소화 액포의 경로는 그것이 신체의 표면에 접근하고 액체 및 소화되지 않은 음식물 잔류물로 구성된 펠리클을 통해 내용물이 배출된다는 사실로 끝납니다. 배변이 발생합니다. 이 과정은 다른 섬모낭과 마찬가지로 신발에서 배변이 어디에서나 발생할 수 있는 아메바와 달리 복부쪽에 위치한 신체의 특정 부위에만 엄격하게 제한됩니다(복부를 일반적으로 동물의 표면이라고 함 peristome과 신체의 뒤쪽 끝 사이의 대략 중간에 perioral recess가 위치합니다.
따라서 세포 내 소화는 서로를 연속적으로 대체하는 여러 단계로 구성된 복잡한 과정입니다.

계산에 따르면 약 30-45분 안에 섬모체의 부피와 동일한 부피의 체액이 수축성 액포를 통해 신발에서 배출됩니다. 따라서 수축성 액포의 활동으로 인해 섬모체의 몸을 통해 지속적인 물의 흐름이 이루어지며, 이는 입 구멍을 통해 외부에서 (소화 액포와 함께) 삼투압으로 직접 펠리클을 통해 들어갑니다. 수축성 공포는 섬모체를 통과하는 물의 흐름을 조절하고 삼투압을 조절하는 데 중요한 역할을 합니다. 이 과정은 원칙적으로 아메바와 같은 방식으로 진행되며 수축성 액포의 구조만 훨씬 더 복잡합니다.
수년 동안 원생 동물을 연구하는 과학자들 사이에서 수축성 액포의 출현과 관련된 세포질 구조가 있는지 또는 매번 새로 형성되는지에 대한 질문에 대한 논쟁이있었습니다. 살아 있는 섬모에서는 형성되기 이전의 특별한 구조를 관찰할 수 없습니다. 액포(수축기)의 수축이 발생한 후에는 이전 액포가 있던 곳의 세포질에서 구조가 전혀 보이지 않습니다. 그런 다음 투명한 기포 또는 내전 채널이 다시 나타나며 크기가 증가하기 시작합니다. 그러나 새로 출현하는 액포와 기존에 존재하는 액포 사이에는 연관성이 없습니다. 수축성 액포의 연속적인 주기 사이에 연속성이 없고 새로운 수축성 액포가 세포질에서 새로 형성되는 것으로 보입니다. 그러나 특별한 연구 방법에 따르면 실제로는 그렇지 않다는 것이 밝혀졌습니다. 매우 높은 배율(최대 100,000배)을 제공하는 전자 현미경을 사용하여 섬모가 수축성 액포가 형성되는 영역에서 가장 얇은 튜브가 얽힌 형태로 특히 분화된 세포질을 가지고 있음을 설득력 있게 보여주었습니다. 따라서 수축성 액포가 세포질에서 발생하지 않는 것으로 밝혀졌습니다 " 빈 곳", 그러나 이전의 특수 세포 오르가노이드를 기반으로 하며, 그 기능은 수축성 액포의 형성입니다.
모든 원생 동물과 마찬가지로 섬모에는 세포 핵이 있습니다. 그러나 핵 장치의 구조 측면에서 섬모는 다른 모든 원생 동물 그룹과 크게 다릅니다.
섬모의 핵 장치는 이원론이 특징입니다. 이것은 섬모가 큰 핵 또는 대핵과 작은 핵 또는 소핵의 두 가지 다른 유형의 핵을 가지고 있음을 의미합니다. 신발의 섬모에 핵 장치가 어떤 구조를 가지고 있는지 봅시다 (그림 85).
섬모 몸체의 중앙 (연관 수준에서)에는 난형 또는 콩 모양의 큰 거대한 핵이 있습니다. 이것은 거대핵입니다. 그것 가까이에는 몇 배나 더 작은 두 번째 핵이 있으며, 일반적으로 대핵에 아주 가깝습니다. 이것은 소핵입니다. 이 두 핵의 차이는 크기뿐 아니라 구조에 깊숙이 영향을 미치는 더 중요합니다.
대핵은 소핵과 비교하여 염색체의 일부인 특수 핵 물질(염색질, 또는 더 정확하게는 디옥시리보핵산, 약칭 DNA)이 훨씬 더 풍부합니다.
최근 연구에 따르면 대핵은 수십 (일부 섬모 및 수백) 큰 수소핵보다 염색체. 거대핵은 다염색체(다배체) 핵의 매우 독특한 유형입니다. 따라서 미세 핵과 거대 핵의 차이는 염색체 구성에 영향을 미치며, 이는 핵 물질인 염색질의 풍부도를 결정합니다.
섬모의 가장 일반적인 유형 중 하나에서 - 신발(Paramecium caudatum) - 1개의 대핵(약어로 Ma)과 1개의 소핵(약어로 Mi)이 있습니다. 핵 장치의 이러한 구조는 많은 섬모류의 특징입니다. 다른 사람들은 여러 개의 Ma와 Mi를 가질 수 있습니다. 그러나 모든 섬모류의 특징은 핵이 질적으로 다른 두 그룹, 즉 Ma와 Mi로의 분화, 즉 핵 이원론의 현상입니다.
섬모는 어떻게 번식합니까? 다시 infusoria 신발을 예로 들어 보겠습니다. 작은 용기(미세 수족관)에 신발 한 켤레를 심으면 하루에 2개, 그리고 종종 4개의 섬모가 생깁니다. 어떻게 이런 일이 발생합니까? 활동적인 수영과 수유의 특정 기간 후에 섬모는 길이가 다소 늘어납니다. 그런 다음 정확히 몸의 중앙에 점점 깊어지는 가로 수축이 나타납니다(그림 90). 결국 섬모는 말 그대로 반으로 묶이고 한 개체에서 두 개체를 얻습니다. 처음에는 부모 개체보다 약간 작습니다. 전체 핵분열 과정은 실온에서 약 1시간이 걸립니다. 내부 과정에 대한 연구는 가로 수축이 나타나기 전에도 핵 장치의 분열 과정이 시작됨을 보여줍니다. Mi가 먼저 공유되고 Ma 이후에만 공유됩니다. 우리는 여기서 멈추지 않을 것입니다 상세한 고려핵분열 과정에서 우리는 Mi가 유사분열로 나뉘는 반면 Ma의 외관상 분열은 직접적인 핵분열 - 유사분열과 유사하다는 점에만 주목합니다. 신발 섬모류의 무성 생식 과정은 우리가 보는 바와 같이 아메바와 편모의 무성 생식과 유사합니다. 대조적으로, 무성 생식 과정에서 섬모는 항상 가로 질러 분할되는 반면, 편모에서는 분할 평면이 신체의 세로 축과 평행합니다.
분열하는 동안 섬모체의 깊은 내부 구조 조정이 발생합니다. 두 개의 새로운 peristomes, 두 개의 인두 및 두 개의 입 구멍이 형성됩니다. 동시에 섬모의 기초 핵 분열이 시간이 지남에 따라 새로운 섬모가 형성됩니다. 번식하는 동안 섬모 수가 증가하지 않으면 각 분할의 결과로 딸 개인은 어머니 개인의 섬모 수의 약 절반을 받게 되어 섬모의 완전한 "대머리"로 이어질 것입니다. 실제로 이것은 발생하지 않습니다.

때때로 신발을 포함한 대부분의 섬모에는 접합이라고 하는 특별하고 극히 독특한 성 과정이 있습니다. 우리는 이 과정에 수반되는 모든 복잡한 핵 변화를 여기에서 자세히 분석하지 않고 가장 중요한 것만 주목합니다. 접합은 다음과 같이 진행된다(Fig. 91), 두 섬모는 서로 접근하고, 복부 측면에 의해 서로 밀착되며, 이 형태에서 그들은 꽤 헤엄친다. 장기함께 (실온에서 약 12 시간 동안 신발에서). 그러면 접합체가 분리됩니다. 접합 중에 섬모체에서 어떤 일이 발생합니까? 이러한 프로세스의 본질은 다음과 같습니다(그림 91). 큰 핵(대핵)이 붕괴되어 세포질에서 점차 용해됩니다. 소핵이 먼저 분열되고, 핵분열의 결과로 형성된 핵의 일부가 파괴됩니다(그림 91 참조). 각 접합체는 두 개의 핵을 보유합니다. 이 핵들 중 하나는 그것이 형성된 개체의 제자리에 남아 있고(고정 핵), 다른 하나는 적극적으로 접합 파트너(이동 핵)로 이동하여 고정 핵과 합쳐집니다. 따라서이 단계의 각 접합체에는 정지 핵과 이동 핵의 융합 결과로 형성된 하나의 핵이 있습니다. 이 복잡한 핵을 synkaryon이라고합니다. synkaryon의 형성은 수정 과정에 불과합니다. 그리고 다세포 생물에서 수정의 필수적인 순간은 생식 세포 핵의 융합입니다. 섬모에는 생식 세포가 형성되지 않고 서로 합쳐지는 성 핵만 있습니다. 따라서 상호 교배가 발생합니다.
synkaryon이 형성된 직후 접합체가 분리됩니다. 핵 장치의 구조에 따르면, 이 단계에서 그들은 각각 하나의 핵만 가지고 있기 때문에 일반적인 소위 중성(비접합) 섬모와 여전히 매우 크게 다릅니다. 미래에는 synkarion으로 인해 정상적인 핵 장치가 복원됩니다. synkaryon은 분할됩니다(1회 이상). 이 부문의 산물 중 일부는 염색체 수의 증가 및 염색질의 농축과 관련된 복잡한 변형을 통해 대핵으로 변합니다. 다른 것들은 소핵의 구조적 특성을 유지합니다. 이런 식으로 섬모의 특징적이고 전형적인 핵 장치가 복원 된 후 섬모가 분열에 의해 무성 생식을 시작합니다.
따라서 접합 과정에는 수정과 synkaryon으로 인한 새로운 거대핵의 복원이라는 두 가지 필수적인 생물학적 순간이 포함됩니다.
접합의 생물학적 중요성은 무엇이며 섬모의 삶에서 어떤 역할을합니까? 개체 수의 증가가 없기 때문에 번식이라고 할 수 없습니다. 위의 질문은 많은 국가에서 수행된 수많은 실험 연구의 자료로 사용되었습니다. 이들 연구의 주요 결과는 다음과 같다. 첫째, 두 유전 원칙 (부계와 모성)이 하나의 유기체에서 결합하는 다른 성 과정과 마찬가지로 접합은 유전 적 다양성, 유전 적 다양성의 증가로 이어집니다. 유전적 다양성의 증가는 환경 조건에 대한 유기체의 적응 능력을 증가시킵니다. 접합의 두 번째 생물학적으로 중요한 측면은 synkaryon의 핵분열 산물과 동시에 오래된 것의 파괴로 인한 새로운 거대핵의 발달입니다. 실험 데이터에 따르면 섬모의 삶에서 매우 중요한 역할을 하는 것은 거대핵입니다. 그것은 모든 주요 생명 과정을 제어하고 그 중 가장 중요한 것 - 살아있는 세포의 원형질의 주요 부분을 구성하는 단백질의 형성 (합성)을 결정합니다. 분열에 의한 무성 생식이 길어지면 대핵의 "노화"라는 독특한 과정과 동시에 전체 세포가 발생합니다. 신진 대사 과정의 활동이 감소하고 분열 속도가 감소합니다. 접합 후(이 기간 동안 이전 대핵이 파괴됨) 신진 대사 속도와 분열 속도가 회복됩니다. 수정 과정은 대부분의 다른 유기체에서 번식 및 새로운 세대의 출현과 관련이 있는 접합 동안 발생하기 때문에 섬모에서 접합 후에 형성된 개체는 또한 여기에서 발생하는 새로운 유성 세대로 간주될 수 있습니다. 옛것의 "회춘" 때문이었습니다.
섬모 신발의 예에서 우리는 섬모의 광범위한 클래스의 전형적인 대표자를 만났습니다. 그러나 이 클래스는 구조와 생활 방식 모두에서 종의 비범한 다양성이 특징입니다. 가장 특징적이고 흥미로운 형태를 좀 더 자세히 살펴보겠습니다.
섬모에서 신발의 섬모는 신체의 전체 표면을 균일하게 덮습니다. 이것은 구조(Holotricha)의 특징입니다. 많은 섬모는 섬모 덮개의 발달 특성이 다릅니다. 사실 섬모의 섬모는 함께 결합될 때 더 복잡한 복합체를 형성할 수 있습니다. 예를 들어, 서로 가까운 1 또는 2열에 위치한 섬모가 함께 결합(접착)하여 섬모와 같이 두드릴 수 있는 판을 형성하는 것이 종종 관찰됩니다. 이러한 층상 수축 형성을 막(짧은 경우) 또는 막(긴 경우)이라고 합니다. 다른 경우에는 섬모가 함께 결합되어 단단한 묶음에 위치합니다. 이러한 형성 - cirres -는 개별 머리카락이 서로 붙어있는 브러시와 비슷합니다. 다양한 복잡한 모양의 모양이 많은 섬모의 특징입니다. 매우 자주 모양체 덮개는 고르게 발달하지 않고 신체의 일부에서만 발생합니다.
INFUSORIA 트럼펫(STENTOR POLYMORPHIC)
민물에서 크고 아름다운 섬모류에 속하는 종 트럼펫의 종류(스텐토르). 이 이름은 한쪽 끝이 넓게 열린 파이프(그림 92)와 실제로 유사한 이 동물의 몸 모양과 매우 일치합니다. 라이브 트럼펫 연주자를 처음 접하면 신발의 특징이 아닌 한 가지 특징을 알 수 있습니다. 기계적 자극을 포함하여 약간의 자극(예: 유리에 연필로 두드리는 것, 트럼펫 연주로 물방울이 있는 곳)에도 신체가 급격하고 매우 빠르게(1초 미만) 수축하여 거의 규칙적인 자세를 취합니다. 구형. 그런 다음, 다소 천천히(시간은 초 단위로 측정됨) 트럼펫 연주자가 곧게 펴져 특유의 모양을 띠게 됩니다. 트럼펫 연주자가 빠르게 수축하는 능력은 신체와 세포질에 위치한 특수 근육 섬유의 존재 때문입니다. 따라서 근육계는 단세포 유기체에서도 발달할 수 있습니다.

트럼펫의 속에 종들이 있으며, 그 중 일부는 다소 밝은 색상이 특징입니다. 민물에서 매우 흔함 블루 트럼펫 연주자(Stentor coeruleus), 밝은 파란색입니다. 트럼펫 연주자의 이러한 착색은 가장 작은 파란색 안료 알갱이가 엑토플라즘에 위치하기 때문입니다.
트럼펫 연주자(Stentor polymorphus)의 또 다른 종은 종종 녹색입니다. 이 착색의 이유는 상당히 다릅니다. 채색작은 단세포 녹조류가 섬모체의 소포체에서 살고 번식하여 트럼펫 연주자의 몸에 특징적인 색을 부여하기 때문입니다. Stentor polymorphus는 상호 유익한 동거 - 공생의 전형적인 예입니다. 트럼펫 연주자와 조류는 상호 공생 관계에 있습니다. 트럼펫 연주자는 체내에 서식하는 조류를 보호하고 호흡 결과 생성된 이산화탄소를 공급합니다. 조류는 광합성 과정에서 방출되는 산소를 트럼펫 연주자에게 제공합니다. 분명히, 조류의 일부는 섬모에 의해 소화되어 트럼펫 연주자의 먹이가 됩니다.
트럼펫 연주자는 넓은 쪽 끝을 앞으로 내밀고 물 속에서 천천히 헤엄칩니다. 그러나 그들은 또한 작은 빨판이 형성되는 몸체의 뒤쪽 좁은 끝 부분에 의해 기질에 일시적으로 부착 될 수 있습니다.
트럼펫의 몸체에서 뒤쪽에서 앞쪽으로 확장되는 몸통 영역과 그것에 거의 수직으로 위치한 넓은 perioral (peristomal) 필드를 구별 할 수 있습니다. 이 필드는 비대칭 평면 깔때기와 비슷하며 한쪽 가장자리에는 오목한 부분이 있습니다. 인두는 섬모의 소포체로 연결됩니다. 트럼펫 연주자의 몸은 짧은 섬모의 세로 줄로 덮여 있습니다. peristomal field의 가장자리를 따라 강력하게 발달 된 막의 구강 근처 (adoral) 영역이 원 안에 위치합니다 (그림 92). 이 구역은 다수의 개별 섬모판으로 구성되며, 각각의 섬모판은 서로 밀접하게 연결된 두 개의 열에 위치한 많은 섬모로 구성됩니다.
구강 개구부의 영역에서, perioral membranes는 인두 쪽으로 싸여 왼쪽 나선을 형성합니다. 입 주위 막의 진동으로 인한 물의 흐름은 입으로 향하게 됩니다(신체의 앞쪽 끝에 의해 형성된 깔때기의 깊이로). 물과 함께 물에 떠 있는 음식물 입자도 인두로 들어갑니다. 트럼펫 연주자의 음식 대상은 슬리퍼의 대상보다 다양합니다. 박테리아와 함께 작은 원생동물(예: 편모충), 단세포 조류 등을 먹습니다.
트럼펫은 독특한 구조를 가진 잘 발달된 수축성 액포를 가지고 있습니다. 중앙 저장소는 몸의 앞쪽 1/3, 입 구멍 약간 아래에 있습니다. 두 개의 긴 가산 채널이 출발합니다. 그들 중 하나는 저수지에서 신체의 후단으로 가고, 두 번째는 막의 perioral zone과 평행한 peristomal field 영역에 위치합니다.
인퓨소리아 트럼펫은 재생에 관한 실험적 연구에서 가장 좋아하는 대상입니다. 수많은 실험을 통해 트럼펫 연주자의 높은 재생 능력이 입증되었습니다. 가는 메스를 가진 섬모는 여러 부분으로 절단될 수 있으며 각각은 짧은 시간(몇 시간, 때로는 하루 또는 그 이상) 비례하여 만들어 지지만 작은 트럼펫으로 바뀌고 활기찬 먹이의 결과로이 종의 전형적인 크기에 도달합니다. 복구 프로세스를 완료하려면 재생 조각에 구슬 모양의 거대핵 부분이 하나 이상 포함되어야 합니다.
우리가 보았듯이 트럼펫 연주자는 섬모가 다릅니다. 한편으로는 짧고 몸 전체를 덮고 다른 한편으로는 막의 구강 근처 영역이 있습니다. 이러한 구조적 특징에 따라 트럼펫 연주자가 속하는 섬모박리의 이름은 섬모 섬모(헤테로트리카).
INFUSORIA BURSARIA(부르사리아 트렁카텔라)
섬모 섬모의 두 번째 흥미로운 대표자는 종종 민물에서 발견됩니다 부르사리아(Bursaria truncatella, 그림 93). 이것은 섬모 중 거인입니다. 치수는 2mm에 달할 수 있으며 가장 일반적인 크기는 0.5-1.0mm입니다. Bursaria는 육안으로 명확하게 볼 수 있습니다. 이름에 따라 부르사리아는 가방 모양을 하고 있으며 앞쪽 끝이 넓게 열려 있고(bursa는 라틴어로 러시아어로 번역하면 "지갑", "가방"을 의미함) 뒤쪽 끝이 약간 확장됩니다. 섬모의 몸 전체는 짧은 섬모의 세로 줄로 덮여 있습니다. 그들의 구타는 동물의 앞으로 느린 움직임을 유발합니다. 부르사리아는 마치 좌우로 "구르는" 것처럼 헤엄칩니다.

앞쪽 끝에서 몸의 깊이 (길이의 약 2/3)로 peristome이 돌출되어 있습니다. 배 쪽에서는 좁은 슬릿을 통해 외부 환경과 소통하고, 배쪽에서는 연동강이 외부 환경과 소통하지 않습니다. 점액낭 몸체의 상부 1/3의 단면을 보면(그림 93, B), 세포막강이 몸체의 대부분을 차지하는 반면 세포질은 테두리 형태로 그것을 둘러싸고 있음을 알 수 있습니다.
몸의 앞쪽 끝, 왼쪽에는 점액낭에서 매우 강하게 발달 된 perioral (adoral) 막 영역이 시작됩니다 (그림 93, 4). 그것은 연동 구멍의 깊이로 내려가 왼쪽으로 향합니다. adoral zone은 peristome의 가장 깊은 부분까지 확장됩니다. 복강의 복측을 따라 뻗어 있는 모양체 스트립을 제외하고는, perioral membrane을 제외하고 peristomal cavity에 다른 모양체 형성물이 없습니다(그림 93, 10). 내부에 뒷벽 peristomal cavity는 거의 전체 길이를 따라 좁은 슬릿을 가지고 있으며 (그림 93, 7), 그 가장자리는 일반적으로 서로 밀접하게 인접합니다. 이것은 입을 여는 것입니다. 가장자리는 먹을 때만 분리됩니다.
부르사리아는 좁은 음식 전문화가 없지만 주로 포식자입니다. 앞으로 나아가다 보면 다양한 작은 동물들을 만난다. 구강 근처의 막의 작업 덕분에 먹이는 더 이상 수영 할 수없는 광대 한 연동 구멍으로 강제로 끌려갑니다. 음식 물체는 연막강의 등벽에 밀착되어 팽창하는 구강 균열을 통해 소포체를 관통합니다. Bursaria는 매우 탐욕스럽고 오히려 큰 물건을 삼킬 수 있습니다. 예를 들어 가장 좋아하는 음식은 신발 섬모입니다. Bursaria는 6-7 신발을 연속으로 삼킬 수 있습니다. 결과적으로 점액낭의 소포체에 매우 큰 소화액포가 형성됩니다.
부르사리아의 핵 장치는 다소 복잡합니다. 그들은 하나의 긴 소시지 모양의 거대핵과 섬모체의 소포체에 무작위로 흩어져 있는 많은 수(최대 약 30개)의 작은 소핵을 가지고 있습니다.
부르사리아는 수축성 액포가 없는 소수의 민물 섬모종 중 하나입니다. 이 큰 섬모에서 삼투압 조절이 어떻게 수행되는지는 아직 완전히 명확하지 않습니다. 신체의 다른 부분에 있는 bursaria의 세포질 아래에서 체적을 변화시키는 다양한 모양과 크기의 유체 기포-액포를 관찰할 수 있습니다. 분명히 이들은 불규칙한 모양액포이며 기능면에서 다른 섬모의 수축성 액포와 일치합니다.

Bursaria의 무성 생식의 연속 단계를 관찰하는 것은 흥미 롭습니다. 분열의 초기 단계에서 peristome의 전체 공동과 막의 구강 근처 영역이 완전히 감소합니다 (그림 94). 바깥쪽 섬모만 보존됩니다. infusoria는 계란의 형태를 취합니다. 그 후, 몸은 가로 홈으로 두 부분으로 묶입니다. 결과 딸 섬모 각각에서 다소 복잡한 변형을 통해 멤브레인의 전형적인 peristome 및 perioral zone이 발생합니다. 부르사리아를 나누는 전체 과정은 빠르게 진행되며 1시간 이상 걸립니다.
점액낭에서 또 다른 중요한 생명 과정을 관찰하는 것은 매우 쉽습니다. 그 시작은 섬모에 불리한 조건, 낭종 형성(포낭 형성) 과정과 관련이 있습니다. 이 현상은 예를 들어 아메바의 특징입니다. 그러나 섬모류와 같이 복잡하게 조직된 원생동물조차도 비활성 상태로 전환될 수 있음이 밝혀졌습니다. 부르사리아가 서식하는 문화가 제 시간에 먹거나 식지 않으면 몇 시간 안에 대량 엔시티테이션이 시작됩니다. 이 과정은 다음과 같이 진행됩니다. Bursarids는 분열 전과 마찬가지로 막의 peristome과 perioral zone을 잃습니다. 그런 다음 완전히 구형이 된 후 특징적인 모양의 이중 껍질을 구별합니다 (그림 94, D).
낭종의 상태에서 bursaria는 몇 달 동안있을 수 있습니다. 유리한 조건이 발생하면 낭포 껍질이 터지고 점액낭이 빠져 나와 peristome을 개발하고 활동적인 삶으로 이동합니다.
스타일로니키아 미틸러스
매우 복잡하고 다양하게 분화된 섬모 기구는 다음과 관련된 섬모를 가지고 있습니다. 위장의 분리(Hypotricha), 민물과 바닷물 모두에 서식하는 수많은 종. 이 중 가장 일반적이고 자주 접하게 되는 대표자 중 하나는 흥미로운 그룹부를 수 있다 스틸로니키아(Stylonichia mytilus). 이것은 담수 저수지 바닥에 사는 다소 큰 섬모 (길이 최대 0.3mm)입니다. 수생 식물(그림 95). 슬리퍼, 트럼펫, 점액낭과 달리 stylonychia는 연속적인 모양체 덮개가 없으며 전체 모양체 장치는 엄격하게 정의된 제한된 수의 모양체 형성으로 나타납니다.

Stilonychia의 몸체(대부분의 다른 복부 섬모와 마찬가지로)는 등-복부 방향으로 강하게 납작하며 등쪽과 복부 측면, 앞과 뒤 끝이 명확하게 구분됩니다. 몸은 앞쪽으로 약간 넓어지고 뒤쪽으로 좁아집니다. 배 쪽에서 동물을 조사할 때 왼쪽의 앞쪽 1/3에 복잡하게 배열된 깃 모양과 입이 있는 구멍이 있음을 분명히 알 수 있습니다.
등쪽에서는 섬모가 거의 위치하지 않아 이길 수 없습니다. 오히려 얇은 탄성 강모라고 할 수 있습니다. 그들은 움직이지 않으며 운동의 기능과 아무 관련이 없습니다. 이 섬모에는 일반적으로 촉각적이고 민감한 기능이 할당됩니다.
움직임 및 음식 포획과 관련된 모든 모양체 형성은 동물의 복부에 집중되어 있습니다(그림 95). 여러 그룹에 위치한 소수의 두꺼운 손가락 모양의 구조물이 있습니다. 이들은 복부 cirri입니다. 그들 각각은 수십 개의 개별 섬모가 밀접하게 연결되어(함께 붙어 있는) 복잡한 섬모 형성입니다. 따라서 cirres는 개별 머리카락이 밀접하게 결합되어 서로 연결된 브러시와 같습니다.
간경변의 도움으로 동물은 매우 빠르게 움직이며 기질을 따라 "달립니다". 기질에서 "크롤링" 및 "달리기" 외에도 스틸로니키아는 다소 날카롭고 강한 점프를 생성하여 기질에서 즉시 떨어져 나갈 수 있습니다. 이러한 날카로운 움직임은 일반적인 "크롤링"에 참여하지 않는 두 개의 강력한 꼬리 꼬리(그림 95)의 도움으로 수행됩니다.
2열의 변연극모(marginal cirrhae)는 몸의 좌우 가장자리를 따라 위치한다. 동물의 오른쪽 가장자리에서 그들은 몸 전체를 따라 달리고 왼쪽 가장자리에서 그들은 peristome 영역에만 도달합니다. 이 모양체 형성은 동물이 기질에서 분리되어 물 속에서 자유롭게 헤엄칠 때 동물을 추진하는 역할을 합니다.
따라서 우리는 stylonychia의 다양하고 전문화된 모양 기구가 예를 들어 신발이나 트럼펫 연주자와 같이 물에서 단순한 슬라이딩과 대조적으로 매우 다양한 움직임을 수행할 수 있다는 것을 알 수 있습니다.
영양 기능과 관련된 모양체 장치도 복잡합니다. 우리는 이미 인두로 이어지는 입이 아래쪽에 있는 구강 근처 오목부(peristome)가 왼쪽 동물의 앞쪽 절반에 위치한다는 것을 보았습니다. 몸의 맨 앞쪽 끝에서 시작하여 왼쪽 여백을 따라 강하게 발달된 입주위막(adoral) 막 영역이 있습니다. 두들겨 패면서 물의 흐름을 입으로 향하게 합니다. 또한, 연동 오목부 영역에는 내부 끝이 있는 인두로 확장되는 3개의 수축성 막(막)과 여러 개의 특수 섬모가 있습니다(그림 95). 이 전체 복잡한 장치는 음식을 포획하여 입으로 보내는 역할을 합니다.
Stilonychia는 매우 광범위한 음식 개체를 가진 원생 동물 중 하나입니다. 당연히 잡식성이라고 할 수 있습니다. 그녀는 신발처럼 박테리아를 먹을 수 있습니다. 음식 개체 중에는 편모류, 단세포 조류(종종 규조류)가 있습니다. 마지막으로, Stilonychia는 포식자가 되어 다른 작은 종의 섬모를 공격하고 잡아먹습니다.
Stilonychia에는 수축성 공포가 있습니다. 그것은 peristome의 왼쪽 후단에 위치한 중앙 저장소와 뒤쪽으로 향하는 하나의 adductor canal로 구성됩니다.
섬모에서 항상 그렇듯이 핵 장치는 대핵과 소핵으로 구성됩니다.
대핵은 얇은 수축으로 연결된 두 개의 반으로 구성됩니다. 두 개의 소핵이 있으며 Ma의 양쪽 절반 근처에 직접 위치합니다.
Stilonychia, 부분적으로 bursaria, trumpeter - 이들은 모두 다양한 음식 개체를 가진 섬모입니다. 다양한 음식을 흡수하는 능력은 대부분의 섬모류의 특징입니다. 그러나 그들 중에는 식품의 특성과 관련하여 엄격하게 전문화 된 종도 찾을 수 있습니다.
인퓨소리안 프레데터
섬모류 중에는 먹이에 대해 매우 "까다로운" 포식자가 있습니다. 좋은 예가 인퓨소리아입니다. 디디니아(디디늄 나수툼). 디디늄은 평균 길이가 약 0.1-0.15mm인 비교적 작은 섬모입니다. 앞 끝은 코 모양으로 길쭉하며 끝 부분에 입이 있습니다. 섬모 장치는 섬모의 두 화관으로 표시됩니다(그림 96). 디디니우스는 물 속에서 빠르게 헤엄치며 종종 방향을 바꿉니다. 디디니아가 선호하는 음식은 신발 섬모입니다. 이 경우 포식자는 먹이보다 작습니다. 디디니우스는 몸통으로 먹잇감을 꿰뚫고 차츰 입을 벌리면서 신발을 통째로 삼켜버린다! 코에는 소위 막대라고하는 특별한 장치가 있습니다. 그것은 코 주변을 따라 세포질에 위치한 탄성이 강한 강한 막대기로 구성됩니다. 이 장치는 신발과 같은 디디늄과 비교하여 그러한 거대한 먹이를 삼킬 때 파열되지 않는 코 벽의 강도를 증가시키는 것으로 믿어집니다. 디디니우스 - 예 최후의 조치원생동물 사이의 포식. 먹이인 신발을 삼키는 디디늄을 고등 동물의 포식과 비교하면 비슷한 예를 찾기가 어렵습니다.

물론 paramecia를 삼키는 Didinius는 매우 부풀어 오릅니다. 소화 과정은 매우 빠르며 실온에서 약 2시간 정도 걸립니다. 그런 다음 소화되지 않은 잔해를 버리고 디디늄은 다른 희생자를 찾기 시작합니다. 특별 연구에 따르면 didinia의 일일 "다이어트"는 12개의 신발입니다. 이는 정말 엄청난 식욕입니다! 다음 "사냥"사이의 간격에서 didinia가 때때로 분할된다는 것을 명심해야합니다. 음식이 부족하면 디디니아는 매우 쉽게 포낭에서 빠져나오며 쉽게 다시 포낭에서 나옵니다.
초식 섬모
포식보다 훨씬 덜 일반적이며 섬모 "순수한 채식주의"에서 발견됩니다. 식물성 식품. "채식주의자" 섬모의 몇 가지 예 중 하나가 대표자가 될 수 있습니다. 속 패슐라(나술라). 그들의 먹이는 사상성 남조류이다(그림 97).

그들은 측면에있는 입을 통해 소포체로 침투 한 다음 주입에 의해 단단한 나선형으로 꼬여 점차적으로 소화됩니다. 조류 안료는 섬모의 세포질에 부분적으로 들어가서 밝은 짙은 녹색으로 염색합니다.
수보이카 (VORTICELLA NEBULIFERA)
종의 수 측면에서 흥미롭고 다소 큰 섬모 그룹은 기질에 부착된 고착 형태이며, 동그란 눈의 분리(페리트리차). 이 그룹의 가장 일반적인 구성원은 다음과 같습니다. 수보이키(Vorticella 속의 종).
수보이키은방울꽃이나 은방울꽃처럼 우아한 꽃을 닮았고 긴 줄기에 달려 있으며 끝이 밑판에 붙어 있다. suvoyka는 기판에 부착 된 상태에서 대부분의 수명을 보냅니다.

섬모체의 구조를 고려하십시오. 다른 종에서 크기는 상당히 넓은 범위(최대 약 150미크론)에 걸쳐 다양합니다. 구강 디스크 (그림 98)는 섬모가 완전히없는 신체의 확장 된 앞쪽 부분에 있습니다. 모양체 장치는 특수 홈의 구강 (연동) 디스크 (그림 98)의 가장자리를 따라 위치하며 외부에 롤러 (연동 립)가 형성됩니다. 세 개의 섬모 막이 롤러의 가장자리를 따라 실행되며 그 중 두 개는 수직으로, 하나(외부)는 수평입니다. 그들은 나선의 전체 회전을 한 번 이상 형성합니다. 이 막은 지속적으로 깜박이는 동작으로 물의 흐름을 입으로 향하게 합니다. 구강 장치는 짧은 인두로 이어지는 구강 구멍이 있는 깊이에 peristomal field(그림 98)의 가장자리에 깔때기로 시작합니다. Suvoy는 신발처럼 박테리아를 먹습니다. 그들의 입은 끊임없이 열리고 입쪽으로 물이 계속 흐르고 있습니다.
내전관이 없는 하나의 수축성 액포가 입 입구 근처에 있습니다. 대핵은 리본 모양 또는 소시지 모양을 가지고 있으며, 그 옆에 하나의 작은 소핵이 있습니다.
Suvoyka는 코르크 마개로 몇 초 만에 꼬인 줄기를 급격히 줄일 수 있습니다. 동시에 섬모의 몸체도 수축합니다. 연동 디스크와 막이 안쪽으로 당겨지고 전체 앞쪽 끝이 닫힙니다.

질문은 자연스럽게 발생합니다. suvoy가 기질에 부착되어 있기 때문에 저수지에서의 정착은 어떤 방식으로 수행됩니까? 이것은 자유롭게 떠 있는 단계인 방랑자의 형성을 통해 발생합니다. 섬모체 후단부에는 섬모 화관이 나타난다(그림 99). 동시에, 연동 디스크는 안쪽으로 들어가고 섬모는 줄기에서 분리됩니다. 결과 부랑자는 몇 시간 동안 수영할 수 있습니다. 이벤트는 다음에서 재생됩니다. 역순으로: 섬모가 후단부로 기질에 부착되어 줄기가 자라며 섬모의 후부 화관이 축소되고 전단부에서 연동원반이 곧게 펴지며 항문막이 작동하기 시작한다. suvoyka에서 방랑자의 형성은 종종 무성 생식 과정과 관련이 있습니다. 줄기의 주입이 분열하고 딸 개체 중 하나(때로는 둘 다)가 부랑자가 되어 헤엄쳐 갑니다.
많은 유형의 수웍이 불리한 조건에서 엔포스팅할 수 있습니다.
섬모류 그룹에 속하는 고착 섬모류 중에서 위에서 논의한 suvoes와 같은 비교적 소수의 종만이 단독 생활 형태입니다. 여기에 포함된 대부분의 종은 식민지 유기체입니다.
일반적으로 식민지는 불완전하게 완료된 무성 생식 또는 식물 생식에서 발생합니다. 번식의 결과로 형성된 개체는 다소간 서로 연결을 유지하고 함께 더 높은 차원의 유기적 개체를 형성하여 결합합니다. 대량식민지의 이름을 얻는 개별 개인 (우리는 이미 식민지 유기체의 예를 만났습니다. 편모류.

둥근 섬모 섬모의 식민지는 분리 된 개인이 방랑자가되지 않고 줄기의 도움으로 서로 연락을 유지한다는 사실의 결과로 형성됩니다 (그림 100). 이 경우 식민지의 주요 줄기와 첫 번째 가지는 개인에게 귀속될 수 없으며 전체 식민지에 속합니다. 때로는 식민지가 소수의 개인으로 구성되는 반면 다른 종의 섬모에서는 식민지의 개별 개인 수가 수백에 달할 수 있습니다. 그러나 식민지의 성장은 무제한이 아닙니다. 이 종의 특징적인 크기에 도달하면 군체의 증가가 중단되고 분열의 결과로 형성된 개체는 섬모 화관을 발달시키고 떠돌아다니며 헤엄쳐 새로운 군체를 생성합니다.
둥근 섬모 섬모의 식민지는 두 가지 유형이 있습니다. 일부에서 식민지의 줄기는 환원 불가능합니다. 자극을 받으면 식민지의 개별 개인 만 계약하여 깃 모양을 끌어 들이지만 전체 식민지는 변화를 겪지 않습니다 (이 유형의 식민지에는 예를 들어 속이 포함됩니다. Epistylis, Opercularia). 다른 종(예: Carchesium 속)에서는 세포질이 모든 가지를 통과하여 군체의 모든 개체를 서로 연결하기 때문에 전체 군체의 줄기가 수축할 수 있습니다. 그러한 식민지가 자극을 받으면 완전히 축소됩니다. 이 경우 전체 식민지는 유기적 개체로서 전체적으로 반응합니다.
식민지의 모든 둥근 섬모 중에서 특히 흥미로운 것은 아마도 다음과 같습니다. 주탐닉(Zoothamnium arbuscula). 이 섬모의 식민지는 구조의 특별한 규칙 성에 의해 구별됩니다. 또한, 다형성의 흥미로운 생물학적 현상이 콜로니 내에서 여기에 설명되어 있습니다.

Zootamnia 식민지는 우산처럼 보입니다. 하나는 군체의 주요 줄기가 2차 가지이다(그림 101). 성체 군체의 크기는 2~3mm로 육안으로 명확하게 볼 수 있다. 주탐니 거주 작은 연못와 함께 깨끗한 물. 그들의 식민지는 일반적으로 수중 식물에서 발견되며 가장 자주 엘로데아(물 전염병)에서 발견됩니다.
Zootamnia 식민지의 줄기는 수축성 세포질이 주 줄기의 기부 부분을 제외하고 식민지의 모든 가지를 통과하기 때문에 수축합니다. 매우 빠르고 갑자기 발생하는 감소로 전체 식민지가 덩어리로 모입니다.

Zootamnia는 엄격하게 규칙적인 가지 배열이 특징입니다. 하나의 주요 줄기가 기판에 부착됩니다. 식민지의 9 개 주요 가지는 줄기에 수직 인 평면에서 상부에서 출발하며 서로에 대해 엄격하게 규칙적으로 위치합니다 (그림 102, 6). 이차 가지가 이 가지에서 뻗어나와 식민지의 개별 개체가 앉아 있습니다. 각 보조 가지는 최대 50개의 섬모를 가질 수 있습니다. 식민지의 총 개인 수는 2-3,000 명에 이릅니다.
그들의 구조에서 식민지 개체의 대부분은 크기가 40-60 마이크론인 작은 단일 수보이크와 유사합니다. 그러나 성인 식민지에서 마이크로 조이드라고 불리는 작은 개체 외에도 대략 주요 가지의 중간에 완전히 다른 유형과 크기의 개체가 발생합니다 (그림 102, 5). 이들은 직경이 200-250 미크론이고 마이크로 조이드의 부피를 100 배 이상 초과하는 큰 구형 개체입니다. 큰 개체를 마크로조이드라고 합니다.
그들의 구조에서 그들은 식민지의 작은 개인과 크게 다릅니다. 그들의 감정은 표현되지 않습니다. 내면으로 끌어당겨져 작동하지 않습니다. microzoid에서 개발 초기부터 macrozoan은 스스로 음식을 섭취하지 않습니다. 소화액포가 부족합니다. 마크로조이드의 성장은 분명히 식민지의 모든 개체를 서로 연결하는 세포질 다리를 통해 들어가는 물질을 희생시키면서 수행됩니다. 줄기에 붙어있는 거대 동물의 몸 부분에는 특별한 곡물 (과립)이 축적되어 있으며, 이는 우리가 볼 수 있듯이 향후 운명에 중요한 역할을합니다. 이 큰 구형 거대동물은 무엇이며 주탐니아 식민지의 삶에서 생물학적 역할은 무엇입니까? 관찰에 따르면 마크로조이드는 새로운 식민지가 발달하는 미래의 방랑자임을 보여줍니다. 도달 크기 제한, 거대동물은 섬모 화관을 발달시키고 군체에서 분리되어 헤엄쳐 나간다. 동시에 모양이 약간 바뀌며 구형에서 원추형이됩니다. 얼마 후 트램프는 입도가있는면이있는 기판에 항상 부착됩니다. 줄기의 형성과 성장이 즉시 시작되고 과립은 방랑자의 후단에 국한된 줄기의 건설에 소비됩니다. 줄기가 자라면서 입자가 사라집니다. 줄기가 Zootamnia의 특징적인 최종 길이에 도달한 후, 일련의 빠르게 연속적인 분열이 시작되어 식민지를 형성합니다. 이러한 구분은 엄격하게 정의된 순서로 이루어집니다(그림 102).
우리는 이 프로세스의 세부 사항에 대해 이야기하지 않을 것입니다. 다음과 같은 흥미로운 현상에만 주목합시다. Zootamnia 트램프의 첫 번째 분열 동안, 형성 개체에서 식민지가 발달하는 동안 깃 모양과 입은 기능하지 않습니다. 어린 식민지가 이미 12-16 개인으로 구성된 나중에 먹이를 먹기 시작합니다. 따라서 식민지 개발의 모든 첫 번째 단계는 모체에서 성장하고 발달하는 동안 거대 동물의 몸에 형성된 매장량을 희생하여 독점적으로 수행됩니다. Zootamnia vagrant의 발달과 다세포 동물의 난자의 발달 사이에는 부인할 수 없는 유사점이 있습니다. 이 유사성은 두 경우 모두의 개발이 외부 환경의 음식에 대한 인식 없이 이전에 축적된 매장량을 희생하여 수행된다는 사실로 표현됩니다.
고착성 섬모 섬모를 연구 할 때 질문이 생깁니다. 섬모의 성적인 과정의 형태가 어떻게 수행됩니까? 앉아있는 생활 방식과 관련하여 몇 가지 중요한 변화가 있음이 밝혀졌습니다. 성적인 과정이 시작될 때 식민지에 특별하고 매우 작은 방랑자가 형성됩니다. 섬모 화관의 도움으로 활발히 움직이는 그들은 식민지를 따라 얼마 동안 기어 다니다가 식민지의 큰 정상 앉아있는 개인과 결합합니다. 따라서 접합체를 두 그룹의 개인으로 구분하는 것은 여기에서 발생합니다: 작고 이동성 있는(미세 접합체) 및 더 큰 부동성(거대 접합체). 접합체를 두 가지 범주로 구분하는 것과 같이 그 중 하나(미세 접합체)는 이동성이 있으며, 이는 좌식 생활 방식에 필요한 적응이었습니다. 이것이 없으면 성적 과정(접합)의 정상적인 과정이 분명히 보장될 수 없습니다.
빨기 솜털 (Suctoria)
먹는 방식 면에서 매우 독특한 그룹은 다음과 같이 표시됩니다. 빨기 섬모(수토리아). suvoyka 및 기타 섬모 섬모와 같은 이러한 유기체는 고착성입니다. 이 목에 속하는 종의 수는 수십 개로 측정됩니다. 빨기 섬모의 몸 모양은 매우 다양합니다. 그들의 특징적인 종 중 일부는 그림 103에 나와 있습니다. 일부는 다소 긴 줄기의 기질에 앉아 있고, 다른 일부는 줄기가 없고, 일부는 몸의 가지가 아주 강하게 가지 않습니다. 그러나 다양한 형태에도 불구하고 모든 빨기 섬모는 다음 두 가지 특징이 있습니다.
1) 모양체 장치의 완전한 부재(성인 형태),
2) 특별한 부속기의 존재 - 먹이를 빨아들이는 역할을 하는 촉수.

빨기 섬모의 다른 유형에서 촉수의 수는 동일하지 않습니다. 종종 그들은 그룹으로 모입니다. 현미경의 고배율로 촉수 끝에 작은 곤봉 모양의 두꺼워진 부분이 있음을 알 수 있습니다.
촉수는 어떻게 작동합니까? 잠시 동안 섬모를 빠는 것을 관찰하여 이 질문에 답하는 것은 어렵지 않습니다. 일부 작은 원생 동물 (편모, infusoria)이 suctoria의 촉수에 닿으면 즉시 달라 붙습니다. 피해자가 탈출하려는 모든 시도는 대개 헛수고입니다. 촉수에 달라붙은 먹이를 계속 관찰하면 점차 크기가 줄어들기 시작하는 것을 볼 수 있습니다. 그 내용물은 희생자에게서 단 하나의 펠리클만 남을 때까지 촉수를 통해 빨아들이는 섬모의 소포체로 점차 "펌핑"되어 폐기됩니다. 따라서 섬모를 빠는 촉수는 완전히 독특하며 동물 세계의 다른 곳에서는 덫을 놓는 동시에 음식을 빨아먹는 기관이 없습니다(그림 103).
빠는 섬모는 먹이를 쫓지 않지만 부주의한 먹이가 그 자체에 닿기만 하면 즉시 잡는 움직이지 않는 포식자입니다.

왜 우리는 이 독특한 유기체를 섬모류의 부류로 지칭합니까? 언뜻보기에 그들은 그들과 아무 관련이 없습니다. 다음 사실은 섬모에 suctoria가 속하는 것에 대해 말합니다. 첫째, 그들은 거대 핵과 소핵으로 구성된 섬모의 전형적인 핵 장치를 가지고 있습니다. 둘째, 번식하는 동안 "성인"개인에게는없는 섬모가 발생합니다. 무성 생식과 동시에 빨기 섬모의 재정착은 섬모의 여러 환형 화관이 장착 된 방랑자의 형성에 의해 수행됩니다. suctoria에서 방랑자의 형성은 다른 방식으로 발생할 수 있습니다. 때때로 그들은 바깥쪽으로 분리되는 각 신장이 대핵 분절과 하나의 소핵을 받는 아주 균일하지 않은 분열(출아)의 결과로 형성됩니다(그림 104, L). 한 명의 모성 개체에서 여러 개의 딸 싹이 한 번에 형성될 수 있습니다(그림 104, 5). 다른 종(그림 104, D, E)에서는 "내부 출아"의 매우 독특한 방법이 관찰됩니다. 동시에, 부정기선 신장이 형성되는 모낭의 몸체 내부에 공동이 형성됩니다. 그것은 알려진 어려움으로 "압착"하는 특수 구멍을 통해 나옵니다.
어머니의 몸 안에서 태아가 발달한 다음 출산하는 행위는 고등 다세포 생물에서 일어나는 일과 가장 단순한 일에 대한 흥미로운 비유입니다.
이전 페이지에서 섬모류 클래스의 몇 가지 전형적인 자유 생활 대표자가 다양한 환경 조건에 다르게 적응하는 것으로 간주되었습니다. 섬모를 생활 조건에 적응시키는 문제에 접근하는 것과 다른 한편으로 특징이 무엇인지 보는 것은 흥미 롭습니다. 일반적인 특징섬모류는 특정하고 명확하게 정의된 환경 조건에서 생활합니다.
예를 들어 플랑크톤 구성의 생명체와 모래 두께의 바닥 생명체라는 두 개의 매우 다른 서식지를 예로 들어 보겠습니다.
플랑크톤 인퓨소리아
상당히 많은 수의 섬모종이 해양 및 민물 플랑크톤에서 발견됩니다.
수중 생활에 대한 적응의 특징은 특히 방산충에서 두드러집니다. 플랑크톤 생활 방식에 대한 주요 적응 라인은 수주에서 유기체의 급증에 기여하는 구조적 특징의 발달로 축소됩니다.

게다가 거의 독점적으로 바다 섬모류의 전형적인 플랑크톤은 다음과 같습니다. 틴틴(Tintinnidae, 그림 105, 5). 지금까지 알려진 틴티니드 종의 총 수는 약 300입니다. 이들은 섬모의 원형질체가 유기물로 구성된 투명하고 가벼우 며 동시에 강한 집에 배치된다는 사실을 특징으로하는 작은 형태입니다. 집에서 디스크가 튀어 나와 끊임없이 깜박이는 섬모 화관을 가지고 있습니다. 수주에 떠있는 infusoria 상태에서 주로 모양체 장치의 지속적인 활성 작업에 의해 지원됩니다. 집은 분명히 섬모의 하체를 보호하는 기능을 수행합니다. 담수에는 2종의 틴티니드만이 산다(바이칼 호수만의 특징인 7종은 제외).
민물 섬모는 플랑크톤의 삶에 다른 적응 기능을 가지고 있습니다. 그들 중 다수에서 세포질은 매우 강하게 액포화되어 있어(Loxodes, Condylostoma, Trachelius) 거품과 비슷합니다. 이것은 비중의 상당한 감소로 이어집니다. 나열된 모든 섬모에는 또한 섬모 덮개가있어 비중 측면에서 섬모 몸체가 비중물, 쉽게 "급상승" 상태로 유지됩니다. 일부 종에서는 몸체의 모양이 비표면적의 증가에 기여하고 물에서 솟아오르는 것을 촉진합니다. 예를 들어, 바이칼 호수의 일부 플랑크톤 섬모는 모양이 우산이나 낙하산과 비슷합니다(Liliomorpha, 그림 105, 2). 바이칼 호수(Mucophrya pelagica, 그림 105, 4)에는 플랑크톤을 빨아들이는 섬모류가 하나 있는데, 이는 고착성 친척과 크게 다릅니다. 이 종에는 줄기가 없습니다. 그것의 원형질 몸은 체중 감소로 이어지는 적응된 넓은 칙칙한 외피로 둘러싸여 있습니다. 길고 얇은 촉수가 튀어 나와 직접적인 기능과 함께 아마도 또 다른 기능을 수행 할 것입니다. 비표면적이 증가하여 물에서 치솟는 데 기여합니다.
마지막으로, 말하자면 섬모가 플랑크톤의 삶에 적응하는 간접적인 형태를 하나 더 언급할 필요가 있습니다. 이것은 플랑크톤 생활 방식을 이끄는 다른 유기체에 작은 섬모가 부착되는 것입니다. 네, 중에서 할례받은 섬모(Peritricha) 플랑크톤 요각류에 부착하는 상당히 많은 종이 있습니다. 이것은 이러한 유형의 섬모에 대한 정상적이고 정상적인 삶의 방식입니다.
주위 섬모와 함께 빠는(Suctoria) 플랑크톤 유기체에 정착하는 종이 있습니다.
모래에 사는 시민
모래 해변과 여울은 매우 독특한 서식지를 나타냅니다. 바다 연안을 따라 그들은 광대 한 공간을 차지하고 독특한 동물 군이 특징입니다.
위해 수행 지난 몇 년여러 국가의 수많은 연구에 따르면 많은 해양 모래의 지층에는 다양한 미시적 또는 미시적 동물군이 매우 풍부합니다. 모래 입자 사이에는 물로 채워진 수많은 작고 작은 공간이 있습니다. 이 공간은 동물 세계에서 가장 다양한 그룹에 속하는 유기체로 풍부하게 채워져 있습니다. 수십 종의 갑각류가 여기에 서식하며, 환형동물, 회충, 특히 수많은 편형동물, 일부 연체동물, 장충. 다수의 원생동물, 주로 섬모류도 있습니다. 현대 데이터에 따르면, 두꺼운 바다 모래에 서식하는 섬모류 동물군의 구성은 약 250-300종을 포함합니다. 섬모류뿐만 아니라 모래 두께에 서식하는 다른 유기체 그룹을 염두에 두면 해당 종의 총 수가 매우 많을 것입니다. 모래 알갱이 사이의 가장 작은 틈에 사는 모래 두께의 동물 전체를 psammophilic 동물이라고합니다.
psammophilic 동물군의 풍부함과 종 구성은 많은 요인에 의해 결정됩니다. 그 중에서도 특히 중요성모래 입자의 크기를 가지고 있습니다. 거친 모래는 열악한 동물군을 가지고 있습니다. 입자 직경이 0.1mm 미만인 매우 미세한 미사질 모래의 동물군도 열악하며, 분명히 입자 사이의 간격이 동물이 살기에는 너무 작습니다. 인생에서 가장 풍부한 모래는 중간 및 세립입니다.
psammophilic 동물군의 발달에 중요한 역할을 하는 두 번째 요소는 유기물 잔해와 부패하는 유기물(소위 saprobity의 정도)에 있는 모래의 풍부함입니다. 유기물이 없는 모래는 삶이 가난하다. 반면에, 모래도 거의 생명이 없고 유기물의 붕괴로 인해 산소가 고갈되기 때문에 유기물이 매우 풍부합니다. 종종 혐기성 황화수소 발효가 여기에 추가됩니다.
유리 황화수소의 존재는 동물군의 발달에 영향을 미치는 극히 부정적인 요인입니다.
단세포 조류(규조류, peridiniums)의 상당히 풍부한 식물상이 때때로 모래 표층에서 발생합니다. 이것은 많은 작은 동물(섬모류 포함)이 조류를 먹기 때문에 psammophilic 동물군의 발달에 유리한 요소입니다.
마지막으로 친친매성 동물군에 매우 부정적인 영향을 미치는 요인은 서핑이다. 이것은 모래의 상층을 씻는 파도가 여기의 모든 생물을 죽이기 때문에 이해할 수 있습니다. psammophilic 동물은 보호되고 잘 가열 된 만에서 가장 풍부합니다. 밀물과 썰물은 psammophilic 동물군의 발달을 막지 못합니다. 썰물 때 물이 일시적으로 빠져나가 모래를 드러낸 다음 모래의 굵기, 모래알 사이의 간격으로 물이 남아 있어 동물의 존재를 막지 못한다.
psammophilous 동물 군의 일부이며 다양한 체계적인 그룹 (목, 가족)에 속하는 섬모류에서는 모래 입자 사이의 독특한 존재 조건에 적응하는 진화 과정에서 많은 공통 기능이 개발됩니다.

그림 106은 다른 목과 과에 속하는 섬모류의 일부 종을 보여줍니다. 그들 사이에는 많은 유사점이 있습니다. 그들 대부분의 몸은 길이가 다소 강하게 길쭉하고 벌레와 비슷합니다. 이렇게 하면 모래 알갱이 사이의 가장 작은 구멍으로 "밀어내기"가 쉽습니다. 매우 많은 종에서(그림 106) 몸체의 신장은 편평함과 결합됩니다. 섬모 장치는 항상 잘 발달되어있어 일정한 힘으로 좁은 틈에서 움직일 수 있습니다. 종종 섬모는 벌레처럼 납작한 몸의 한쪽에 발달하고 반대쪽은 맨손입니다. 이 특징은 아마도 대부분의 psammophilic 종에서 발음되는, 모양체 장치를 통해 기질에 매우 밀접하고 매우 단단히 부착(부착)하는 능력(thigmotaxis라고 하는 현상)과 관련이 있을 것입니다. 이 속성은 동물이 살고 있는 좁은 틈에 물의 흐름이 발생하는 경우 제자리에 머물 수 있도록 합니다. 이 경우, 동물이 기질에 부착된 반대면이 매끄럽게 하는 것이 더 유리할 수 있다.
psammophilic 섬모는 무엇을 먹습니까? 많은 종에서 "식이 요법"의 중요한 부분은 조류, 특히 규조류입니다. 박테리아는 음식으로 덜 먹습니다. 그것은 또한 심하게 오염되지 않은 모래에 박테리아가 거의 없다는 사실에 크게 의존합니다. 마지막으로, 특히 가장 큰 psammophilous ciliates 중에는 더 작은 종의 다른 ciliates를 먹는 포식 형태가 상당히 많습니다. Psammophilic ciliates는 분명히 모든 곳에 분포합니다.
인퓨소리아 아포스토메이트

섬모 스피로프리아포낭 상태의 (Spirophrya subparasitica)는 작은 플랑크톤 해양 갑각류(특히 종종 Idia 속의 갑각류)의 작은 줄기에 앉아 있는 경우가 많습니다. 갑각류가 바닷물에서 활발하게 헤엄치는 동안, 그 위에 앉아 있는 스피로프리아는 아무런 변화를 겪지 않습니다. 섬모의 추가 발달을 위해서는 갑각류가 해양 하이드로 로이드 폴립에 의해 먹는 것이 필요합니다. 이는 꽤 자주 발생합니다 (그림 107). spirophria 낭종은 갑각류와 함께 소화관에 들어가 자마자 작은 섬모가 즉시 나와 삼킨 갑각류의 소화로 인한 음식 죽을 격렬하게 먹기 시작합니다. 1 시간 이내에 infusoria의 크기가 3-4 배 증가합니다. 그러나 이 단계에서는 번식이 일어나지 않습니다. 우리 앞에는 섬모의 전형적인 성장 단계가 있으며, 이를 트로폰이라고 합니다. 얼마 후 소화되지 않은 음식 잔류 물과 함께 trophont는 폴립에 의해 바닷물로 버려집니다. 여기에서 적극적으로 수영하면서 폴립의 몸을 따라 밑창까지 내려와 낭종으로 둘러싸여 자체 부착됩니다. 포낭이 있는 큰 섬모가 폴립 위에 앉아 있는 이 단계를 토몬트라고 합니다. 이것은 번식 단계입니다. Tomont는 먹이를 먹지 않지만 연속적으로 여러 번 빠르게 나눕니다 (그림 107, 7). 결과는 매우 작은 섬모의 전체 그룹입니다. 그들의 수는 토몬트의 크기에 따라 달라지며, 토몬트는 토몬트의 기원을 제공한 트로폰의 크기에 따라 결정됩니다. 토몬트의 분할 결과 형성된 작은 섬모(토마이트 또는 방랑자라고 함)는 정착 단계를 나타냅니다.
그들은 낭종을 떠나 빠르게 수영합니다 (동시에 먹지 않고 세포질에있는 매장량을 사용). 그들이 요각류를 만날 수 있을 만큼 "운이 좋은" 경우, 그들은 즉시 그것에 붙어서 포집합니다. 이것이 우리가 주기에 대한 고려를 시작한 단계입니다.
검토된 라이프 사이클스피로프리아는 생물학적 중요성이 다른 단계의 날카로운 구분에 주의를 기울입니다. Trophon은 성장 단계입니다. 그것은 활발하고 빠른 영양으로 인해 많은 양의 세포질과 모든 종류의 예비 물질을 성장시키고 축적합니다. 트로폰은 번식이 불가능합니다. 반대 현상은 tomont에서 관찰됩니다 - 먹이를 먹을 수없고 활발한 빠른 번식. 각 분할 후에는 성장이 없으므로 tomont의 번식은 많은 방랑자로 급속한 부패로 축소됩니다. 마지막으로, 방랑자는 그들의 특별하고 유일한 특징적인 기능을 수행합니다. 그들은 개인, 즉 종의 정착자이자 분배자입니다. 그들은 먹거나 번식할 수 없습니다.
Ichthyophthirius의 수명주기

성장 기간이 끝날 때까지 ichthyophthirius는 직경 0.5-1mm의 방랑자에 비해 매우 큰 크기에 이릅니다. 한계 값에 도달하면 섬모는 물고기의 조직에서 적극적으로 물 속으로 이동하고 몸 전체를 덮는 섬모 장치의 도움으로 잠시 천천히 수영합니다. 곧 큰 ichthyophthirius가 수중 물체에 정착하여 낭종을 분비합니다. entsting 직후 섬모의 연속적인 분할이 시작됩니다. 처음에는 절반으로, 각 딸 개인은 다시 두 개로 나뉘고 10-11 번까지 계속됩니다. 결과적으로 낭포 내부에 섬모로 덮인 최대 2000개의 작고 거의 둥근 개체가 형성됩니다. 낭종 내부에서 방랑자가 활발히 움직이고 있습니다. 그들은 껍질을 뚫고 나옵니다. 활발하게 헤엄치는 방랑자는 새로운 물고기를 감염시킵니다.
낭포에서 ichthyophthirius의 분열 속도와 물고기 조직에서의 성장 속도는 온도에 크게 좌우됩니다. 다양한 저자의 연구에 따르면 다음 수치가 제공됩니다. 26-27°C에서 낭종의 발달 과정은 10-12시간, 15-16°C에서 28-30시간, 4시 -5°C에서 6-7일 동안 지속됩니다.
ichthyophthirius와의 싸움은 상당한 어려움을 나타냅니다. 여기에서 가장 중요한 것은 자유롭게 수영하는 떠돌이가 물고기의 조직에 침투하는 것을 방지하기 위한 예방 조치입니다. 이렇게하려면 ichthyophthirius와의 싸움에서 특히 효과적인 흐름 조건을 만들기 위해 아픈 물고기를 새로운 저수지 또는 수족관에 자주 이식하는 것이 유용합니다.
섬모 트리코다인

숙주 표면의 생명체에 대한 트리코딘의 적응 전체 시스템은 이동성을 유지하면서 숙주의 신체(거의 항상 죽음에 해당함)에서 이탈하지 않는 것을 목표로 합니다. 이러한 장치는 매우 완벽합니다. 대부분의 트리코디나스의 몸체는 다소 납작한 원반 모양이며 때로는 뚜껑이 있습니다. 숙주의 몸과 마주하는 면은 약간 오목하여 부착흡반을 이룬다. 빨판의 바깥 쪽 가장자리를 따라 잘 발달 된 섬모의 화관이 있으며 섬모의 움직임 (크롤링)은 주로 물고기의 몸 표면에서 발생합니다. 이 화관은 위에서 논의한 고착성 둥근 섬모 섬모 방랑자에서 발견되는 화관에 해당합니다. 따라서 trichodina는 트램프와 비교할 수 있습니다. 복부 표면(흡반 위)에서 트리코딘은 숙주에 섬모를 유지하는 데 도움이 되는 매우 복잡한 지지 및 부착 장치를 가지고 있습니다. 구조의 세부 사항으로 들어가지 않고 기본은 외부 및 내부 치아를 포함하는 별도의 세그먼트로 구성된 복잡한 구성의 링입니다(그림 109, B). 이 고리는 흡반 역할을 하는 복부 표면의 탄력성과 동시에 강력한 기초를 형성합니다. 다른 유형의 트리코딘은 고리를 형성하는 세그먼트의 수와 외부 및 내부 후크의 구성이 서로 다릅니다.
편모체의 원반 맞은편 측면에는 깃 모양의 구(口)구가 있다. 그 구조는 다소 전형적인 할례받은 섬모. 시계 방향으로 꼬인 항문 막은 아래쪽에 입이 있는 오목한 부분으로 이어집니다. 트리코딘의 핵 장치는 일반적으로 섬모를 위해 배열됩니다: 하나의 리본 모양의 거대핵과 그 옆에 위치한 하나의 소핵. 수축성 공포가 하나 있습니다.
트리코딘은 모든 유형의 저장소에 널리 분포되어 있습니다. 특히 종종 그들은 다른 종의 물고기 튀김에서 발견됩니다. 대량 번식으로 트리코딘이 적용됩니다. 큰 피해물고기, 특히 대중이 아가미를 덮는 경우. 이것은 물고기의 정상적인 호흡을 방해합니다.
trichodin의 물고기를 청소하려면 2 % 염화나트륨 용액 또는 0.01 % 과망간산 칼륨 용액으로 치료 목욕을 만드는 것이 좋습니다 (튀김 - 10-20 분 동안).
UNGATE의 장내 염증
흉터에서 메쉬를 통해 음식이 구강으로 트림되어 추가로 씹습니다(껌). 식도의 주름에 의해 형성된 특별한 관을 통해 다시 삼켜진 씹은 음식 덩어리는 더 이상 흉터로 가지 않고 책으로, 거기에서 복부로 이동하여 반추동물의 소화액에 노출됩니다. rennet에서는 산성 반응과 소화 효소의 존재 조건에서 섬모가 죽습니다. 껌으로 거기에 도착하면 소화됩니다.
반추위(그물과 마찬가지로)에 있는 원생동물의 수는 엄청난 값에 도달할 수 있습니다. 흉터의 내용물을 한 방울 떨어뜨려 현미경으로 관찰하면(가열 시 섬모가 실온에서 멈추므로) 시야에 섬모가 말 그대로 떼를 지어 옵니다. 문화에서도 그러한 덩어리의 섬모를 얻는 것은 어렵습니다. 흉터 내용의 1cm3에있는 섬모 수는 백만, 종종 더 많습니다. 흉터의 전체 부피 측면에서 이것은 진정한 천문학적 수치를 제공합니다! 섬모가 있는 반추위 내용물의 풍부함은 반추 동물의 음식 특성에 크게 좌우됩니다. 음식에 섬유질이 풍부하고 탄수화물과 단백질(잔디, 짚)이 부족하면 반추위에는 섬모가 상대적으로 적습니다. 탄수화물과 단백질(겨)이 식단에 추가되면 섬모의 수가 급격히 증가하여 엄청난 수에 도달합니다. 섬모의 지속적인 유출이 있음을 명심해야합니다. 껌과 함께 abomasum에 들어가면 죽습니다. 높은 레벨섬모의 수는 활발한 번식으로 유지됩니다.
홀수 발가락 유제류(말, 당나귀, 얼룩말)도 소화관에 많은 섬모가 있지만 숙주에서의 국소화는 다릅니다. 홀수 발가락 유제류는 위장이 복잡하지 않기 때문에 소화관의 앞쪽 부분에서 원생 동물이 발생할 가능성이 없습니다. 그러나 말의 경우 큰 맹장과 맹장이 매우 잘 발달되어 있어 일반적으로 음식물 덩어리로 막혀 소화에 필수적인 역할을 합니다. 반추동물의 반추위와 그물망에서와 같이 장의 이 부분에서 원생동물의 매우 풍부한 동물군, 주로 섬모류가 발생하며, 대부분은 또한 내분비형(endodiniomorphs) 목에 속합니다. 그러나 종 구성면에서 반추동물의 반추위 동물군과 말의 대장군 동물군은 일치하지 않는다.
반추동물의 장의 도상인
가장 큰 관심은 섬모입니다 ofrioscolecid 가족(Ophryoscolecidae), 관련 내분비형을 주문하다. 이 분리의 특징은 연속적인 모양체 덮개가 없다는 것입니다. 복잡한 모양체 형성 - cirres -는 입이 열리는 부분의 섬모체 몸체의 앞쪽 끝에 위치합니다. 모양체 장치의 이러한 기본 요소는 몸의 앞쪽이나 뒤쪽 끝에 위치한 추가 극모 그룹으로 보완될 수 있습니다. ofrioscolecid 가족의 섬모의 총 종의 수는 약 120입니다.

그림 110은 반추동물의 반추위에서 나온 가장 전형적인 리오스코레시드의 일부를 보여줍니다. Entodinium 속의 섬모(Etodinium, 그림 110, L)는 가장 간단하게 배열되어 있습니다. 그들의 몸의 앞쪽 끝에는 cirrus의 perioral zone이 있습니다. 입구가 있는 몸체의 앞부분은 안쪽으로 끌어당길 수 있습니다. 세포질과 소포체는 뚜렷하게 구분됩니다. 항문관은 소화되지 않은 음식물 찌꺼기를 제거하는 역할을 하는 뒤쪽 끝에 명확하게 보입니다. 다소 복잡한 구조 무배증(아노플로디늄, 그림 110, B). 그들은 섬모 장치의 두 영역이 있습니다 - perioral cirrhi와 dorsal cirrhi. 둘 다 프론트 엔드에 있습니다. 그림에 표시된 종의 몸체 뒤쪽 끝에는 길고 날카로운 파생물이 있습니다. 이것은 많은 종의 리오스콜레시드에서 매우 일반적입니다. 이러한 파생물은 흉터를 채우는 식물 입자 사이의 섬모를 "밀어내는" 데 기여한다고 제안되었습니다.
종류 속 유디플로디니아(유디플로디늄, 그림 110, B)는 다음과 유사합니다. 무배증, 그러나 그들과 달리 인두를 따라 오른쪽 가장자리에 골격 기반 플레이트가 있습니다. 이 골격 판은 다음과 유사한 물질로 구성됩니다. 화학적 성질섬유질, 즉 식물 세포의 껍질을 구성하는 물질.
~에 속 폴리플라스트론(Polyplastron, 그림 110, D, E) 골격의 추가 합병증이 있습니다. 이 섬모의 구조는 eudiplodynia에 가깝습니다. 차이점은 주로 하나의 골격 판 대신에 5개의 섬모가 있다는 사실로 요약됩니다. 그 중 가장 큰 두 개는 오른쪽에 있고 작은 세 개는 섬모 왼쪽에 있습니다. 폴리플라스트론의 두 번째 특징은 수축성 액포의 수가 증가한다는 것입니다. 엔토디니아는 1개의 수축성 액포를 가지고 있으며, anoplodynia와 eudiplodynia는 2개의 수축성 액포를 가지고 있으며, polyplastron은 그 중 약 12개를 가지고 있습니다.
~에 상피(Epidinium, 그림 110) 몸의 오른쪽에 잘 발달 된 탄수화물 골격이있는 간극의 등쪽 영역은 앞쪽 끝에서 뒤쪽으로 이동합니다. 가시는 종종 이 속의 섬모의 뒤쪽 끝에 발달합니다.
가장 복잡한 구조가 드러납니다. 속 Ofrioscolex(Ophryoscolex), 그 후에 섬모의 전체 가족이 명명됩니다 (그림 110, E). 그들은 몸과 골격 판 둘레의 약 2/3를 덮는 잘 발달 된 등쪽 권운이 있습니다. 많은 가시가 후단에 형성되며 그 중 하나는 일반적으로 특히 길다.
일부 전형적인 대표자와의 친분 오브리오스코레시드이 가족 내에서 조직의 중요한 합병증이 있음을 보여줍니다(내통에서 ofrioscolex까지).
섬모 외에 ofrioscolecid 가족, 반추 동물의 반추위에서 우리에게 이미 알려진 대표자는 소량으로 발견됩니다. 등섬모섬모 분리. 그들은 소수의 종으로 대표됩니다. 그들의 몸은 섬모의 세로 줄로 고르게 덮여 있으며 골격 요소는 없습니다. 반추위 섬모 인구의 총 질량에서 중요한 역할을하지 않으므로 여기에서 다루지 않을 것입니다.
섬모류 ofrioscolecides는 무엇을 어떻게 먹습니까? 이 문제는 많은 과학자들, 특히 V. A. Dogel 교수에 의해 자세히 연구되었습니다.

Ofrioscolecid 음식은 매우 다양하며 다른 종에서 특정 전문화가 관찰됩니다. Entodynia 속의 가장 작은 종은 박테리아, 전분 알갱이, 곰팡이 및 기타 작은 입자를 먹습니다. 매우 많은 중형 및 대형 리오스콜레시드가 반추위 내용물의 대부분을 구성하는 식물 조직의 입자를 흡수합니다. 일부 종의 소포체는 말 그대로 식물 입자로 막혀 있습니다. 섬모가 어떻게 식물 조직 조각을 습격하고 말 그대로 조각으로 찢은 다음 삼키며 종종 몸에서 나선형으로 비틀리는지를 볼 수 있습니다(그림 111, 4). 때로는 삼킨 큰 입자로 인해 섬모 자체의 몸체가 변형 될 때 그러한 그림 (그림 111, 2)을 관찰해야합니다.
ofrioscolecid에서는 때때로 포식이 관찰됩니다. 큰 종은 작은 종을 잡아 먹습니다. 포식 (그림 112)은 식물 입자를 먹는 동일한 종의 능력과 결합됩니다.

섬모는 어떻게 반추동물의 반추위에 들어가나요? ofrioscolecides의 감염 경로는 무엇입니까? 신생아 반추 동물의 반추위에는 섬모가 아직 없는 것으로 나타났습니다. 동물이 우유를 먹는 동안에도 그들은 결석합니다. 그러나 반추 동물이 식물성 식품으로 전환하자마자 섬모가 흉터와 메쉬에 즉시 나타나며 그 수가 급격히 증가하고 있습니다. 그들은 어디에서 왔니? 오랫동안 반추위 섬모는 일종의 휴지기(대부분 낭포)를 형성하며, 이는 자연계에 널리 퍼져 있으며 삼키면 섬모의 활성 단계를 유발한다고 가정했습니다. 추가 연구에 따르면 반추동물 섬모에는 휴지기가 없습니다. 새김질을 할 때 구강을 관통하는 활동적인 이동 섬모로 감염이 발생함을 증명할 수 있었습니다. 에서 찍은 현미경으로 관찰하면 구강껌이지만 항상 많은 수의 활발하게 떠다니는 섬모가 있습니다. 이러한 활성 형태는 풀, 건초(섬모가 있는 타액을 포함할 수 있음) 등과 함께 일반적인 음주용 용기에서 입으로 쉽게 침투할 수 있고 다른 반추동물의 반추위까지 침투할 수 있습니다. 이 감염 경로는 실험적으로 입증되었습니다.
ofrioscolecids에 휴지기가 없다면, 그들이 여전히 우유를 먹고 있을 때 그들을 분리함으로써 "주입되지 않는" 동물을 얻는 것은 분명히 쉽습니다. 성장하는 새끼와 섬모 반추동물 간의 직접적인 접촉을 피하면 어린 동물은 반추위에서 섬모 없이 남겨질 수 있습니다. 이러한 실험은 여러 국가의 여러 과학자에 의해 수행되었습니다. 결과는 명확했습니다. 반추위에서 섬모가 있는 새끼(우유를 먹이는 동안 어미에게서 빼앗음)와 반추동물 사이에 접촉이 없으면 동물은 섬모와 관련하여 불임으로 자랍니다. 그러나 섬모가 있는 동물과 단기간 접촉(공동 급식기, 일반 식수통, 일반 목초지)만으로도 섬모가 불임 동물의 반추위에서 나타날 수 있습니다.
위는 반추위와 그물에 섬모가 전혀 없는 반추동물의 함량에 대한 실험 결과입니다. 이것은 우리가 보았듯이 젊은이를 조기에 격리함으로써 달성됩니다. 실험은 양과 염소에 대해 수행되었습니다.
이러한 방식으로 상당한 기간(1년 이상) 동안 "주입 장치가 없는" 동물을 관찰하는 것이 가능했습니다. 반추위에서 섬모가 없으면 숙주의 수명에 어떤 영향을 줍니까? 섬모가 없으면 숙주에게 부정적인 영향을 미치거나 긍정적인 영향을 미칩니까? 이 질문에 답하기 위해 염소를 대상으로 다음과 같은 실험을 하였다. 더 유사한 재료를 갖기 위해 쌍둥이 아이(같은 새끼, 같은 성별)를 데려갔습니다. 그런 다음이 쌍의 쌍둥이 중 하나는 반추위에서 섬모없이 자랐고 (조기 격리), 다른 하나는 식물성 식품을 먹기 시작했을 때부터 많은 유형의 섬모에 풍부하게 감염되었습니다. 둘 다 똑같은 식단을 받았고 동일한 조건에서 자랐습니다. 그들 사이의 유일한 차이점은 섬모의 존재 여부였습니다. 이러한 방식으로 연구된 여러 쌍의 어린이에서 각 쌍의 두 구성원("주입기" 및 "비주입기")의 발달 과정에서 차이가 발견되지 않았습니다. 따라서 반추위와 그물에 서식하는 섬모류는 숙주 동물의 생명 기능에 예리한 영향을 미치지 않는다고 주장할 수 있습니다.
그러나 위의 실험 결과는 반추위의 섬모가 소유자에게 완전히 무관심하다고 주장하는 것을 허용하지 않습니다. 이 실험은 숙주의 정상적인 식단으로 수행되었습니다. 다른 조건에서 다른식이 요법 (예 : 불충분 한 먹이)으로 반추위에 서식하는 원생 동물 동물군의 숙주에 미치는 영향을 밝힐 수 있습니다.
반추위의 원생동물 동물군이 숙주의 소화 과정에 긍정적인 영향을 미칠 수 있다는 가능성에 대해 문헌에서 다양한 제안이 있었습니다. 반추위에서 활발하게 헤엄치고 식물 조직을 부수는 수백만 개의 섬모가 소화관의 앞쪽 부분에 위치한 음식 덩어리의 발효와 소화에 기여한다는 것이 지적되었습니다. 츄잉껌과 함께 abomasum으로 들어가는 상당수의 섬모가 소화되고 섬모체의 상당 부분을 구성하는 단백질이 흡수됩니다. 따라서 Infusoria는 숙주에 대한 추가 단백질 공급원이 될 수 있습니다. 섬모는 또한 반추동물의 음식의 대부분을 구성하는 섬유소의 소화와 소화가 잘 되는 상태로의 전환에 기여한다고 제안되었습니다.
이러한 가정은 모두 입증되지 않았으며 일부에 대해 반론이 제기되었습니다. 예를 들어 섬모는 숙주의 음식과 함께 반추위에 들어가는 단백질로부터 몸의 원형질을 만든다는 것이 지적되었습니다. 식물성 단백질을 흡수하여 체내의 동물성 단백질로 전환한 다음, 소화액에서 소화됩니다. 이것이 호스트에게 어떤 이점을 제공하는지 여부는 불분명합니다. 우리는 축산의 주요 대상인 반추동물의 소화에 대해 이야기하고 있기 때문에 이러한 모든 문제는 실제적으로 매우 중요합니다. 반추동물의 소화에서 반추위 섬모의 역할에 대한 추가 연구가 매우 바람직합니다.
반추 동물의 Ofrioscolecides는 일반적으로 광범위한 특이성을 가지고 있습니다. 종으로 보면 소, 양, 염소의 흉터와 그물 개체군이 매우 가깝습니다. 아프리카 영양 반추위의 종 구성을 소와 비교하면 여기에서도 전체 종의 약 40 %가 공통적입니다. 그러나 영양 또는 사슴에서만 발견되는 많은 종류의 리오스코레시드(rioscolecid)가 있습니다. 따라서 rioscolecides의 일반적이고 광범위한 특이성을 배경으로 별도의 더 좁게 특정 유형에 대해 말할 수 있습니다.
말의 창자의 주입
이제 큰 말과 맹장에 서식하는 섬모에 대해 간략히 알아보도록 하겠습니다.
종의 측면에서 반추 동물의 반추 동물과 마찬가지로이 동물 군도 매우 다양합니다. 현재 약 100종의 섬모가 말 가족 동물의 대장에 서식하는 것으로 설명되었습니다. 다른 체계적인 그룹에 속한다는 의미에서 여기에서 발견되는 섬모는 반추 동물의 반추위 섬모보다 더 다양합니다.

말의 내장에는 등섬모목에 속하는 섬모 종류가 꽤 있습니다. 즉, 모양체 기구가 구강 부근에서 막이나 간경변을 형성하지 않는 섬모입니다(그림 113, 1).
엔토디니오모프 주문(Etodiniomorpha)는 또한 말의 내장에 풍부하게 나타납니다. 반추동물의 반추위에서는 단 하나의 endodiniomorphs(ofrioscolecid family) 과만이 발견되는 반면, 3개의 과의 대표자는 말의 창자에 살고 있지만 여기에서 그 특징에 대해서는 언급하지 않고 몇 가지 그림으로 제한합니다. 전형적인 말 종(그림 113) .
A. Strelkov의 자세한 연구에 따르면 다양한 유형의 섬모가 말의 대장을 따라 고르게 분포되어 있지 않습니다. 두 개의 다른 그룹의 종, 두 개의 동물군이 있습니다. 그들 중 하나는 맹장과 대장의 복부 부분(대장의 초기 부분)에 서식하고 다른 하나는 대장의 등쪽 부분과 작은 대장에 서식합니다. 이 두 종의 복합 단지는 다소 뚜렷하게 구분됩니다. 이 두 부분에 공통적인 종은 거의 없습니다. 12개 미만입니다.
, 
말의 대장에 서식하는 수많은 섬모류 종 중에서 섬모류를 빨아먹는 것과 관련된 한 속의 대표자가 있다는 점은 흥미롭습니다. 위에서 보았듯이, 빨기 섬모(Suctoria)는 촉수의 도움으로 매우 특별한 먹이를 주는 전형적인 자유 생활 정착 유기체입니다(그림 103). 중 하나 출산 수술예를 들어 말의 대장과 같은 겉보기에는 특이한 서식지에 적응 알란토시스(알란토소마). 이 매우 독특한 동물(그림 114)은 줄기가 없고 섬모가 없으며 끝이 두꺼워진 곤봉 모양의 촉수가 잘 발달되어 있다.
촉수의 도움으로 알란토솜은 다양한 유형의 섬모에 달라붙어 빨아냅니다. 종종 먹이는 포식자보다 몇 배나 더 큽니다.
말의 대장 섬모와 숙주 사이의 관계의 본질에 대한 질문은 여전히 불분명합니다. 섬모의 수는 반추동물의 반추위만큼 높을 수 있으며 때로는 더 많을 수 있습니다. 말의 대장에 있는 섬모의 양이 1cm3에서 300만 개에 달할 수 있다는 데이터가 있습니다. 일부 과학자들이 제안한 공생적 중요성은 반추위 섬모보다 가능성이 훨씬 적습니다.
가장 가능성있는 의견은 상당한 양의 음식을 흡수하여 숙주에게 약간의 해를 끼친다는 것입니다. 섬모의 일부는 배설물과 함께 제거되므로 몸을 구성하는 유기 물질(단백질 포함)은 숙주가 사용하지 않은 채로 남아 있습니다.
대장에 서식하는 섬모로 말을 감염시키는 방법에 대한 질문은 아직 해결되지 않았습니다.

Balantidia는 대장의 내용물에서 다양한 음식 입자를 포착합니다. 특히 기꺼이 그는 전분 곡물을 먹습니다. 발란티디아가 인간 결장의 내강에 살면 장의 내용물을 먹고 해로운 영향을 미치지 않습니다. 이것은 이질 아메바를 고려할 때 이미 만난 전형적인 "마차"입니다. 그러나 발란티디아는 이질성 아메바보다 그러한 "무해한 거주자"로 남을 가능성이 적습니다.
현재 전문가들은 잘 발달되어 있습니다. 다양한 방법, 숙주 유기체 외부의 인공 환경에서 balantidia의 재배를 허용합니다.

그림에서 알 수 있듯이 troglodptella는 복잡한 endodiniomorphs 중 하나입니다. 그녀는 간경변의 주변 영역 (신체의 앞쪽 끝 부분) 외에도 섬모의 몸을 고리 모양으로 덮는 잘 발달 된 간경변 영역이 3 개 더 있습니다. Trogloditells는 탄수화물로 구성된 잘 발달 된 골격 장치를 가지고 있으며 신체의 거의 전체 앞쪽 끝을 덮습니다. 이 독특한 섬모의 크기는 상당히 중요합니다. 길이는 200-280 미크론에 이릅니다.
Mouthless Ciliates ASTOMATS

지지하는 골격 형성은 주로 신체의 앞쪽 끝에서 발달하며, 이는 기계적 스트레스를 경험하고 장애물을 극복해야 하며 음식물 입자 사이의 장 내강을 통과해야 합니다. 종 속 방사선 사증(Radiophrya) 신체의 한 쪽 앞쪽 끝(일반적으로 복부 쪽으로 간주됨)에는 외질의 표층에 매우 강한 탄성 늑골(spicules)이 있습니다(그림 117, B, D, E) . 종 메닐렐라 속(Mesnilella) 또한 지지선(spicules)이 있는데, 그 길이의 대부분은 세포질의 더 깊은 층에 있습니다(소포체, 그림 117, A). 유사하게 배열된 지지 구조물은 다른 Astomat 속의 종에서도 발달합니다.
일부 섬모류 astomat의 무성 생식은 독특한 방식으로 진행됩니다. 대부분의 섬모의 특징인 가로 분할 대신에 많은 astomat는 고르지 않은 분할(출아)을 가지고 있습니다. 동시에, 후단부에서 분리된 신장은 일정 기간 동안 모체와 관련된 상태를 유지합니다(그림 117, B). 결과적으로 앞쪽의 큰 개체와 뒤쪽의 작은 개체(신장)로 구성된 사슬이 얻어집니다. 앞으로 신장은 점차 사슬에서 분리되어 독립적인 존재로 나아갑니다. 이 독특한 형태의 번식은 예를 들어 이미 우리에게 알려진 방사선 사증에서 널리 퍼져 있습니다. 싹이 트기 때문에 생긴 일부 기공의 사슬은 다음과 유사합니다. 모습촌충의 사슬. 여기서 우리는 수렴 현상을 다시 만난다.
astomat의 핵 장치는 섬모의 구조 특성을 가지고 있습니다 : 대핵, 가장 자주 리본 모양의 모양 (그림 117) 및 하나의 소핵. 수축성 공포는 일반적으로 잘 발달되어 있습니다. 대부분의 종에는 하나의 세로 줄에 배열된 여러(때로는 12개 이상) 수축성 액포가 있습니다.
다른 유형의 기주에 의한 Astomat 종의 분포에 대한 연구에 따르면 대부분의 Astomat 종은 엄격하게 정의된 기주 종에 국한되어 있습니다. 대부분의 astomats는 좁은 특이성을 특징으로합니다. 한 종의 동물 만이 호스트 역할을 할 수 있습니다.
astomat 섬모 연구에 대한 많은 연구에도 불구하고 생물학의 한 가지 매우 중요한 측면은 완전히 불분명합니다. 어떻게 이러한 섬모가 한 숙주 개체에서 다른 개체로 전염됩니까? 이 섬모에서 낭종의 형성을 관찰하는 것은 결코 가능하지 않았습니다.
따라서 감염이 활발하게 발생하는 것이 좋습니다 - 이동 단계.
성게의 내장
성게는 우리 북부 (바렌츠)와 극동 바다 (일본해, 쿠릴 열도의 태평양 연안)의 해안 지역에서 매우 많습니다. 다수 성게주로 해조류와 같은 식물성 식품을 먹으며 입을 벌리고 있는 특별한 날카로운 "이빨"이 있는 수중 물체를 긁습니다. 이 초식성 고슴도치의 내장에는 섬모류의 풍부한 동물군이 있습니다. 종종 그들은 대량으로 여기에서 발생하며 현미경으로 볼 때 성게의 내장 내용물은 반추 동물의 반추위 내용물만큼 섬모로 "충만"합니다. 성게의 내장 섬모와 반추 동물의 반추위의 생활 조건의 깊은 차이점 외에도 몇 가지 유사점이 있다고 말해야합니다. 그들은 여기 저기 섬모가 식물 잔해가 매우 풍부한 환경에서 산다는 사실에 있습니다. 현재 성게의 장에는 50종 이상의 섬모류가 서식하는 것으로 알려져 있으며, 성게가 해조류를 잡아먹는 해안가에서만 볼 수 있다. 조류가 더 이상 자라지 않는 깊은 곳에서는 성게에 섬모가 없습니다.

성게의 내장 섬모는 대부분 생활 방식과 영양의 성질에 따라 초식성입니다. 그들은 숙주의 내장을 대량으로 채우는 조류를 먹습니다. 일부 종은 음식 선택에 있어 상당히 "까다롭습니다". 예를 들어, 스트로빌리듐(Strobilidium, 그림 118, A)는 거의 독점적으로 큰 규조류를 먹습니다. 다른 동물의 대표자를 잡아먹는 포식자도 있습니다. 작은 종섬모.
성게의 창자에서 나온 섬모에는 astomat와 달리 특정 유형의 숙주에 대한 엄격한 제한이 없습니다. 그들은 조류를 먹는 성게의 다양한 종류에 살고 있습니다.
성게가 섬모에 감염되는 방법은 연구되지 않았습니다. 그러나 여기서는 높은 확률로 활성 자유 부동 형태로 발생한다고 가정할 수 있습니다. 사실 성게의 창자에서 나온 섬모는 바닷물에서 오랫동안(수시간) 살 수 있습니다. 그러나 그들은 이미 고슴도치의 내장 생활에 너무 많이 적응하여 몸 밖에서 바닷물에서 조만간 죽습니다.
섬모류에 대한 소개를 마치면서, 섬모가 동물 세계의 종의 풍부하고 광범위하며 번영하는 그룹(계급)을 대표한다는 점을 다시 한 번 강조해야 합니다. 세포 조직의 수준에 남아있는 infusoria는 다른 종류의 원생 동물과 비교할 때 구조와 기능이 가장 복잡합니다.
이러한 점진적 발전(진화)에서 특히 중요한 역할은 아마도 핵 장치의 변형과 핵 이원론(핵의 질적 비등가)의 출현에 의해 수행되었을 것입니다. 핵산 물질에서 거대핵의 풍부함은 세포질과 핵에서 단백질 합성의 활발한 과정과 함께 활성 대사 과정과 관련이 있습니다.
결론
우리는 광범위한 유형의 동물 세계의 구조와 생활 방식에 대한 검토를 마쳤습니다. 원생 동물문. 위에서 반복적으로 강조했듯이 그들의 특징은 단세포성입니다. 구조면에서 원생동물은 세포입니다. 그러나 다세포 유기체의 몸을 구성하는 세포와는 비교할 수 없습니다. 그 자체가 유기체이기 때문입니다. 따라서 원생동물은 조직의 세포 수준에 있는 유기체입니다. 많은 핵을 소유하고 있는 고도로 조직화된 일부 원생동물은 이미 세포 구조의 형태학적 한계를 넘어선 것으로 보이며, 일부 과학자들은 그러한 원생동물을 "세포상(supracellular)"이라고 부르는 이유를 제공합니다. 단세포 조직이 여전히 원생동물의 전형이기 때문에 이것은 문제의 본질을 거의 변경하지 않습니다.
단세포성의 한계 내에서, 원생동물은 진화적 발달의 먼 길을 왔고 가장 다양한 삶의 조건에 적응된 매우 다양한 형태를 제공했습니다. 원생동물의 가계 줄기의 중심에는 두 가지 종류가 있습니다. 이러한 클래스 중 어느 것이 더 원시적인 것인지에 대한 질문은 여전히 과학에서 논쟁 중입니다. 한편으로는 Sarcodidae (ameba)의 하위 대표자가 가장 원시적 인 구조를 가지고 있습니다. 그러나 편모는 신진 대사 유형의 가장 큰 가소성을 나타내며 동물과 식물 세계의 경계에 서 있습니다. 일부 유충(예: 유공충)의 수명 주기에는 편모 단계(배우체)가 있으며, 이는 편모류와의 관계를 나타냅니다. 현대 sarcod도 현대 편모도 동물 세계의 초기 진화 그룹이 될 수 없다는 것은 분명합니다. 왜냐하면 그들 자체가 역사적 발전에서 먼 길을 갔고 지구의 현대 생활 조건에 대한 수많은 적응을 개발했기 때문입니다. 아마도 현대 원생동물의 이 두 부류는 진화의 두 줄기로 간주되어야 하며, 이는 오늘날까지 생존하지 못한 고대 형태에서 기원하며, 이는 우리 행성에서 생명체가 발달하는 여명기에 살았습니다.
원생 동물의 추가 진화에서 다른 성격의 변화가 발생했습니다. 그들 중 일부는 조직 수준의 일반적인 증가, 활동의 증가 및 삶의 과정의 강도로 이어졌습니다. 그러한 계통 발생학적(진화적) 변형 중에는 예를 들어 섬모류에서 높은 수준의 완성도에 도달한 운동 및 음식 포획 소기관의 발달이 포함되어야 합니다. 섬모가 편모에 해당하는(상동성인) 소기관이라는 것은 의심의 여지가 없습니다. 편모에서는 몇 가지 예외를 제외하고 편모의 수는 적지만 섬모에서는 섬모의 수가 수천 개에 달합니다. 섬모 장치의 발달은 원생 동물의 활동을 급격히 증가 시켰고 환경과의 관계 형태를 더욱 다양하고 복잡하게 만들고 외부 자극에 대한 반응 형태를 만들었습니다. 분화된 모양체 기구의 존재는 의심할 여지 없이 다른 서식지에 적응한 다양한 형태가 발생하는 섬모류 부류의 점진적인 진화의 주요 원인 중 하나였습니다.
섬모 섬모 장치의 발달은 Acad에 의해 명명된 그러한 진화적 변화의 한 예입니다. 세베르초프 아로모포스. Aromorphoses는 조직의 일반적인 증가, 광범위한 의미의 적응 개발이 특징입니다. 조직의 증가는 유기체의 중요한 활동을 증가시키는 변화로 이해됩니다. 그것들은 그 부분들의 기능적 분화와 연관되어 유기체와 환경 사이에 보다 다양한 형태의 의사소통을 유도합니다. 섬모 섬모 장치의 발달은 진화 과정에서 이러한 종류의 구조적 변형을 정확하게 나타냅니다. 이것은 전형적인 아로모포시스입니다.
V. A. Dogel이 강조한 바와 같이, 원생동물에서 기형 유형의 변화는 일반적으로 소기관 수의 증가와 관련이 있습니다. 소기관의 중합이 발생합니다. 섬모에서 섬모 기구의 발달은 그러한 변화의 전형적인 예입니다. 섬모의 진화에서 aromorphosis의 두 번째 예는 그들의 핵 장치입니다. 우리는 섬모 코어의 구조적 특징을 위에서 조사했습니다. 섬모의 핵 이원성(소핵과 거대핵의 존재)은 거대핵의 염색체 수의 증가(배수성 현상)를 동반했습니다. 염색체는 주로 단백질 합성과 함께 세포의 주요 합성 과정과 관련되어 있기 때문에 이 과정은 주요 생명 기능의 강도를 전반적으로 증가시켰습니다. 여기에서도 중합이 일어나 핵의 염색체 복합체에 영향을 미쳤습니다.
섬모- 가장 많고 진보적인 원생동물 그룹 중 하나는 편모류에서 유래합니다. 이것은 운동 오르가노이드의 완전한 형태학적 유사성에 의해 입증됩니다. 이 진화 단계는 두 가지 큰 형태와 관련이 있습니다. 그 중 하나는 운동 소기관에 영향을 미치고 두 번째는 핵 장치에 영향을 미칩니다. 이러한 두 가지 유형의 변화는 모두 상호 연결되어 있습니다. 둘 다 중요한 활동의 증가와 외부 환경과의 상호 관계 형태의 복잡성으로 이어지기 때문입니다.
aromorphoses와 함께, 명확하게 정의된 특정 존재 조건에 대한 적응(적응)의 발달로 표현되는 또 다른 유형의 진화적 변화가 있습니다. 이러한 유형의 진화적 변화를 Severtsov는 idioadaptation이라고 불렀습니다. 원생동물의 진화에서 이러한 유형의 변화는 매우 중요한 역할을 했습니다. 위에서 고려할 때 다른 수업 protozoa, idioadaptive 변화의 수많은 예가 제공됩니다. 다양한 원생 동물 그룹의 플랑크톤 생활 방식에 대한 적응, 섬모류의 모래 생활에 대한 적응, 콕시듐에서 난모낭의 보호 껍질 형성 등 - 이 모든 것이 출현과 발달에 큰 역할을 한 특발성 적응입니다. 개별 그룹의 그룹이지만 조직의 일반적인 점진적 변화와 관련이 없습니다.
원생동물의 다양한 특정 서식지에 대한 적응은 매우 다양합니다. 그들은 개별 클래스를 설명할 때 위에서 자세히 논의한 다양한 서식지에서 이 유형의 광범위한 분포를 보장했습니다.
큰 의학 백과사전
Infusoria Tetrahymena thermophila ... Wikipedia
-(Infusoria) 가장 고도로 발달된 원생동물(Protozoa)의 클래스. I.의 주요 특징 : 섬모의 존재 (운동 및 영양), 두 가지 유형의 핵 (다배체 거대 핵 및 이배체 소핵, 구조가 다르며 ... ... 위대한 소비에트 백과사전
또는 액체 동물(Infusoria)은 반짝이는 땋은 모양이나 섬모가 있는 원생 동물의 한 종류로, 때로는 성인기에 원통형 빨기 파생물로 대체되며 대부분의 경우 ... ... 백과사전에프. 브로크하우스와 I.A. 에프론
특히 원생동물의 분류학의 일반적 개념에 따른 섬모류 종류에 속하는 유기체의 분류 및 체계화 및 동물 일반의 분류법 Contents 1 Systematics 1.1 Classical ... Wikipedia
또는 액체 동물 (Infusoria) 반짝이는 엮기 또는 섬모가 장착 된 원생 동물의 부류로, 성인 상태에서 때로는 원통형 빠는 파생물로 대체되며 대부분의 경우 ... ... Brockhaus와 Efron의 백과사전
섬모의 유형은 일반적으로 섬모라고도 합니다. 이 원생동물의 운동 기관은 다음과 같습니다. 속눈썹. 섬모 세포에는 두 개의 핵이 있으며 이를 소형 및 대형이라고 합니다. 첫 번째는 번식 과정을 조절하고 두 번째는 영양, 운동 및 호흡 과정을 담당합니다.
이 유형의 삶의 특징은 섬모 신발의 예에서 고려해야합니다.
움직임과 호흡
길이가 약 0.5mm인 infusoria-shoe는 수역을 서식지로 선택합니다. 가장 단순한 몸체의 모양은 이름으로 추측하기 쉽습니다. 신발과 비슷합니다. 이동 속도는 초당 약 2.5mm입니다.
외부 탄성 쉘의 존재는 안정적인 체형을 제공합니다.
껍질에 인접한 세포질에는지지 섬유가 있으며 발달은 섬모의 일정한 형태를 보존하는 것을 보장합니다.
섬모의 표면에는 15,000개의 섬모가 있고, 그 바닥에는 기초체가 있습니다. 움직임은 섬모의 진동의 도움으로 발생합니다. 그들은 초당 약 30 스트로크를 생성하여 섬모 신발을 앞으로 밀어냅니다.
그것은 신체의 표면을 통해 호흡합니다.
음식
섬모의 특징은 존재입니다 세포 입, 특히 길고 빽빽한 섬모가 있습니다. 세포 인두는 세포 인두와 함께 계속됩니다. 섬모는 물과 섬모 음식을 밀어 넣습니다. 박테리아.
주입은 화학 물질을 감지하여 박테리아 클러스터를 방출합니다. 따라서 그녀는 먹이를 찾습니다.
그런 다음 음식은 소화 액포에서 끝납니다. 여기에서 이미 세포질로 이어집니다.
선택
선택은 다음을 사용하여 수행됩니다. 두 개의 수축성 공포, 하나는 전면 끝에 있고 다른 하나는 후면에 있습니다. 액포는 저장소와 채널로 구성됩니다.
액체는 채널을 채우고 중앙 저장소를 따라간 다음 주입기를 빠져 나옵니다. 액포 수축 과정은 10-20초가 걸립니다.
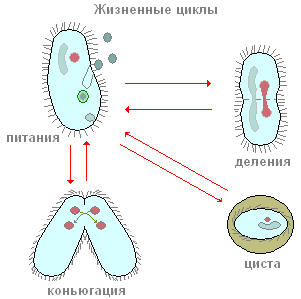
생식
infusoria는 무성으로 번식합니다. 두 개로 나뉩니다. 그 특징은 몸 전체에 분할입니다.
섬모의 핵은 두 부분으로 나뉩니다. 새로 형성된 섬모에는 크고 작은 핵이 있습니다. 딸 섬모는 오르가노이드의 일부를 소유하고 누락된 것은 독립적으로 형성됩니다. 번식은 하루에 여러 번 발생합니다.
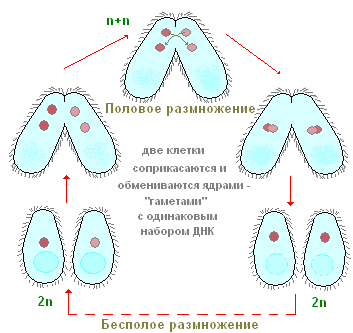
섬모 신발의 경우 유성 생식도 가능하지만이 경우 개체 수가 증가하지 않습니다. 일시적으로 원생동물이 연결되어 세포질에서 연결 다리를 형성합니다.
각 개인에서 큰 핵은 사라지고 작은 핵은 두 번 나누어집니다. 4 개의 핵이 나타납니다. 이 중 하나의 핵만 남고 이 핵도 분열합니다. 개인은 두 개의 핵을 포함하고 핵 교환이 발생합니다. 핵 중 하나가 다른 개인으로 이동합니다.
거기에서 그것은 남아있는 핵과 합쳐지고, 따라서 각각의 개체에 크고 작은 핵이 형성됩니다. 라고 하는 이 과정을 동사 변화, 개인 간의 유전 물질 재생에 필요합니다.
섬모의 종류
섬모는 복잡하게 조직된 원생동물이며 약 7,000종이 있습니다.
1. 다양한 sarcodes.
2. 다양한 편모.
두 번째 하위 유형의 대표자는이 그룹의 이름에 반영된 편모의 도움으로 움직입니다.
- 편모. 유글레나 그린은 이 그룹의 대표적인 대표자입니다. 이 원생동물은 조밀한 탄성막으로 덮인 방추형을 가지고 있습니다. 몸의 앞쪽 끝에는 편모가 있으며 그 기저부 근처에는 빛에 민감한 눈이 있습니다 - 낙인과 수축성 액포. 후단에 가까울수록 세포질의 두께에는 신체의 모든 중요한 과정을 제어하는 핵이 있습니다. 유글레나의 녹색은 수많은 엽록체 때문입니다. 빛에서 이 편모는 광합성을 하지만 저수지의 조명이 없는 지역에 오랫동안 있으면 복잡한 유기 물질의 부패 생성물을 먹고 환경에서 추출합니다. 따라서 녹색 유글레나는 독립 영양 유형의 신진 대사와 종속 영양 유형을 모두 결합 할 수 있습니다. 이렇게 혼합된 형태의 영양을 가진 유기체의 존재는 동식물 세계의 관계를 나타냅니다. 대부분의 편모
3. 다양한 포자동물과 섬모.
섬모류 유형에는 단세포 동물 중에서 가장 복잡한 조직으로 구별되는 원생 동물이 포함됩니다. 이 그룹의 전형적인 대표자는 담수의 일반적인 거주자 인 infusoria-shoe입니다. 그녀의 몸은 조밀한 껍질 - 펠리클로 덮여 있으므로 비교적 일정한 모양을 가지고 있습니다. 구조의 특징은 신발의 몸 전체를 고르게 덮는 섬모가 있다는 것입니다. 섬모의 움직임은 세포질의 표층에 위치한 수축성 섬유 네트워크로 인해 조정됩니다. 두 번째 특징은 큰(거대핵)과 작은(소핵) 두 개의 핵이 있다는 것입니다. 핵은 기능적으로도 다릅니다. 큰 핵은 신진대사를 조절하고 작은 핵은 성 과정(접합)에 참여합니다. infusoria-shoe는 박테리아인 단세포 조류를 먹고살며, 이 조류는 섬모에 의해 구강 내 바닥에 있는 세포 입으로 조정됩니다. 세포질로 끝나는 세포 인두를 통과한 후 음식물 입자는 소화액포에 둘러싸여 효소의 작용에 의해 분해됩니다. 소화되지 않은 잔류 물은 분말을 통해 버려집니다. 신발의 몸체에는 두 개의 교대로 맥동하는 복합 수축 액포가 있습니다. 신발은 반으로 나누어 무성생식한다. 여러 개의 무성 생식은 두 개의 신발이 함께 와서 유전 물질을 교환하는 성적인 과정 - 접합으로 신발에서 대체됩니다. 그 후, 신발은 갈라지고 곧 다시 무성 생식을 시작합니다. 접합의 생물학적 중요성은 두 개인의 유전 특성이 한 유기체에 결합되어 있다는 것입니다. 이것은 환경 조건에 대한 더 나은 적응력으로 표현되는 생존력을 증가시킵니다.
찾고 있는 것을 찾지 못하셨나요? 검색 사용
이 페이지에서 주제에 대한 자료:
- 편모류, 위족류, 포자동물 및 섬모류에 대해 간략하게
- sarcodes 및 sporozoans
- 다양한 편모
- 인퓨소리아 슬리퍼 리포트 요약
- 섬모와 sarcode 종의 차이점은 무엇입니까
섬모의 가장 전형적인 잘 알려진 대표자 중 하나는 섬모 신발입니다. 그것은 원칙적으로 서있는 방향의 물과 담수 형 저수지에서 살고 있습니다.이 저수지에서는 현재가 독단성을 제외하고 구별됩니다. 서식지는 반드시 부패하는 유기물을 포함해야 합니다. 이 동물 군 대표의 삶의 모든 측면을 자세히 고려하는 것이 좋습니다.
속눈썹의 대표자

섬모는 "팅크"라는 단어에서 이름이 유래한 유형이라는 점에 유의해야 합니다. 라틴어). 이것은 원생 동물의 첫 번째 대표자가 초본 팅크에서 정확하게 발견되었다는 사실로 설명 할 수 있습니다. 시간이 지남에 따라 이러한 유형의 개발이 빠르게 추진력을 얻기 시작했습니다. 따라서 이미 오늘날 생물학에서는 섬모의 유형을 포함하여 약 6-7,000종이 알려져 있습니다. 1980년대의 데이터에 의존한다면 문제의 유형이 구조에 두 개의 클래스를 포함한다고 주장할 수 있습니다. 이 정보와 관련하여 우리는 살아있는 유기체의 다양성이 매우 광범위하다는 결론을 내릴 수 있으며 이는 진정한 관심입니다.
Infusoria의 유형: 대표자

이 유형의 저명한 대표자는 섬모-발란티디아 및 섬모-신발입니다. 고유 한 특징이 동물들은 이동에 사용되는 섬모로 펠리클을 덮고, 특별히 설계된 기관, trichocysts (껍질의 세포질에 위치)를 통한 섬모 보호 및 두 개의 핵 세포 (식물성 및 생성). 또한 섬모 몸체의 구강은 구강 깔때기를 형성하여 인두로 이어지는 세포 입으로 들어가는 경향이 있습니다. 음식의 소화에 직접 작용하는 소화 액포가 생성되는 곳입니다. 그러나 소화되지 않은 성분은 분말을 통해 몸에서 제거됩니다. 섬모 유형의 특성매우 다양하지만 주요 사항은 위에서 논의했습니다. 추가해야 할 유일한 것은 두 개의 섬모가 신체의 반대쪽에 있다는 것입니다. 과도한 수분이나 대사 산물이 신체에서 배설되는 것은 그들의 기능을 통해서입니다.
인퓨소리아 신발

이러한 단세포 구조의 흥미로운 유기체의 구조와 생활 방식을 질적으로 고려하려면 해당 예를 참조하는 것이 좋습니다. 이를 위해서는 담수 저수지에 널리 퍼져있는 infusoria-shoes가 필요합니다. 그들은 일반 용기 (예 : 수족관)에서 쉽게 자랄 수 있으며 초원 건초 만이 가장 쉽습니다. 민물,이 유형의 팅크에서는 원칙적으로 섬모 신발을 포함하여 많은 종의 원생 동물이 발생하기 때문입니다. 따라서 현미경을 통해 기사에서 제공하는 모든 정보를 실제로 연구할 수 있습니다.
섬모 신발의 특성
위에서 언급했듯이 섬모는 많은 요소를 포함하는 유형이며 그 중 가장 흥미로운 것은 섬모 신발입니다. 이것은 길이가 반 밀리미터이며 스핀들 모양이 부여됩니다. 시각적으로이 유기체는 신발과 비슷하므로 흥미로운 이름입니다. infusoria-shoe는 끊임없이 움직이는 상태에 있으며 뭉툭한 끝을 앞으로 수영합니다. 흥미롭게도 이동 속도는 종종 초당 2.5mm에 도달하며 이는 이러한 유형의 대표자에게 매우 좋습니다. 섬모 신발의 몸체 표면에는 운동 소기관 역할을 하는 섬모가 관찰될 수 있습니다. 모든 섬모류와 마찬가지로 해당 유기체는 구조에 두 개의 핵이 있습니다. 큰 핵은 영양, 호흡, 운동 및 대사 과정을 담당하고 작은 핵은 성적 측면에 참여합니다.
섬모 신발의 몸

섬모 신발의 구조는 매우 복잡합니다. 이 대표자의 외부 코팅은 얇은 탄성 쉘입니다. 평생 동안 신체의 올바른 형태를 유지할 수 있습니다. 이것의 충실한 조수는 막에 단단히 인접한 세포질 층에 위치한 완벽하게 발달 된지지 섬유입니다. 섬모 신발의 몸체 표면에는 외부 환경에 관계없이 변동하는 엄청난 수(약 15,000)의 섬모가 있습니다. 그들 각각의 기초에는 기초 몸체가 있습니다. 섬모는 몸을 앞으로 밀기 위해 초당 약 30번 움직입니다. 이 악기의 물결 모양의 움직임은 매우 조화롭게 이루어지므로 infusoria가 움직임의 과정에서 몸의 세로 축을 중심으로 천천히 그리고 아름답게 회전할 수 있습니다.
섬모 - 확실한 관심 유형
섬모 신발의 모든 기능을 절대적으로 이해하려면 수명의 주요 과정을 고려하는 것이 좋습니다. 따라서 박테리아와 조류를 먹는 것으로 귀결됩니다. 유기체의 몸에는 세포 입이라고하는 오목한 부분이 부여되어 인두로 전달되며 바닥에서 음식이 액포로 직접 들어갑니다. 그곳에서 약 1시간 동안 소화되어 그 과정에서 산성 환경에서 알칼리성 환경으로 전환됩니다. 액포는 세포질의 흐름을 통해 섬모체의 몸에서 움직이고, 소화되지 않은 잔류물은 분말을 통해 몸의 뒤쪽으로 나간다.
섬모 신발의 호흡은 신체의 외피를 통해 세포질에 들어가는 산소를 통해 수행됩니다. 그리고 배설 과정은 두 개의 수축성 액포를 통해 발생합니다. 유기체의 과민성에 관해서는 섬모-신발은 박테리아에 의해 분비되는 물질의 작용에 반응하여 박테리아 복합체로 조립되는 경향이 있습니다. 그리고 그들은 식탁용 소금과 같은 자극제로부터 멀리 떠납니다.
생식

신발 섬모는 두 가지 방법 중 하나로 번식할 수 있습니다. 무성 생식은 핵이 두 부분으로 나뉘는 것에 따라 더 널리 퍼졌습니다. 이 작업의 결과로 각 섬모에는 2개의 핵(큰 것과 작은 것)이 있습니다. 유성 생식은 영양 결핍이 있거나 동물 신체의 온도 체계에 변화가 있을 때 적절합니다. 그 후에 infusoria가 낭종으로 변할 수 있다는 점에 유의해야합니다. 그러나 성적 유형의 번식으로 개인 수의 증가는 제외됩니다. 따라서 두 개의 섬모가 일정 시간 동안 서로 연결되어 껍질이 용해되고 동물 사이에 연결 다리가 형성됩니다. 각각의 큰 핵은 흔적도 없이 사라지고 작은 것은 두 번 핵분열 과정을 거치는 것이 중요하다. 따라서 각 섬모에는 4 개의 딸 핵이 형성되고 그 후 3 개는 파괴되고 네 번째는 다시 분할됩니다. 이 성적인 과정을 접합이라고 합니다. 지속 시간은 최대 12시간입니다.
infusoria-shoe는 작은 고인 저수지에 산다. 길이 0.5mm의 이 단세포 동물은 방추 모양의 몸체를 가지고 있으며 어렴풋이 신발과 비슷합니다. 섬모는 끊임없이 움직이며 뭉툭한 끝을 앞으로 내밀고 있습니다. 이 동물의 이동 속도는 초당 2.5mm에 이릅니다. 몸의 표면에는 운동 소기관인 섬모가 있습니다. 세포에는 두 개의 핵이 있습니다. 큰 핵은 영양, 호흡, 운동, 신진 대사를 담당합니다. 작은 핵은 성적인 과정에 관여합니다.
신발 인퓨소리아의 구조
섬모의 유기체는 더 복잡합니다. 섬모의 외부를 덮는 얇은 탄성 껍질은 몸의 일정한 모양을 유지합니다. 이것은 또한 껍질에 인접한 세포질 층에 위치한 잘 발달된 지지 섬유소에 의해 촉진됩니다. 섬모체의 표면에는 약 15,000개의 진동하는 섬모가 있습니다. 각 섬모의 기저부에는 기초체가 있습니다. 각 속눈썹의 움직임은 한 방향으로의 날카로운 스트로크와 원래 위치로의 느리고 부드러운 복귀로 구성됩니다. 섬모는 1초에 약 30번 진동하며 노처럼 주입부를 앞으로 밀어냅니다. 섬모의 물결 모양의 움직임이 조정됩니다. 섬모 신발은 수영할 때 몸의 세로 축을 중심으로 천천히 회전합니다.
생활 과정
음식
슬리퍼 및 기타 자유 생활 섬모는 박테리아와 조류를 먹습니다.
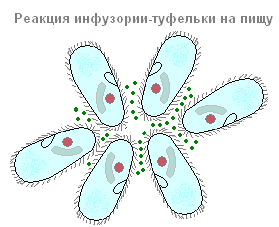
섬모-신발의 음식 반응
얇은 탄성 쉘, ( 세포막) 섬모를 외부에서 덮고 일정한 체형을 유지합니다. 약 15,000개의 섬모가 신체 표면에 있습니다. 몸에는 세포 인두로 들어가는 세포 입이 있습니다. 인두의 바닥에서 음식은 소화액포로 들어갑니다. 소화 액포에서 음식은 1시간 이내에 소화되며, 처음에는 산성 반응으로, 그 다음에는 알칼리 반응으로 소화됩니다. 소화액포는 세포질의 흐름에 따라 섬모체에서 움직입니다. 소화되지 않은 잔류 물은 입 구멍 뒤에 위치한 분말이라는 특수 구조를 통해 몸의 뒤쪽 끝에 버려집니다.
호흡
호흡은 신체의 외피를 통해 발생합니다. 산소는 신체의 전체 표면을 통해 세포질에 들어가고 복잡한 유기 물질을 산화시켜 결과적으로 물, 이산화탄소 및 기타 화합물로 변합니다. 동시에 동물의 삶에 필요한 에너지가 방출됩니다. 이산화탄소는 호흡하는 동안 신체의 전체 표면을 통해 제거됩니다.
선택
섬모-신발의 몸에는 몸의 앞쪽과 뒤쪽 끝에 위치한 두 개의 수축성 액포가 있습니다. 그들은 복잡한 유기 물질의 산화 중에 형성된 용해 물질로 물을 수집합니다. 한계 값에 도달하면 수축성 액포가 신체 표면에 접근하고 내용물이 쏟아집니다. 민물 단세포 동물에서 과도한 물은 수축성 액포를 통해 제거되며, 이는 환경에서 지속적으로 몸으로 들어갑니다.
과민성
섬모-신발은 그들이 분비하는 물질의 작용에 반응하여 박테리아 덩어리로 모여들지만 식탁용 소금과 같은 자극 물질로부터 멀리 수영합니다.
과민성은 빛, 열, 습기, 화학 물질, 기계적 영향과 같은 자극의 작용에 반응하는 모든 살아있는 유기체의 특성입니다. 과민성 덕분에 단세포 동물은 불리한 조건을 피하고 음식, 올해의 개인을 찾습니다.
생식
성기이 없는
섬모는 일반적으로 둘로 나누어 무성생식한다. 핵은 두 부분으로 나뉘며 각각의 새로운 섬모에는 하나의 큰 핵과 하나의 작은 핵이 있습니다. 두 딸 각각은 소기관의 일부를 받고 나머지는 새로 형성됩니다.
섬모 신발 복제
성적
음식이 부족하거나 온도가 바뀌면 섬모가 유성 생식을 한 다음 낭종으로 변할 수 있습니다.
성적인 과정에서 개인 수의 증가는 발생하지 않습니다. 두 섬모는 일시적으로 서로 연결되어 있습니다. 접촉 지점에서 껍질이 녹고 동물 사이에 연결 다리가 형성됩니다. 각 섬모의 큰 핵이 사라집니다. 작은 핵은 두 번 분열합니다. 각 섬모에는 4개의 딸핵이 형성됩니다. 그 중 셋은 멸망하고, 넷째는 다시 나누어집니다. 결과적으로 각각에 두 개의 코어가 남아 있습니다. 핵 교환은 세포질 다리를 따라 일어나고 거기에서 나머지 핵과 합쳐집니다. 새로 형성된 핵은 크고 작은 핵을 형성하고 섬모는 갈라집니다. 이 성적인 과정을 접합이라고 합니다. 약 12시간 지속됩니다. 성적인 과정은 갱신, 개인 간의 교환 및 유전(유전) 물질의 재분배로 이어져 유기체의 생존력을 증가시킵니다.
섬모 신발의 수명주기





