Devoirs. Chausson d'infusoire
Comparés à d'autres groupes de protozoaires, les ciliés ont la structure la plus complexe, qui est associée à la diversité et à la complexité de leurs fonctions.
D'où vient le nom "pantoufle d'infusoires" ? Vous ne serez pas surpris si vous regardez au microscope un cilié vivant, ou même son image (Fig. 85).
En effet, la forme du corps de ce cilié ressemble à un élégant soulier de dame.
La chaussure infusoria est en mouvement continu assez rapide. Sa vitesse (à température ambiante) est d'environ 2,0-2,5 mm/sec. C'est beaucoup de vitesse pour un si petit animal ! Après tout, cela signifie qu'en une seconde, la chaussure parcourt une distance dépassant de 10 à 15 fois la longueur de son corps. La trajectoire de la chaussure est plutôt compliquée. Elle bouge l'avant tout droit
CHAUSSURE INFUSORIUM (PARAMECIUM CAUDATUM)
Pour se familiariser avec la structure et le mode de vie de ces organismes unicellulaires intéressants, tournons-nous d'abord vers un exemple caractéristique. Prenons les ciliés sabots (espèces du genre Paramecium), très répandus dans les réservoirs d'eau douce peu profonds. Ces ciliés sont très faciles à reproduire dans de petits aquariums, si vous remplissez l'étang avec du foin de prairie ordinaire. Dans de telles teintures, de nombreux types différents de protozoaires se développent et des ciliés de chaussures se développent presque toujours. Avec l'aide d'un microscope éducatif ordinaire, vous pouvez voir une grande partie de ce qui sera discuté plus loin.
Parmi les plus simples chaussures de ciliés sont des organismes assez gros. La longueur de leur corps est d'environ 1/6-1/3 mm. et tourne vers la droite le long de l'axe longitudinal du corps.
Un tel mouvement actif de la chaussure dépend du travail d'un grand nombre des plus beaux appendices ressemblant à des cheveux - des cils qui couvrent tout le corps du cilié. Le nombre de cils chez un individu des ciliés de la chaussure est de 10 à 15 000!
Chaque cil fait des mouvements de pagaie très fréquents - à température ambiante jusqu'à 30 battements par seconde. Pendant le refoulement, le cil est maintenu dans une position redressée. Lorsqu'il revient à sa position d'origine (en descendant), il se déplace 3 à 5 fois plus lentement et décrit un demi-cercle.
Lorsqu'une chaussure nage, les mouvements de nombreux cils recouvrant son corps se résument. Les actions des cils individuels sont coordonnées, ce qui se traduit par des vibrations ondulatoires correctes de tous les cils. L'onde d'oscillation commence à l'avant du corps et se propage vers l'arrière. En même temps, 2-3 vagues de contraction passent le long du corps de la chaussure. Ainsi, l'ensemble de l'appareil ciliaire du cilié est, pour ainsi dire, un ensemble physiologique fonctionnel unique, dont les actions des unités structurelles individuelles (cils) sont étroitement liées (coordonnées) les unes aux autres.
La structure de chaque cil de chaussure, comme le montrent les études au microscope électronique, est très complexe.
La direction et la vitesse de déplacement de la chaussure ne sont pas des quantités constantes et immuables. La chaussure, comme tous les organismes vivants (nous l'avons déjà vu avec l'exemple de l'amibe), réagit à un changement environnement externe changement de sens de déplacement.
Un changement dans la direction du mouvement des protozoaires sous l'influence de divers stimuli est appelé taxis. Chez les ciliés, il est facile d'observer diverses taxis. Si dans une goutte où les chaussures flottent, placez une substance qui les affecte négativement (par exemple, un cristal sel de table), puis les chaussures s'éloignent (comme si elles fuyaient) de ce facteur qui leur est défavorable (Fig. 86).

Nous avons devant nous un exemple de taxis négatif sur un effet chimique (chimiotaxie négative). Vous pouvez observer la chaussure et la chimiotaxie positive. Si, par exemple, une goutte d'eau dans laquelle nagent des ciliés est recouverte d'un couvercle en verre et qu'une bulle de dioxyde de carbone (CO2) est laissée en dessous, alors la plupart des ciliés iront vers cette bulle et s'installeront autour d'elle dans un anneau.
Le phénomène des taxis se manifeste très clairement dans les chaussures sous l'influence d'un courant électrique. Si un faible courant électrique traverse le liquide dans lequel flottent les chaussures, on peut alors observer l'image suivante: tous les ciliés orientent leur axe longitudinal parallèlement à la ligne de courant, puis, comme sur commande, se dirigent vers la cathode, dans la zone où ils forment un amas dense. Le mouvement des ciliés, déterminé par la direction du courant électrique, est appelé galvanotaxie. Diverses taxis chez les ciliés peuvent être détectées sous l'influence d'une grande variété de facteurs environnementaux.
L'ensemble du corps cytoplasmique du cilié est clairement divisé en 2 couches : l'extérieure est plus claire (ectoplasme) et l'intérieure est plus foncée et granuleuse (endoplasme), un rôle important dans le maintien de la constance de la forme corporelle des ciliés.
Dans la couche externe (dans l'ectoplasme) du corps d'une chaussure vivante, de nombreuses tiges courtes sont clairement visibles, situées perpendiculairement à la surface (Fig. 85, 7). Ces formations sont appelées trichocystes. Leur fonction est très intéressante et s'apparente à la protection des plus simples. Avec une irritation mécanique, chimique ou autre forte irritation, les trichocystes sont projetés avec force, se transformant en longs fils minces qui frappent un prédateur attaquant la chaussure. Les trichocystes sont une défense puissante. Ils sont disposés régulièrement entre les cils, de sorte que le nombre de trichocystes correspond approximativement au nombre de cils. A la place des trichocystes utilisés ("shot"), de nouveaux se développent dans l'ectoplasme de la chaussure.
D'un côté, approximativement au milieu du corps (Fig. 85, 5), la chaussure présente une dépression assez profonde. C'est la cavité buccale, ou péristome. Les cils sont situés le long des parois du péristome, ainsi que le long de la surface du corps. Ils se développent ici beaucoup plus puissamment que sur le reste de la surface du corps. Ces cils rapprochés sont disposés en deux groupes. La fonction de ces cils très différenciés n'est pas liée au mouvement, mais à la nutrition (Fig. 87).

Comment et que mangent les chaussures, comment digèrent-elles ?
Les chaussures font partie des ciliés, dont la nourriture principale est la bactérie. En plus des bactéries, ils peuvent également avaler d'autres particules en suspension dans l'eau, quelle que soit leur valeur nutritionnelle. Les cils péribuccaux créent un flux continu d'eau avec des particules en suspension en direction de l'ouverture buccale, située profondément dans le péristome. De petites particules alimentaires (le plus souvent des bactéries) pénètrent par la bouche dans un petit pharynx tubulaire et s'accumulent au fond de celui-ci, à la frontière avec l'endoplasme. L'ouverture de la bouche est toujours ouverte. Ce ne sera peut-être pas une erreur de dire que le sabot cilié est l'un des animaux les plus voraces : il se nourrit en permanence. Ce processus n'est interrompu qu'à certains moments de la vie liés à la reproduction et au processus sexuel.
Le morceau de nourriture accumulé au fond du pharynx se détache alors du fond du pharynx et, avec une petite quantité de liquide, pénètre dans l'endoplasme, formant une vacuole digestive. Ce dernier ne reste pas sur le site de sa formation, mais, tombant dans les courants de l'endoplasme, fait un chemin assez complexe et régulier dans le corps de la chaussure, appelé la cyclose de la vacuole digestive (Fig. 88). Au cours de ce parcours assez long (à température ambiante, environ une heure) de la vacuole digestive, un certain nombre de changements se produisent à l'intérieur de celle-ci associés à la digestion des aliments qu'elle contient.
Ici, comme chez les amibes et certains flagellés, une digestion intracellulaire typique se produit. De l'endoplasme entourant la vacuole digestive, des enzymes digestives y pénètrent, qui agissent sur les particules alimentaires. Les produits de digestion sont absorbés par la vacuole digestive dans l'endoplasme.
Au cours de la cyclose de la vacuole digestive, plusieurs phases de digestion y sont remplacées. Dans les premiers instants après la formation d'une vacuole, le liquide qui la remplit diffère peu du liquide de l'environnement. Bientôt, les enzymes digestives commencent à pénétrer dans la vacuole à partir de l'endoplasme et la réaction de l'environnement à l'intérieur devient fortement acide. Ceci est facile à détecter en ajoutant un indicateur aux aliments, dont la couleur change en fonction de la réaction (acide, neutre ou alcaline) de l'environnement. Dans ce milieu acide, les premières phases de la digestion ont lieu. Ensuite, l'image change et la réaction à l'intérieur des vacuoles digestives devient légèrement alcaline. Dans ces conditions, il y a prochaines étapes digestion intracellulaire. La phase acide est généralement plus courte que la phase alcaline ; elle dure environ 1/6-1/4 du séjour total de la vacuole digestive dans le corps du cilié. Cependant, le rapport des phases acides et alcalines peut varier dans des limites assez larges, selon la nature de l'aliment.
Le chemin de la vacuole digestive dans l'endoplasme se termine par le fait qu'elle s'approche de la surface du corps et à travers la pellicule, son contenu, constitué de résidus alimentaires liquides et non digérés, est rejeté - la défécation se produit. Ce processus, contrairement aux amibes, dans lequel la défécation peut se produire n'importe où, dans les chaussures, comme dans les autres ciliés, est strictement confiné à une zone spécifique du corps située sur la face ventrale (l'abdomen est conventionnellement appelé la surface de l'animal sur lequel est placé le récessus périoral), approximativement à mi-chemin entre le péristome et l'extrémité postérieure du corps.
Ainsi, la digestion intracellulaire est un processus complexe constitué de plusieurs phases se remplaçant successivement.

Les calculs montrent qu'en 30 à 45 minutes environ, un volume de liquide égal au volume du corps cilié est excrété de la chaussure par les vacuoles contractiles. Ainsi, en raison de l'activité des vacuoles contractiles, un flux continu d'eau est effectué à travers le corps du cilié, qui pénètre de l'extérieur par l'ouverture de la bouche (avec les vacuoles digestives), ainsi que par voie osmotique directement à travers la pellicule. Les vacuoles contractiles jouent un rôle important dans la régulation du flux d'eau traversant le corps du cilié et dans la régulation de la pression osmotique. Ce processus se déroule ici en principe de la même manière que chez les amibes, seule la structure de la vacuole contractile est beaucoup plus compliquée.
Pendant de nombreuses années, parmi les scientifiques étudiant les protozoaires, il y a eu un différend sur la question de savoir s'il existe des structures dans le cytoplasme associées à l'apparition d'une vacuole contractile ou si elle se forme à chaque fois. Sur un cilié vivant, aucune structure particulière qui aurait précédé sa formation ne peut être observée. Après la contraction de la vacuole - systole -, aucune structure n'est visible dans le cytoplasme à l'emplacement de l'ancienne vacuole. Ensuite, une bulle transparente ou des canaux adducteurs réapparaissent, qui commencent à augmenter de taille. Cependant, aucun lien entre la vacuole nouvellement émergente et la vacuole précédemment existante n'est trouvé. Il semble qu'il n'y ait pas de continuité entre les cycles successifs de la vacuole contractile et toute nouvelle vacuole contractile se forme à nouveau dans le cytoplasme. Cependant, des méthodes de recherche spéciales ont montré que ce n'est pas réellement le cas. L'utilisation de la microscopie électronique, qui donne un très fort grossissement (jusqu'à 100 000 fois), a montré de manière convaincante que le cilié a un cytoplasme particulièrement différencié dans la zone où se forment les vacuoles contractiles, consistant en un entrelacement des tubes les plus fins. Ainsi, il s'est avéré que la vacuole contractile ne se pose pas dans le cytoplasme sur " lieu vide”, mais sur la base de l'organoïde cellulaire spécial précédent, dont la fonction est la formation d'une vacuole contractile.
Comme tous les protozoaires, les ciliés possèdent un noyau cellulaire. Cependant, en termes de structure de l'appareil nucléaire, les ciliés diffèrent fortement de tous les autres groupes de protozoaires.
L'appareil nucléaire des ciliés se caractérise par son dualisme. Cela signifie que les ciliés ont deux types de noyaux différents - les gros noyaux, ou macronoyaux, et les petits noyaux, ou micronoyaux. Voyons quelle structure a l'appareil nucléaire dans les ciliés de la chaussure (Fig. 85).
Au centre du corps du cilié (au niveau du péristome), un gros noyau massif de forme ovoïde ou en forme de haricot est placé. C'est le macronoyau. À proximité immédiate de celui-ci se trouve un deuxième noyau plusieurs fois plus petit, généralement assez proche du macronoyau. C'est un micronoyau. La différence entre ces deux noyaux n'est pas seulement de taille, elle est plus significative, affectant profondément leur structure.
Le macronoyau, par rapport au micronoyau, est beaucoup plus riche en une substance nucléaire spéciale (chromatine ou, plus précisément, acide désoxyribonucléique, en abrégé ADN), qui fait partie des chromosomes.
Des études récentes ont montré que le macronoyau a plusieurs dizaines (et dans certains ciliés et des centaines) fois un grand nombre chromosomes que les micronoyaux. Le macronoyau est un type très particulier de noyaux multichromosomiques (polyploïdes). Ainsi, la différence entre les micro et macronoyaux affecte leur composition chromosomique, qui détermine la richesse plus ou moins grande de leur substance nucléaire - la chromatine.
Dans l'un des types de ciliés les plus courants - des chaussures(Paramecium caudatum) - possède un macronoyau (en abrégé Ma) et un micronoyau (en abrégé Mi). Cette structure de l'appareil nucléaire est caractéristique de nombreux ciliés. D'autres peuvent avoir plusieurs Ma et Mi. Mais un trait caractéristique de tous les ciliés est la différenciation des noyaux en deux groupes qualitativement différents, en Ma et Mi, ou, en d'autres termes, le phénomène de dualisme nucléaire.
Comment se reproduisent les ciliés ? Prenons à nouveau comme exemple la chaussure à infusoires. Si vous plantez une seule copie de la chaussure dans un petit récipient (microaquarium), alors en une journée, il y aura deux, et souvent quatre ciliés. Comment cela peut-il arriver? Après une certaine période de nage active et d'alimentation, le cilié est quelque peu étiré en longueur. Puis, exactement au milieu du corps, apparaît une constriction transversale de plus en plus profonde (Fig. 90). Au final, les ciliés, pour ainsi dire, sont entrelacés en deux et deux individus sont obtenus à partir d'un individu, initialement un peu plus petit que l'individu parent. L'ensemble du processus de fission prend environ une heure à température ambiante. L'étude des processus internes montre qu'avant même l'apparition de la constriction transversale, le processus de fission de l'appareil nucléaire commence. Mi est partagé en premier, et seulement après Ma. Nous ne nous arrêterons pas ici considération détaillée processus de fission nucléaire et nous notons seulement que Mi est divisé par mitose, tandis que la division de Ma ressemble en apparence à la fission nucléaire directe - amitose. Ce processus de reproduction asexuée des ciliés de la chaussure, comme nous le voyons, est similaire à la reproduction asexuée des amibes et des flagellés. En revanche, les ciliés dans le processus de reproduction asexuée se divisent toujours en travers, tandis que chez les flagellés, le plan de division est parallèle à l'axe longitudinal du corps.
Lors de la division, une profonde restructuration interne du corps cilié se produit. Deux nouveaux péristomes, deux pharynx et deux ouvertures buccales sont formés. Dans le même temps, la division des noyaux basaux des cils est chronométrée, grâce à laquelle de nouveaux cils sont formés. Si le nombre de cils n'augmentait pas pendant la reproduction, à la suite de chaque division, les individus filles recevraient environ la moitié du nombre de cils de l'individu mère, ce qui conduirait à une «calvitie» complète des ciliés. En fait, cela ne se produit pas.

De temps en temps, la plupart des ciliés, y compris les chaussures, ont une forme spéciale et extrêmement particulière du processus sexuel, appelée conjugaison. Nous n'analyserons pas ici en détail toutes les mutations nucléaires complexes qui accompagnent ce processus, mais relevons seulement les plus importantes. La conjugaison se déroule comme suit (Fig. 91), deux ciliés s'approchent, sont étroitement appliqués l'un à l'autre par les faces ventrales, et sous cette forme ils nagent assez Longtemps ensemble (à la chaussure pendant environ 12 heures à température ambiante). Les conjugants se séparent alors. Que se passe-t-il dans le corps des ciliés lors de la conjugaison ? L'essence de ces processus est la suivante (Fig. 91). Le gros noyau (macronoyau) s'effondre et se dissout progressivement dans le cytoplasme. Les micronoyaux se divisent d'abord, certains des noyaux formés à la suite de la fission sont détruits (voir Fig. 91). Chacun des conjugants conserve deux noyaux. L'un de ces noyaux reste en place dans l'individu dans lequel il s'est formé (le noyau stationnaire), tandis que l'autre se déplace activement dans le partenaire de conjugaison (le noyau migrant) et fusionne avec son noyau stationnaire. Ainsi, dans chacun des conjugants à ce stade, un noyau est formé à la suite de la fusion des noyaux stationnaire et migrant. Ce noyau complexe est appelé syncaryon. La formation d'un syncaryon n'est rien de plus qu'un processus de fécondation. Et chez les organismes multicellulaires, le moment essentiel de la fécondation est la fusion des noyaux des cellules germinales. Chez les ciliés, les cellules germinales ne se forment pas, il n'y a que des noyaux sexuels, qui fusionnent les uns avec les autres. Ainsi, une fertilisation croisée mutuelle se produit.
Peu de temps après la formation du syncaryon, les conjugants se séparent. Selon la structure de leur appareil nucléaire, ils diffèrent encore à ce stade de manière très significative des ciliés dits neutres (non conjugués) habituels, puisqu'ils ne possèdent qu'un seul noyau chacun. À l'avenir, grâce au synkarion, l'appareil nucléaire normal est restauré. Le syncaryon est divisé (une ou plusieurs fois). Une partie des produits de cette division, par des transformations complexes associées à une augmentation du nombre de chromosomes et à un enrichissement en chromatine, se transforme en macronoyaux. D'autres conservent la structure caractéristique des micronoyaux. De cette manière, l'appareil nucléaire caractéristique et typique des ciliés est restauré, après quoi les ciliés commencent la reproduction asexuée par fission.
Ainsi, le processus de conjugaison comprend deux moments biologiques essentiels : la fécondation et la restauration d'un nouveau macronoyau dû au synkaryon.
Quelle est la signification biologique de la conjugaison, quel rôle joue-t-elle dans la vie des ciliés ? On ne peut pas l'appeler reproduction, car il n'y a pas d'augmentation du nombre d'individus. Les questions ci-dessus ont servi de matière à de nombreuses études expérimentales menées dans de nombreux pays. Le principal résultat de ces études est le suivant. Premièrement, la conjugaison, comme tout autre processus sexuel, dans lequel deux principes héréditaires (paternel et maternel) s'unissent dans un seul organisme, conduit à une augmentation de la variabilité héréditaire, de la diversité héréditaire. Une augmentation de la variabilité héréditaire augmente les capacités d'adaptation de l'organisme aux conditions environnementales. Le deuxième aspect biologiquement important de la conjugaison est le développement d'un nouveau macronoyau dû aux produits de fission du syncaryon et, en même temps, la destruction de l'ancien. Les données expérimentales montrent que c'est le macronoyau qui joue un rôle extrêmement important dans la vie des ciliés. Il contrôle tous les principaux processus vitaux et détermine le plus important d'entre eux - la formation (synthèse) d'une protéine qui constitue la partie principale du protoplasme d'une cellule vivante. Avec une reproduction asexuée prolongée par division, un processus particulier de «vieillissement» du macronoyau, et en même temps de la cellule entière, se produit: l'activité du processus métabolique diminue, le taux de division diminue. Après la conjugaison (au cours de laquelle, comme nous l'avons vu, l'ancien macronoyau est détruit), le taux métabolique et le taux de division sont restaurés. Étant donné que le processus de fécondation se produit pendant la conjugaison, qui dans la plupart des autres organismes est associée à la reproduction et à l'apparition d'une nouvelle génération, chez les ciliés, l'individu formé après la conjugaison peut également être considéré comme une nouvelle génération sexuée, qui surgit ici, car elle étaient, en raison du « rajeunissement » de l'ancien.
Sur l'exemple des chaussures ciliées, nous avons rencontré un représentant typique d'une classe étendue de ciliés. Cependant, cette classe se caractérise par une extraordinaire diversité d'espèces tant dans la structure que dans le mode de vie. Examinons de plus près certaines des formes les plus caractéristiques et les plus intéressantes.
Chez les ciliés, les cils de la chaussure couvrent uniformément toute la surface du corps. C'est un trait caractéristique de la structure (Holotricha). De nombreux ciliés se caractérisent par une nature différente du développement de la couverture ciliaire. Le fait est que les cils des ciliés sont capables, lorsqu'ils sont combinés, de former des complexes plus complexes. Par exemple, on observe souvent que des cils situés sur une ou deux rangées proches les unes des autres se rejoignent (se collent) ensemble, formant une plaque qui, comme les cils, est capable de battre. Ces formations contractiles lamellaires sont appelées membranella (si elles sont courtes) ou membranes (si elles sont longues). Dans d'autres cas, les cils sont réunis, situés dans un faisceau serré. Ces formations - cirres - ressemblent à un pinceau dont les poils individuels sont collés ensemble. Diverses formations ciliaires complexes sont caractéristiques de nombreux ciliés. Très souvent, la couverture ciliaire ne se développe pas uniformément, mais seulement dans certaines parties du corps.
INFUSORIA trompettiste (STENTOR POLYMORPHIQUE)
En eau douce, des espèces de grands beaux ciliés appartenant à sorte de trompettistes(Stentor). Ce nom est tout à fait cohérent avec la forme du corps de ces animaux, qui ressemble vraiment à un tuyau (Fig. 92), largement ouvert à une extrémité. Lors de la première rencontre avec des trompettistes en direct, on peut remarquer une caractéristique qui n'est pas caractéristique d'une chaussure. A la moindre irritation, y compris mécanique (par exemple, tapoter avec un crayon sur du verre, où il y a une goutte d'eau avec des trompettistes), leur corps se contracte brusquement et très rapidement (en une fraction de seconde), prenant une allure presque régulière forme sphérique. Puis, assez lentement (le temps est mesuré en secondes), le trompettiste se redresse, prenant sa forme caractéristique. Cette capacité du trompettiste à se contracter rapidement est due à la présence de fibres musculaires particulières situées le long du corps et dans l'ectoplasme. Ainsi, un système musculaire peut également se développer dans un organisme unicellulaire.

Il existe des espèces dans le genre des trompettistes, dont certaines se caractérisent par une couleur plutôt vive. Très commun en eau douce trompettiste bleu(Stentor coeruleus), qui est bleu vif. Cette coloration du trompettiste est due au fait que les plus petits grains de pigment bleu se situent dans son ectoplasme.
Une autre espèce de trompettiste (Stentor polymorphus) est souvent de couleur verte. La raison de cette coloration est tout autre. Couleur verte du fait que de petites algues vertes unicellulaires vivent et se multiplient dans l'endoplasme du cilié, ce qui donne au corps du trompettiste une couleur caractéristique. Stentor polymorphus est un exemple typique de cohabitation mutuellement bénéfique - symbiose. Le trompettiste et les algues sont dans une relation symbiotique mutuelle : le trompettiste protège les algues vivant dans son corps et leur fournit du dioxyde de carbone formé à la suite de la respiration ; de leur côté, les algues fournissent au trompettiste de l'oxygène, libéré lors du processus de photosynthèse. Apparemment, une partie des algues est digérée par les ciliés, nourriture du trompettiste.
Les trompettistes nagent lentement dans l'eau avec l'extrémité large vers l'avant. Mais ils peuvent aussi être temporairement attachés au substrat par l'extrémité étroite postérieure du corps, sur laquelle se forme une petite ventouse.
Dans le corps du trompettiste, on peut distinguer la région du tronc s'étendant de l'arrière vers l'avant et un large champ périoral (péristomial) situé presque perpendiculairement à celui-ci. Ce champ ressemble à un entonnoir plat asymétrique, sur un bord duquel se trouve un évidement - un pharynx menant à l'endoplasme du cilié. Le corps du trompettiste est couvert de rangées longitudinales de cils courts. Le long du bord du champ péristomial, une zone quasi-orale (adorale) fortement développée de membranella est située dans un cercle (Fig. 92). Cette zone est constituée d'un grand nombre de plaques ciliées individuelles, chacune d'entre elles étant composée à son tour de nombreux cils collés les uns aux autres, situés sur deux rangées rapprochées.
Dans la région de l'ouverture buccale, les membranes péribuccales sont enroulées vers le pharynx, formant une spirale gauche. Le flux d'eau, provoqué par l'oscillation de la membranella périorale, est dirigé vers l'ouverture buccale (dans la profondeur de l'entonnoir formé par l'extrémité antérieure du corps). Avec l'eau, les particules alimentaires en suspension dans l'eau pénètrent également dans le pharynx. Les objets alimentaires du trompettiste sont plus diversifiés que ceux du chausson. En plus des bactéries, il mange de petits protozoaires (par exemple, des flagellés), des algues unicellulaires, etc.
Le trompettiste a une vacuole contractile bien développée, qui a une structure particulière. Le réservoir central est situé dans le tiers antérieur du corps, légèrement en dessous de l'ouverture buccale. Deux longs canaux d'adduction en partent. L'un d'eux va du réservoir à l'extrémité postérieure du corps, le second est situé dans la région du champ péristomial parallèle à la zone péribuccale de la membranella.
L'infusoire trompettiste est un objet de prédilection pour les recherches expérimentales sur la régénération. De nombreuses expériences ont prouvé la grande capacité de régénération des trompettistes. Un cilié avec un scalpel fin peut être coupé en plusieurs parties, et chacune d'elles à travers un temps limité(plusieurs heures, parfois un jour ou plus) se transformera en un petit trompettiste proportionnellement construit, qui, à la suite d'une alimentation énergique, atteindra la taille typique de cette espèce. Pour compléter les processus de récupération, la pièce de régénération doit contenir au moins un segment d'un macronoyau perlé.
Le trompettiste, comme nous l'avons vu, a des cils différents : d'une part, ils sont courts, couvrant tout le corps, et d'autre part, il y a une zone quasi-orale de membranella. Conformément à ce trait caractéristique de la structure, le détachement de ciliés, auquel appartient le trompettiste, a été nommé ciliés ciliés(Hétérotriche).
INFUSORIA BURSARIA (BURSARIA TRUNCATELLA)
Le deuxième représentant intéressant des ciliés ciliés se trouve souvent dans les eaux douces bourse(Bursaria truncatella, fig. 93). C'est un géant parmi les ciliés : ses dimensions peuvent atteindre 2 mm, les tailles les plus courantes sont de 0,5 à 1,0 mm. Bursaria est clairement visible à l'œil nu. Conformément à son nom, la bourse a la forme d'un sac, largement ouvert à l'extrémité antérieure (bourse est un mot latin, traduit en russe signifie «bourse», «sac») et quelque peu élargi à l'extrémité postérieure. Le corps entier du cilié est couvert de rangées longitudinales de cils courts. Leur battement provoque un mouvement vers l'avant plutôt lent de l'animal. La bourse nage comme si elle "roulait" d'un côté à l'autre.

De l'extrémité avant dans les profondeurs du corps (environ 2/3 de sa longueur) une dépression périorale fait saillie - un péristome. Du côté ventral, il communique avec le milieu extérieur par une fente étroite ; du côté dorsal, la cavité du péristome ne communique pas avec le milieu extérieur. Si vous regardez la coupe transversale du tiers supérieur du corps de la bourse (Fig. 93, B), vous pouvez voir que la cavité du péristome occupe la majeure partie du corps, tandis que le cytoplasme l'entoure sous la forme d'un rebord.
A l'extrémité antérieure du corps, à gauche, une zone très puissamment développée de membranella périorale (adorale) (Fig. 93, 4) prend naissance dans la bourse. Il descend dans la profondeur de la cavité du péristome en tournant vers la gauche. La zone adorale s'étend jusqu'à la partie la plus profonde du péristome. Il n'y a pas d'autres formations ciliaires dans la cavité péristomiale en dehors des membranes péribuccales, à l'exception de la bande ciliaire longeant la face ventrale de la cavité péristomiale (Fig. 93, 10). À l'intérieur mur arrière la cavité péristomiale a une fente étroite sur presque toute sa longueur (Fig. 93, 7), dont les bords sont généralement étroitement contiguës. C'est l'ouverture de la bouche. Ses bords ne s'écartent qu'au moment de manger.
Les Bursaria n'ont pas une spécialisation alimentaire étroite, mais ce sont principalement des prédateurs. En avançant, ils rencontrent divers petits animaux. Grâce au travail des membranelles de la zone quasi-orale, la proie est entraînée avec force dans la vaste cavité péristomiale, d'où elle ne peut plus nager. Les objets alimentaires sont pressés contre la paroi dorsale de la cavité péristomiale et pénètrent dans l'endoplasme à travers la fissure buccale en expansion. Les Bursaria sont très voraces, elles peuvent avaler des objets assez volumineux : par exemple, leur nourriture préférée est les ciliés de chaussures. Bursaria est capable d'avaler 6 à 7 chaussures d'affilée. En conséquence, de très grandes vacuoles digestives se forment dans l'endoplasme de la bourse.
L'appareil nucléaire de la bourse est assez compliqué. Ils ont un long macronoyau en forme de saucisse et un grand nombre (jusqu'à environ 30) de petits micronoyaux dispersés au hasard dans l'endoplasme du cilié.
Les Bursaria font partie des rares espèces de ciliés d'eau douce qui n'ont pas de vacuole contractile. La façon dont l'osmorégulation est effectuée dans ce grand cilié n'est pas encore tout à fait claire. Sous l'ectoplasme de la bourse dans différentes parties du corps, on peut observer différentes formes et tailles de bulles de liquide - vacuoles, qui changent de volume. Apparemment ces forme irrégulière vacuoles et correspondent dans leur fonction aux vacuoles contractiles des autres ciliés.

Il est intéressant d'observer les étapes successives de la reproduction asexuée de Bursaria. Aux stades initiaux de la division, il y a une réduction complète de toute la cavité du péristome et de la zone quasi-orale de la membranella (Fig. 94). Seul le cil externe est conservé. L'infusoire prend la forme d'un œuf. Après cela, le corps est lacé d'une rainure transversale en deux moitiés. Dans chacun des ciliés filles résultants, à travers des transformations assez complexes, un péristome typique et une zone péribuccale de membranella se développent. L'ensemble du processus de répartition de la bourse se déroule rapidement et prend un peu plus d'une heure.
Il est très facile d'observer un autre processus vital important dans la bourse, dont l'apparition est associée à des conditions défavorables aux ciliés, le processus de formation de kystes (enkystation). Ce phénomène est caractéristique, par exemple, de l'amibe. Mais il s'avère que même des protozoaires aussi complexes que les ciliés sont capables de passer à un état inactif. Si la culture où vit la bourse n'est pas alimentée ou refroidie à temps, l'enkystement de masse commencera dans quelques heures. Ce processus se déroule comme suit. Les bursarides, ainsi qu'avant la division, perdent le péristome et la zone péribuccale des membranells. Ensuite, ils deviennent complètement sphériques, après quoi ils distinguent une double coque de forme caractéristique (Fig. 94, D).
Dans l'état de kystes, la bourse peut durer des mois. Lorsque des conditions favorables se présentent, la coquille du kyste éclate, la bourse en sort, développe un péristome et passe à une vie active.
STYLONICHIA MYTILUS
Un appareil ciliaire très complexe et diversement différencié a des ciliés liés à décollement gastro-intestinal(Hypotricha), dont de nombreuses espèces vivent aussi bien en eau douce qu'en eau de mer. L'un des représentants les plus courants et les plus fréquemment rencontrés de ce groupe intéressant peut être appelé stilonychie(Stylonichia mytilus). C'est un cilié assez grand (longueur jusqu'à 0,3 mm), vivant au fond des réservoirs d'eau douce, sur végétation aquatique(Fig. 95). Contrairement au chausson, au trompettiste et à la bourse, la stylonychie n'a pas de couverture ciliaire continue et l'ensemble de l'appareil ciliaire est représenté par un nombre limité de formations ciliaires strictement définies.

Le corps de Stilonychia (comme la plupart des autres ciliés ventraux) est fortement aplati dans la direction dorso-abdominale, et ses côtés dorsal et ventral, ses extrémités antérieure et postérieure se distinguent clairement. Le corps est un peu élargi vers l'avant, rétréci vers l'arrière. Lors de l'examen de l'animal du côté ventral, on voit clairement que dans le tiers antérieur à gauche, il y a une ouverture pennée et orale complexe.
Sur la face dorsale, les cils sont assez rarement situés, qui ne sont pas capables de battre. On peut plutôt les appeler poils élastiques fins. Ils sont immobiles et n'ont rien à voir avec la fonction du mouvement. Ces cils se voient généralement attribuer une fonction tactile et sensible.
Toutes les formations ciliaires associées au mouvement et à la capture de nourriture sont concentrées sur la face ventrale de l'animal (Fig. 95). Il existe un petit nombre de formations épaisses en forme de doigt situées dans plusieurs groupes. Ce sont des cirrhi abdominaux. Chacun d'eux est une formation ciliaire complexe, résultat d'une connexion étroite (collage) de plusieurs dizaines de cils individuels. Ainsi, les cirres sont comme des pinceaux dont les poils individuels sont étroitement rapprochés et reliés les uns aux autres.
Avec l'aide des cirrhos, l'animal se déplace assez rapidement, «court» le long du substrat. En plus de "ramper" et de "courir" sur le substrat, la stilonychie est capable de produire des sauts assez nets et forts, se détachant immédiatement du substrat. Ces mouvements brusques sont effectués à l'aide de deux puissants cirres de queue (Fig. 95), qui ne participent pas au "rampement" habituel.
Deux rangées de cirrhées marginales (marginales) sont situées le long du bord du corps à droite et à gauche. Du bord droit de l'animal, ils parcourent tout le corps, tandis que du bord gauche, ils n'atteignent que la région du péristome. Ces formations ciliaires servent à propulser l'animal lorsqu'il est détaché du substrat et nage librement dans l'eau.
On voit donc que l'appareil ciliaire varié et spécialisé de la stylonychie lui permet d'effectuer des mouvements très divers, contrairement par exemple à de simples glissades dans l'eau, comme une chaussure ou un trompettiste.
L'appareil ciliaire associé à la fonction de nutrition est également complexe. Nous avons déjà vu que la cavité quasi-orale (péristome), au fond de laquelle se trouve l'ouverture buccale menant au pharynx, est située dans la moitié antérieure de l'animal à gauche. Le long de la marge gauche, à partir de l'extrémité très antérieure du corps, il y a une zone fortement développée de membranolles périorales (adorales). Avec leurs coups, ils dirigent le flux d'eau vers l'ouverture de la bouche. De plus, dans la région de la cavité péristomiale, il existe trois autres membranes contractiles (membranes), qui s'étendent dans le pharynx avec leurs extrémités internes, et un certain nombre de cils périoraux spéciaux (Fig. 95). Tout cet appareil complexe sert à capturer et à diriger les aliments dans l'ouverture de la bouche.
Stilonychia est l'un des protozoaires avec une très large gamme d'objets alimentaires. On peut à juste titre l'appeler omnivore. Elle peut manger, comme une chaussure, des bactéries. Parmi ses objets alimentaires figurent les flagellés, des algues unicellulaires (souvent des diatomées). Enfin, Stilonychia peut également être un prédateur, attaquant d'autres espèces de ciliés plus petites et les dévorant.
Stilonychia a une vacuole contractile. Il est constitué d'un réservoir central situé à l'extrémité postérieure gauche du péristome et d'un canal adducteur dirigé vers l'arrière.
L'appareil nucléaire, comme toujours chez les ciliés, est constitué d'un macronoyau et d'un micronoyau.
Le macronoyau est composé de deux moitiés reliées par une mince constriction ; il y a deux micronoyaux, ils sont situés directement près des deux moitiés de Ma.
Stilonychia, en partie bursaria, trompettiste - ce sont tous des ciliés avec une large gamme d'objets alimentaires. La capacité d'absorber divers aliments est caractéristique de la plupart des ciliés. Cependant, parmi eux, on peut également trouver de telles espèces qui sont strictement spécialisées par rapport à la nature de la nourriture.
PREDATEURS INFUSORIENS
Parmi les ciliés, il y a des prédateurs très "pointilleux" sur leurs proies. Un bon exemple est l'infusoire. didynie(Didinium nasutum). Didinium est un cilié relativement petit, mesurant en moyenne environ 0,1 à 0,15 mm de long. L'extrémité avant est allongée en forme de trompe, à l'extrémité de laquelle est placée l'ouverture buccale. L'appareil ciliaire est représenté par deux corolles de cils (Fig. 96). Didinius nage rapidement dans l'eau, changeant souvent de direction. La nourriture préférée du didinia est la chaussure ciliée. Dans ce cas, le prédateur est plus petit que sa proie. Didinius pénètre dans la proie avec un tronc, puis, élargissant progressivement la bouche s'ouvrant de plus en plus, avale la chaussure entière! Dans la trompe, il y a un appareil spécial, appelé tige. Il se compose d'un certain nombre de bâtonnets élastiques solides situés dans le cytoplasme le long de la périphérie de la trompe. On pense que cet appareil augmente la résistance des parois de la trompe, qui ne se brise pas en avalant une proie aussi énorme par rapport au didinium, comme une chaussure. Didinius - exemple dernier recours prédation parmi les protozoaires. Si nous comparons le didinium avalant sa proie - les chaussures - avec la prédation chez les animaux supérieurs, il est difficile de trouver des exemples similaires.

Didinius, avalant des paramécies, bien sûr, gonfle beaucoup. Le processus de digestion est très rapide, à température ambiante, il ne prend que deux heures environ. Ensuite, les restes non digérés sont jetés et le didinium commence à chasser une autre victime. Des études spéciales ont montré que le "régime" quotidien de didinia est de 12 chaussures - un appétit vraiment colossal ! Il faut garder à l'esprit que dans les intervalles entre les "chasses" suivantes, la didinia se divise parfois. En manque de nourriture, les didinia s'enkystent très facilement et sortent tout aussi facilement des kystes.
ciliés herbivores
Beaucoup moins fréquent que la prédation, on le retrouve chez les ciliés « pur végétarisme » - se nourrissant exclusivement aliment végétal. L'un des rares exemples de ciliés « végétariens » peut être représentatif genre passula(Nassule). Leur objet alimentaire est une algue bleu-vert filamenteuse (Fig. 97).

Ils pénètrent dans l'endoplasme par la bouche, située sur le côté, puis sont tordus par les infusoires en une spirale serrée, qui est progressivement digérée. Les pigments d'algues pénètrent partiellement dans le cytoplasme des ciliés et le colorent d'une couleur vert foncé brillant.
SUVOIKA (VORTICELLA NEBULIFERA)
Un groupe intéressant et assez important de ciliés en termes de nombre d'espèces sont les formes sessiles attachées au substrat, formant détachement des yeux ronds(Peritricha). Les membres les plus courants de ce groupe sont suvoyki(espèce du genre Vorticella).
Souvoyki ressemblent à une fleur élégante comme une cloche ou un muguet, assis sur une longue tige, qui est attachée au substrat avec son extrémité. La suvoyka passe la majeure partie de sa vie dans un état attaché au substrat.

Considérez la structure du corps des ciliés. Chez différentes espèces, leurs tailles varient dans une gamme assez large (jusqu'à environ 150 microns). Le disque oral (Fig. 98) est situé sur une partie antérieure élargie du corps, qui est complètement dépourvue de cils. L'appareil ciliaire est situé uniquement le long du bord du disque oral (péristomal) (Fig. 98) dans une rainure spéciale, à l'extérieur de laquelle un rouleau (lèvre péristomale) est formé. Trois membranes ciliées courent le long du bord du rouleau, dont deux sont situées verticalement, une (extérieure) est horizontale. Ils forment un peu plus d'un tour complet de l'hélice. Ces membranes sont en mouvement de scintillement constant, dirigeant le flux d'eau vers l'ouverture de la bouche. L'appareil buccal commence assez profondément par un entonnoir au bord du champ péristomial (Fig. 98), dans la profondeur duquel se trouve une ouverture buccale menant à un court pharynx. Les Suvoys, comme les chaussures, se nourrissent de bactéries. Leur ouverture buccale est constamment ouverte et il y a un flux continu d'eau vers la bouche.
Une vacuole contractile sans canaux adducteurs est située près de l'ouverture de la bouche. Le macronoyau a la forme d'un ruban ou d'une saucisse, à côté duquel se trouve un seul petit micronoyau.
Suvoyka est capable de raccourcir fortement la tige qui, en une fraction de seconde, est tordue avec un tire-bouchon. Dans le même temps, le corps du cilié se contracte également : le disque et les membranes péristomiales sont tirés vers l'intérieur et toute l'extrémité antérieure se ferme.

La question se pose naturellement : les suvoys étant fixés au substrat, de quelle manière s'effectue leur installation dans le réservoir ? Cela se produit par la formation d'une étape flottante, le vagabond. À l'extrémité postérieure du corps cilié, une corolle de cils apparaît (Fig. 99). En même temps, le disque péristomial se rétracte vers l'intérieur et le cilié se sépare de la tige. Le clochard qui en résulte est capable de nager pendant plusieurs heures. Les événements se déroulent alors dans ordre inverse: le cilié s'attache au substrat par son extrémité postérieure, la tige se développe, la corolle postérieure des cils se réduit, le disque péristomial se redresse à l'extrémité antérieure et les membranes adorales commencent à fonctionner. La formation de vagabonds dans la suvoyka est souvent associée au processus de reproduction asexuée. L'infusoire sur la tige se divise et l'un des individus filles (et parfois les deux) devient un clochard et nage.
De nombreux types de suwoks sont capables de s'enkyster dans des conditions défavorables.
Parmi les ciliés sessiles appartenant au groupe des ciliés, seules relativement peu d'espèces, comme les suvoes discutés ci-dessus, sont des formes vivantes solitaires. La plupart des espèces incluses ici sont des organismes coloniaux.
Habituellement, la colonialité résulte d'une reproduction asexuée ou végétative incomplète. Les individus formés à la suite de la reproduction, dans une plus ou moins grande mesure, conservent des liens les uns avec les autres et forment ensemble une individualité organique d'un ordre supérieur, unissant grandes quantités individus individuels, qui prend le nom de la colonie (nous avons déjà rencontré des exemples d'organismes coloniaux dans classe des flagellés.

Des colonies de ciliés ronds se forment du fait que les individus séparés ne se transforment pas en vagabonds, mais restent en contact les uns avec les autres à l'aide de tiges (Fig. 100). Dans ce cas, la tige principale de la colonie, ainsi que ses premières branches, ne peut être attribuée à aucun des individus, mais appartient à l'ensemble de la colonie dans son ensemble. Parfois, la colonie n'est constituée que d'un petit nombre d'individus, alors que chez d'autres espèces de ciliés, le nombre d'individus individuels de la colonie peut atteindre plusieurs centaines. Cependant, la croissance de toute colonie n'est pas illimitée. En atteignant les dimensions caractéristiques de cette espèce, la colonie cesse d'augmenter et les individus formés à la suite de la division développent une corolle de cils, deviennent vagabonds et s'éloignent, donnant naissance à de nouvelles colonies.
Les colonies de ciliés ronds sont de deux types. Chez certains, la tige de la colonie est irréductible : lorsqu'ils sont irrités, seuls les individus individuels de la colonie se contractent, dessinant dans le penné, mais l'ensemble de la colonie dans son ensemble ne subit pas de modifications (ce type de colonie comprend, par exemple, les genres Epistylis, Opercularia). Chez d'autres (par exemple, le genre Carchesium), la tige de toute la colonie est capable de se contracter, puisque le cytoplasme traverse toutes les branches et relie ainsi tous les individus de la colonie entre eux. Lorsque ces colonies sont irritées, elles rétrécissent entièrement. Toute la colonie réagit alors comme un tout, comme une individualité organique.
Parmi tous les ciliés coloniaux à ciliés ronds, un intérêt particulier est peut-être zootamnique(Zoothamnium arbuscula). Les colonies de ce cilié se distinguent par une régularité particulière de la structure. De plus, un phénomène biologique intéressant de polymorphisme est mis en évidence ici au sein de la colonie.

La colonie de zootamnies ressemble à un parapluie. Sur une tige principale de la colonie se trouvent des branches secondaires (Fig. 101). La taille d'une colonie adulte est de 2 à 3 mm, elle est donc clairement visible à l'œil nu. Zootamnii vivent dans petits étangs avec eau propre. Leurs colonies se trouvent généralement sur des plantes sous-marines, le plus souvent sur des élodées (peste de l'eau).
Les tiges d'une colonie de zootamnies sont contractiles, puisque le cytoplasme contractile traverse toutes les branches de la colonie, à l'exception de la partie basale de la tige principale. Avec une réduction qui se produit très rapidement et brusquement, toute la colonie se rassemble en une masse.

La zootamnie se caractérise par une disposition strictement régulière des branches. Une tige principale est attachée au substrat. Neuf branches principales de la colonie partent de sa partie supérieure dans un plan perpendiculaire à la tige, strictement régulièrement situées les unes par rapport aux autres (Fig. 102, 6). Des branches secondaires s'étendent à partir de ces branches, sur lesquelles sont assis les individus individuels de la colonie. Chaque branche secondaire peut avoir jusqu'à 50 ciliés. Le nombre total d'individus dans la colonie atteint 2 à 3 000 individus.
La plupart des individus de la colonie dans leur structure ressemblent à de petits suvoiks simples, d'une taille de 40 à 60 microns. Mais en plus des petits individus, appelés microzoïdes, sur les colonies adultes, approximativement au milieu des branches principales, se développent des individus d'un type et d'une taille complètement différents (Fig. 102, 5). Ce sont de grands individus sphériques d'un diamètre de 200 à 250 microns, dépassant le volume du microzoïde d'une centaine de fois ou plus en masse. Les grands individus sont appelés macrozoïdes.
Dans leur structure, ils diffèrent considérablement des petits individus de la colonie. Leur péristome n'est pas exprimé : il est rentré et ne fonctionne pas. Dès le début de son développement à partir d'un microzoïde, un macrozoaire cesse de se nourrir seul. Il est dépourvu de vacuoles digestives. La croissance du macrozoïde se fait apparemment aux dépens de substances pénétrant par les ponts cytoplasmiques reliant entre eux tous les individus de la colonie. Dans la partie du corps du macrozoïde, avec laquelle il est attaché à la tige, il y a une accumulation de grains spéciaux (granules) qui, comme nous le verrons, jouent un rôle important dans son destin ultérieur. Que sont ces gros macrozoïdes sphériques, quel est leur rôle biologique dans la vie d'une colonie de zootamnies ? L'observation montre que les macrozoïdes sont de futurs vagabonds à partir desquels de nouvelles colonies se développent. Atteindre limite de taille, le macrozoïde développe une corolle de cils, se sépare de la colonie et nage. En même temps, sa forme change quelque peu, de sphérique elle devient conique. Après un certain temps, le tramp est toujours attaché au substrat avec le côté sur lequel se trouve la granularité. La formation et la croissance de la tige commencent immédiatement et des granules sont dépensées pour la construction de la tige, qui sont localisées à l'extrémité postérieure du vagabond. Au fur et à mesure que la tige grandit, le grain disparaît. Une fois que la tige a atteint la longueur finale caractéristique de la zootamnie, une série de divisions rapidement successives commence, conduisant à la formation d'une colonie. Ces divisions sont faites dans un ordre strictement défini (Fig. 102).
Nous ne nous attarderons pas sur les détails de ce processus. Faisons seulement attention au phénomène intéressant suivant. Lors des premières divisions des clochards zootamnia, lors du développement d'une colonie chez les individus en formation, le penné et la bouche ne fonctionnent pas. L'alimentation commence plus tard, lorsque la jeune colonie compte déjà 12 à 16 individus. Ainsi, toutes les premières étapes du développement d'une colonie se déroulent exclusivement aux dépens des réserves qui se sont formées dans le corps du macrozoïde lors de sa croissance et de son développement sur la colonie mère. Il existe une similitude indéniable entre le développement de la zootamnie vagrante et le développement de l'œuf chez les animaux multicellulaires. Cette similitude s'exprime dans le fait que le développement dans les deux cas s'effectue au détriment des réserves précédemment accumulées, sans la perception de nourriture de l'environnement extérieur.
Lors de l'étude des ciliés ronds sessiles, la question se pose: comment se déroule la forme du processus sexuel caractéristique des ciliés - conjugaison? Il s'avère que dans le cadre d'un mode de vie sédentaire, il subit des changements importants. Au début du processus sexuel, de très petits vagabonds spéciaux se forment sur la colonie. Se déplaçant activement à l'aide d'une corolle de cils, ils rampent pendant un certain temps le long de la colonie, puis se conjuguent avec de grands individus sédentaires normaux de la colonie. Ainsi, la différenciation des conjugants en deux groupes d'individus se produit ici : petits, mobiles (microconjugants) et plus grands, immobiles (macroconjugants). Cette différenciation des conjugants en deux catégories, dont l'une (les microconjugants) est mobile, était une nécessaire adaptation à la sédentarité. Sans cela, le déroulement normal du processus sexuel (conjugaison) ne pourrait évidemment pas être assuré.
SUCEUR CILITAIRE (SUCTORIA)
Un groupe très particulier en termes de mode d'alimentation est représenté par ciliés suceurs(Suctoria). Ces organismes, comme le suvoyka et d'autres ciliés ciliés, sont sessiles. Le nombre d'espèces appartenant à cet ordre se mesure par plusieurs dizaines. La forme du corps des ciliés suceurs est très diversifiée. Certaines de leurs espèces caractéristiques sont représentées à la figure 103. Certaines reposent sur le substrat sur des tiges plus ou moins longues, d'autres n'ont pas de tiges, chez certaines le corps se ramifie assez fortement, etc. Cependant, malgré la variété des formes, tous les ciliés suceurs se caractérisent par les deux caractéristiques suivantes :
1) absence complète (dans les formes adultes) de l'appareil ciliaire,
2) la présence d'appendices spéciaux - des tentacules qui servent à aspirer les proies.

Dans différents types de ciliés suceurs, le nombre de tentacules n'est pas le même. Souvent, ils sont réunis en groupes. Avec un fort grossissement du microscope, on peut voir qu'à l'extrémité du tentacule est équipé d'un petit épaississement en forme de massue.
Comment fonctionnent les tentacules ? Il n'est pas difficile de répondre à cette question en observant pendant un certain temps des ciliés suceurs. Si un petit protozoaire (flagellé, infusoire) touche le tentacule de la succion, il s'y collera instantanément. Toutes les tentatives de la victime pour s'échapper sont généralement vaines. Si vous continuez à observer la proie collée aux tentacules, vous pouvez voir qu'elle commence progressivement à diminuer de taille. Son contenu est progressivement "pompé" à travers les tentacules dans l'endoplasme du cilié suceur jusqu'à ce qu'il ne reste qu'une seule pellicule de la victime, qui est jetée. Ainsi, les tentacules des ciliés suceurs sont tout à fait uniques, nulle part ailleurs dans le monde animal se trouvent des organes permettant de piéger et en même temps de sucer de la nourriture (Fig. 103).
Les ciliés suceurs sont des prédateurs immobiles qui ne chassent pas les proies, mais les attrapent instantanément, si seulement une proie imprudente les touche elle-même.

Pourquoi référons-nous ces organismes particuliers à la classe des ciliés ? À première vue, ils n'ont rien à voir avec eux. Les faits suivants parlent de l'appartenance des suctoria aux ciliés. Premièrement, ils ont un appareil nucléaire typique des ciliés, composé d'un macronoyau et d'un micronoyau. Deuxièmement, lors de la reproduction, ils développent des cils qui sont absents chez les individus "adultes". La reproduction asexuée et, en même temps, la réinstallation des ciliés suceurs s'effectuent par la formation de vagabonds, équipés de plusieurs corolles annulaires de cils. La formation de vagabonds à suctoria peut se produire de différentes manières. Parfois, ils se forment à la suite d'une division pas tout à fait uniforme (bourgeonnement), dans laquelle chaque rein qui se sépare vers l'extérieur reçoit un segment de macronoyau et un micronoyau (Fig. 104, L). Sur un individu maternel, plusieurs bourgeons filles peuvent se former simultanément (Fig. 104, 5). Chez d'autres espèces (Fig. 104, D, E), on observe une méthode très particulière de "bourgeonnement interne". Dans le même temps, une cavité se forme à l'intérieur du corps de la mère suceuse, dans laquelle se forme le rein clochard. Il sort par des trous spéciaux, à travers lesquels il "se serre" avec une difficulté connue.
Ce développement de l'embryon à l'intérieur du corps de la mère, puis l'acte de procréer, est une analogie intéressante des plus simples avec ce qui se passe dans les organismes multicellulaires supérieurs.
Dans les pages précédentes, plusieurs représentants typiques libres de la classe des ciliés ont été considérés, différemment adaptés aux différentes conditions environnementales. Il est intéressant d'aborder la question de l'adaptation des ciliés aux conditions de vie et, d'autre part, de voir quelles sont les caractéristiques caractéristiques communes ciliés vivant dans certaines conditions environnementales bien définies.
A titre d'exemple, prenons deux habitats très différents : la vie dans la composition du plancton et la vie sur le fond dans l'épaisseur du sable.
INFUSORIE PLANCTONIQUE
Un assez grand nombre d'espèces ciliées se trouvent dans le plancton marin et d'eau douce.
Les caractéristiques d'adaptation à la vie dans la colonne d'eau sont particulièrement prononcées chez les radiolaires. La principale ligne d'adaptation à un mode de vie planctonique est réduite au développement de telles caractéristiques structurelles qui contribuent à la montée en flèche de l'organisme dans la colonne d'eau.

Une famille planctonique typique, d'ailleurs presque exclusivement marine, de ciliés est tintinnides(Tintinnidés, fig. 105, 5). Le nombre total d'espèces de tintinnidés connues à ce jour est d'environ 300. Ce sont de petites formes, caractérisées par le fait que le corps protoplasmique du cilié est placé dans une maison transparente, légère et en même temps solide, constituée de matière organique. Un disque dépasse de la maison, portant une corolle de cils, qui sont en mouvement de scintillement constant. À l'état d'infusoires planant dans la colonne d'eau, il est principalement soutenu par le travail actif constant de l'appareil ciliaire. La maison remplit évidemment la fonction de protection du bas du corps du cilié. Seules 2 espèces de tintinnidés vivent en eau douce (sans compter 7 espèces caractéristiques uniquement du lac Baïkal).
Les ciliés d'eau douce ont d'autres adaptations à la vie dans le plancton. Chez beaucoup d'entre eux, le cytoplasme est très fortement vacuolé (Loxodes, Condylostoma, Trachelius), de sorte qu'il ressemble à de la mousse. Cela conduit à une réduction significative de la gravité spécifique. Tous les ciliés répertoriés ont en outre une couverture ciliaire, grâce à laquelle le corps des ciliés, en termes de gravité spécifique, n'est que légèrement plus grand que gravité spécifique l'eau, facilement maintenue dans un état de "soaring". Chez certaines espèces, la forme du corps contribue à augmenter la surface spécifique et facilite l'envolée dans l'eau. Par exemple, certains ciliés planctoniques du lac Baïkal ressemblent à un parapluie ou à un parachute (Liliomorpha, Fig. 105, 2). Il existe un cilié suceur planctonique dans le lac Baïkal (Mucophrya pelagica, Fig. 105, 4), qui diffère nettement de ses parents sessiles. Cette espèce est dépourvue de tige. Son corps protoplasmique est entouré d'une large gaine visqueuse, une adaptation conduisant à une perte de poids. De longs tentacules minces dépassent, qui, avec leur fonction directe, en remplissent probablement une autre - une augmentation de la surface spécifique, ce qui contribue à la montée en flèche dans l'eau.
Enfin, il est nécessaire de mentionner une autre forme, pour ainsi dire indirecte, d'adaptation des ciliés à la vie dans le plancton. Il s'agit de l'attachement de petits ciliés à d'autres organismes qui mènent un mode de vie planctonique. Oui, parmi ciliés circonciliés(Peritricha) il existe des espèces assez nombreuses qui se fixent sur les copépodes planctoniques. C'est un mode de vie normal et normal pour ces types de ciliés.
Avec les ciliés circonciliaires et parmi succion(Suctoria) il y a des espèces qui s'installent sur des organismes planctoniques.
CILICIENS VIVANT DANS LE SABLE
Les plages de sable et les hauts-fonds représentent un habitat extrêmement particulier. Le long de la côte des mers, ils occupent de vastes espaces et se caractérisent par une faune particulière.
Réalisé pour dernières années De nombreuses études dans divers pays ont montré que la strate de nombreux sables marins est très riche en faune microscopique ou microscopique variée. Entre les particules de sable, il y a de nombreux petits et minuscules espaces remplis d'eau. Il s'avère que ces espaces sont richement peuplés d'organismes appartenant aux groupes les plus divers du monde animal. Des dizaines d'espèces de crustacés vivent ici, annélides, vers ronds, en particulier de nombreux vers plats, certains mollusques, coelentérés. En grand nombre, il y a aussi des protozoaires, principalement des ciliés. Selon les données modernes, la faune de ciliés habitant l'épaisseur des sables marins comprend environ 250 à 300 espèces. Si nous gardons à l'esprit non seulement les ciliés, mais également d'autres groupes d'organismes habitant l'épaisseur du sable, le nombre total de leurs espèces sera très important. L'ensemble des animaux habitant l'épaisseur du sable, vivant dans les moindres interstices entre les grains de sable, s'appelle la faune psammophile.
La richesse et la composition spécifique de la faune psammophile sont déterminées par de nombreux facteurs. Parmi eux, notamment importance a la taille de particules de sable. Les sables à gros grains ont une faune pauvre. La faune des sables limoneux à grains très fins (avec un diamètre de particule inférieur à 0,1 mm) est également pauvre, où, évidemment, les espaces entre les particules sont trop petits pour que les animaux y vivent. Les sables les plus riches en vie sont à grains moyens et fins.
Le deuxième facteur qui joue un rôle important dans le développement de la faune psammophile est la richesse du sable en restes organiques et en matière organique en décomposition (le degré dit de saprobité). Les sables dépourvus de matière organique sont pauvres en vie. D'autre part, les sables sont également presque sans vie et sont très riches en matière organique, car la décomposition de la matière organique entraîne un appauvrissement en oxygène. Souvent, la fermentation anaérobie du sulfure d'hydrogène est ajoutée à cela.
La présence de sulfure d'hydrogène libre est un facteur extrêmement négatif affectant le développement de la faune.
Une flore assez riche d'algues unicellulaires (diatomées, péridiniums) se développe parfois dans les couches superficielles de sable. C'est un facteur qui favorise le développement de la faune psammophile, puisque de nombreux petits animaux (dont les ciliés) se nourrissent d'algues.
Enfin, un facteur qui a un effet très négatif sur la faune psammophile est le surf. C'est tout à fait compréhensible, car les vagues, qui déferlent sur les couches supérieures de sable, tuent ici tous les êtres vivants. La faune psammophile est la plus riche dans les criques abritées et bien chauffées. Le flux et le reflux n'empêchent pas le développement de la faune psammophile. Lorsque l'eau sort temporairement à marée basse, exposant le sable, puis dans l'épaisseur du sable, dans les intervalles entre les grains de sable, elle demeure, ce qui n'empêche pas l'existence des animaux.
Chez les ciliés, qui font partie de la faune psammophile et appartiennent à divers groupes systématiques (ordres, familles), de nombreux traits communs se développent au cours du processus d'évolution, qui sont des adaptations aux conditions particulières d'existence entre les particules de sable.

La figure 106 montre quelques espèces de la faune psammophile de ciliés appartenant à différents ordres et familles. Il existe de nombreuses similitudes entre eux. Le corps de la plupart d'entre eux est plus ou moins fortement allongé, semblable à un ver. Cela facilite le "serrage" dans les plus petits trous entre les grains de sable. Chez de très nombreuses espèces (Fig. 106) l'allongement du corps se conjugue avec son aplatissement. L'appareil ciliaire est toujours bien développé, ce qui permet aux actifs, avec une certaine force, de se déplacer dans des interstices étroits. Assez souvent, les cils se développent d'un côté du corps aplati en forme de ver, le côté opposé est nu. Cette caractéristique est probablement associée à la capacité, qui est prononcée chez la plupart des espèces psammophiles, de coller (s'attacher) au substrat très étroitement et très fermement à travers l'appareil ciliaire (un phénomène appelé thigmotaxis). Cette propriété permet aux animaux de rester en place dans les cas où des courants d'eau apparaissent dans les interstices étroits où ils vivent. Dans ce cas, il est probablement plus avantageux que le côté opposé à celui sur lequel l'animal s'est fixé sur le substrat soit lisse.
Que mangent les ciliés psammophiles ? Une partie importante du "régime" de nombreuses espèces est constituée d'algues, en particulier de diatomées. Les bactéries leur servent dans une moindre mesure de nourriture. Cela dépend aussi en grande partie du fait qu'il y a peu de bactéries dans les sables qui ne sont pas fortement pollués. Enfin, surtout parmi les plus grands ciliés psammophiles, il existe un nombre considérable de formes prédatrices qui mangent d'autres ciliés appartenant à des espèces plus petites. Les ciliés psammophiles sont distribués, apparemment, partout.
INFUSORIA APOSTOMATES

ciliés spirophrie(Spirophrya subparasitica) à l'état enkysté se trouve souvent assis sur une petite tige sur de petits crustacés planctoniques marins (surtout souvent sur des crustacés du genre Idia). Alors que le crustacé nage activement dans l'eau de mer, la spirophrie qui s'y trouve ne subit aucun changement. Pour le développement ultérieur des ciliés, il est nécessaire que le crustacé soit mangé par un polype hydroïde marin, ce qui arrive assez souvent (Fig. 107). Dès que les kystes de spirophrie, avec le crustacé, pénètrent dans la cavité digestive, de petits ciliés en sortent immédiatement, qui commencent à se nourrir énergiquement de la bouillie alimentaire formée à la suite de la digestion du crustacé avalé. En une heure, la taille des infusoires augmente de 3 à 4 fois. Cependant, la reproduction ne se produit pas à ce stade. Devant nous se trouve un stade de croissance typique des ciliés, appelé trophont. Après un certain temps, avec les résidus alimentaires non digérés, le trophont est rejeté par le polype dans l'eau de mer. Ici, nageant activement, il descend le long du corps du polype jusqu'à sa semelle, où il se fixe, entouré d'un kyste. Ce stade d'un grand cilié enkysté assis sur un polype s'appelle un tomont. C'est la phase de reproduction. Tomont ne se nourrit pas, mais se divise rapidement plusieurs fois de suite (Fig. 107, 7). Le résultat est tout un groupe de très petits ciliés. Leur nombre dépend de la taille du tomont, elle-même déterminée par la taille du trophont qui lui a donné son origine. Les petits ciliés formés à la suite de la division du tomont (on les appelle tomites ou vagabonds) représentent le stade de la colonisation.
Ils sortent du kyste, nagent rapidement (sans manger en même temps, mais en utilisant les réserves qu'ils ont dans le cytoplasme). S'ils ont « la chance » de tomber sur un copépode, ils s'y fixent immédiatement et s'enkystent. C'est à partir de cette étape que nous avons commencé notre réflexion sur le cycle.
Dans la revue cycle de vie spirophria attire l'attention sur la délimitation nette des stades qui ont une signification biologique différente. Trophon est le stade de croissance. Il ne fait que croître, accumule une grande quantité de cytoplasme et toutes sortes de substances de réserve grâce à une nutrition vigoureuse et rapide. Le trophont n'est pas capable de se reproduire. Le phénomène inverse est observé dans le tomont - l'incapacité de se nourrir et une reproduction rapide et vigoureuse. Après chaque division, il n'y a pas de croissance, et donc la reproduction du tomont est réduite à une décomposition rapide en de nombreux vagabonds. Enfin, les vagabonds remplissent leur fonction spéciale et unique caractéristique : ce sont des individus - des colons et des distributeurs de l'espèce. Ils sont incapables de manger ou de se reproduire.
CYCLE DE VIE D'Ichthyophthirius

À la fin de la période de croissance, l'ichtyophthirius atteint une très grande taille par rapport aux vagabonds : 0,5 à 1 mm de diamètre. En atteignant la valeur limite, les ciliés sortent activement des tissus du poisson dans l'eau et nagent lentement pendant un certain temps à l'aide de l'appareil ciliaire couvrant tout leur corps. Bientôt, de grands ichthyophthirius se déposent sur un objet sous-marin et sécrètent un kyste. Immédiatement après l'enkystement, les divisions successives des ciliés commencent : d'abord en deux, puis chaque individu fille est à nouveau divisé en deux, et ainsi de suite jusqu'à 10-11 fois. En conséquence, jusqu'à 2000 petits individus presque arrondis couverts de cils se forment à l'intérieur du kyste. À l'intérieur du kyste, les vagabonds se déplacent activement. Ils perforent la coquille et sortent. Les vagabonds qui nagent activement infectent de nouveaux poissons.
Le taux de division de l'ichtyophthirius dans les kystes, ainsi que le taux de sa croissance dans les tissus des poissons, dépend dans une large mesure de la température. Selon des études de différents auteurs, les chiffres suivants sont donnés : à 26-27°C, le processus de développement des vagabonds dans le kyste prend 10-12 heures, à 15-16°C il prend 28-30 heures, à 4 -5°C il dure 6 à 7 jours.
La lutte contre l'ichtyophthirius présente des difficultés importantes. Les mesures préventives visant à empêcher les vagabonds nageant librement de pénétrer dans les tissus des poissons sont d'une importance primordiale. Pour ce faire, il est utile de procéder à des transplantations fréquentes de poissons malades dans de nouveaux réservoirs ou aquariums, afin de créer des conditions d'écoulement, particulièrement efficaces dans la lutte contre l'ichtyophthirius.
ciliés trichodynes

Tout le système d'adaptation des trichodines à la vie à la surface de l'hôte vise à ne pas se détacher du corps de l'hôte (ce qui équivaut presque toujours à la mort), tout en maintenant la mobilité. Ces appareils sont très parfaits. Le corps de la plupart des trichodinas se présente sous la forme d'un disque plutôt plat, parfois d'une calotte. La face tournée vers le corps de l'hôte est légèrement concave, elle forme une ventouse d'attache. Le long du bord extérieur de la ventouse, il y a une corolle de cils bien développés, à l'aide desquels le mouvement (rampage) des ciliés se produit principalement à la surface du corps du poisson. Cette corolle correspond à la corolle trouvée dans les vagabonds ciliés ronds sessiles discutés ci-dessus. Ainsi, trichodina peut être comparé à un clochard. Sur la surface abdominale (sur la ventouse), les trichodines ont un appareil de support et de fixation très complexe, qui aide à maintenir les ciliés sur l'hôte. Sans entrer dans les détails de sa structure, notons que sa base est un anneau de configuration complexe, constitué de segments séparés portant des dents externes et internes (Fig. 109, B). Cet anneau forme une base à la fois élastique et solide de la surface abdominale, qui agit comme une ventouse. Différents types de trichodine diffèrent les uns des autres par le nombre de segments qui forment l'anneau et par la configuration des crochets externes et internes.
Sur le côté du corps de trichodina opposé au disque, il y a un appareil penné et oral. Sa structure est plus ou moins typique de ciliés circonciliés. Les membranes adorales torsadées dans le sens des aiguilles d'une montre conduisent à un évidement au fond duquel se trouve la bouche. L'appareil nucléaire de la trichodine est généralement arrangé pour les ciliés : un macronoyau en forme de ruban et un micronoyau situé à côté. Il y a une vacuole contractile.
Les trichodines sont largement distribuées dans des réservoirs de tous types. Surtout souvent, ils se trouvent sur les alevins de différentes espèces de poissons. Avec la reproduction de masse, les trichodines sont appliquées grand mal poisson, surtout si les masses recouvrent les branchies. Cela perturbe la respiration normale du poisson.
Afin de nettoyer le poisson de la trichodine, il est recommandé de faire des bains thérapeutiques à partir d'une solution à 2% de chlorure de sodium ou d'une solution à 0,01% de permanganate de potassium (pour les alevins - pendant 10 à 20 minutes).
INFUSOIRES DU TRACTUS INTESTINAL DES UNGATES
De la cicatrice à travers le maillage, la nourriture est rotée dans la cavité buccale, où elle est en outre mâchée (gomme à mâcher). La masse de nourriture mâchée avalée à nouveau par un tube spécial formé par les plis de l'œsophage ne va plus à la cicatrice, mais au livre et de là à la caillette, où elle est exposée aux sucs digestifs du ruminant. Dans la présure, dans des conditions de réaction acide et de présence d'enzymes digestives, les ciliés meurent. En y arrivant avec du chewing-gum, ils sont digérés.
Le nombre de protozoaires dans le rumen (ainsi que dans le filet) peut atteindre des valeurs colossales. Si vous prenez une goutte du contenu de la cicatrice et que vous l'examinez au microscope (lorsqu'elle est chauffée, puisque les ciliés s'arrêtent à température ambiante), les ciliés pullulent littéralement dans le champ de vision. Il est difficile même en culture d'obtenir une telle masse de ciliés. Le nombre de ciliés dans 1 cm3 du contenu de la cicatrice atteint un million, et souvent plus. Au niveau du volume total de la cicatrice, cela donne des chiffres vraiment astronomiques ! La richesse du contenu du rumen en ciliés dépend en grande partie de la nature de l'alimentation du ruminant. Si l'aliment est riche en fibres et pauvre en glucides et en protéines (herbe, paille), alors il y a relativement peu de ciliés dans le rumen. Lorsque des glucides et des protéines (son) sont ajoutés à l'alimentation, le nombre de ciliés augmente considérablement et atteint un nombre énorme. Il faut garder à l'esprit qu'il y a un flux constant de ciliés. Se réunissant avec le chewing-gum dans la caillette, ils meurent. Haut niveau le nombre de ciliés est maintenu par leur reproduction vigoureuse.
Les ongulés aux doigts impairs (cheval, âne, zèbre) possèdent également un grand nombre de ciliés dans le tube digestif, mais leur localisation chez l'hôte est différente. Les ongulés aux doigts impairs n'ont pas d'estomac complexe, en raison duquel la possibilité de développer des protozoaires dans les parties antérieures du tube digestif est absente. Mais chez les équidés, le gros et le caecum sont très bien développés, qui sont généralement obstrués par des masses alimentaires et jouent un rôle essentiel dans la digestion. Dans cette partie de l'intestin, tout comme dans le rumen et les mailles des ruminants, se développe une très riche faune de protozoaires, principalement des ciliés, dont la plupart appartiennent également à l'ordre des endodiniomorphes. Cependant, en termes de composition spécifique, la faune du rumen des ruminants et la faune du gros intestin des équidés ne coïncident pas.
CILICIENS DU TRACTUS INTESTINAL DES RUMINANTS
Les ciliés sont les plus intéressants famille des ofrioscolécides(Ophryoscolecidae), apparenté à ordre endodiniomorphe. Un trait caractéristique de ce détachement est l'absence de couverture ciliaire continue. Les formations ciliaires complexes - cirres - sont situées à l'avant du corps des ciliés dans la région de l'ouverture de la bouche. A ces éléments de base de l'appareil ciliaire peuvent s'ajouter des groupes supplémentaires de cirrhées, situés soit à l'extrémité antérieure, soit à l'extrémité postérieure du corps. Le nombre total d'espèces de ciliés de la famille des ofrioscolécidés est d'environ 120.

La figure 110 montre quelques-uns des rioscolécides les plus typiques du rumen des ruminants. Les ciliés du genre Entodinium (Entodinium, fig. 110, L) sont plus simplement disposés. À l'extrémité antérieure de leur corps, il y a une zone péribuccale de cirrhus. L'extrémité avant du corps, sur laquelle se trouve l'ouverture de la bouche, peut être tirée vers l'intérieur. L'ectoplasme et l'endoplasme sont nettement délimités. Le tube anal est clairement visible à l'extrémité postérieure, qui sert à éliminer les résidus alimentaires non digérés. Structure un peu plus complexe anoplodynie(Anoplodinium, fig. 110, B). Ils ont deux zones de l'appareil ciliaire - cirrhi périoral et cirrhi dorsal. Les deux sont situés à l'avant. À l'arrière du corps de l'espèce représentée sur la figure, il y a de longues excroissances pointues - c'est assez typique pour de nombreuses espèces d'ofrioscolécidés. Il a été suggéré que ces excroissances contribuent à la "poussée" des ciliés entre les particules végétales qui remplissent la cicatrice.
Sortes Genre Eudiplodynia(Eudiplodinium, fig. 110, B) sont similaires à anoplodynie, mais, contrairement à eux, ont une plaque de base squelettique située sur le bord droit le long du pharynx. Cette plaque squelettique se compose d'une substance similaire dans nature chimiqueà la fibre, c'est-à-dire à la substance qui compose les coquilles des cellules végétales.
À genre polyplastron(Polyplastron, fig. 110, D, E) il y a une autre complication du squelette. La structure de ces ciliés est proche de l'eudiplodynie. Les différences se résument principalement au fait qu'au lieu d'une plaque squelettique, ces ciliés en ont cinq. Deux d'entre eux, les plus grands, sont situés sur le côté droit et trois, les plus petits, sont sur le côté gauche du cilié. La deuxième caractéristique du polyplastron est une augmentation du nombre de vacuoles contractiles. L'entodynie a une vacuole contractile, l'anoplodynie et l'eudiplodynie ont deux vacuoles contractiles et le polyplastron en a environ une douzaine.
À épidinium(Epidinium, Fig. 110), qui ont un squelette glucidique bien développé situé sur le côté droit du corps, la zone dorsale du cirrhus se déplace de l'extrémité antérieure vers la face dorsale. Les épines se développent souvent à l'extrémité postérieure des ciliés de ce genre.
La structure la plus complexe révèle genre Ofrioscolex(Ophryoscolex), après quoi toute la famille des ciliés est nommée (Fig. 110, E). Ils ont une zone dorsale bien développée de cirrhus, couvrant environ les 2/3 de la circonférence du corps et des plaques squelettiques. De nombreuses épines sont formées à l'extrémité postérieure, dont une est généralement particulièrement longue.
Connaissance de quelques représentants typiques ofrioscolécide montre qu'au sein de cette famille il y a eu une complication importante de l'organisation (de l'entodynie à l'ofrioscolex).
En plus des ciliés famille des ofrioscolécides, dans le rumen des ruminants, des représentants de ceux que nous connaissons déjà se trouvent en petites quantités. détachement des ciliés isociliaires. Ils sont représentés par un petit nombre d'espèces. Leur corps est uniformément recouvert de rangées longitudinales de cils, les éléments squelettiques sont absents. Dans la masse totale de la population ciliée du rumen, ils ne jouent pas un rôle significatif, nous ne nous y attarderons donc pas ici.
Que mangent et comment les ciliés des rioscolécides mangent-ils ? Cette question a été étudiée en détail par de nombreux scientifiques, en particulier en détail par le professeur V. A. Dogel.

La nourriture des ofrioscolécides est assez diversifiée et une certaine spécialisation est observée chez différentes espèces. Les plus petites espèces du genre Entodynia se nourrissent de bactéries, de grains d'amidon, de champignons et d'autres petites particules. De très nombreux ofrioscolécides moyens et grands absorbent les particules de tissus végétaux, qui constituent l'essentiel du contenu du rumen. L'endoplasme de certaines espèces est littéralement obstrué par des particules végétales. Vous pouvez voir comment les ciliés bondissent sur des bouts de tissus végétaux, les déchirent littéralement en morceaux puis les avalent, les tordant souvent en spirale dans leur corps (Fig. 111, 4). Parfois, il est nécessaire d'observer de telles images (Fig. 111, 2), lorsque le corps du cilié lui-même est déformé en raison de grosses particules avalées.
Chez les ofrioscolécidés, une prédation est parfois observée. Les grandes espèces dévorent les plus petites. La prédation (Fig. 112) se conjugue à la capacité d'une même espèce à se nourrir de particules végétales.

Comment les ciliés pénètrent-ils dans le rumen d'un ruminant ? Quelles sont les voies d'infection par les ofrioscolécides ? Il s'avère que les ruminants nouveau-nés n'ont pas encore de ciliés dans le rumen. Ils sont également absents lorsque l'animal se nourrit de lait. Mais dès que le ruminant passe à la nourriture végétale, des ciliés apparaissent immédiatement dans la cicatrice et le maillage, dont le nombre augmente rapidement. D'où viennent-ils? Pendant longtemps, on a supposé que les ciliés du rumen formaient une sorte de stades de repos (très probablement des kystes), qui sont largement dispersés dans la nature et, lorsqu'ils sont avalés, donnent naissance à des stades actifs de ciliés. D'autres études ont montré que les ciliés des ruminants n'ont pas de stades de repos. Il a été possible de prouver que l'infection se produit avec des ciliés mobiles actifs qui pénètrent dans la cavité buccale lors de l'urine. Si vous examinez sous un microscope pris de cavité buccale chewing-gum, mais il a toujours un grand nombre de ciliés flottants. Ces formes actives peuvent facilement pénétrer dans la bouche et plus loin dans le rumen d'autres ruminants à partir d'un récipient commun pour boire, ainsi que de l'herbe, du foin (qui peut contenir de la salive avec des ciliés), etc. Cette voie d'infection a été prouvée expérimentalement.
S'il n'y a pas d'étapes de repos chez les ofrioscolécides, il est évidemment facile d'obtenir des animaux "sans infuseur" en les isolant alors qu'ils se nourrissent encore de lait. Si le contact direct n'est pas autorisé entre les jeunes en croissance et les ruminants avec des ciliés, les jeunes animaux peuvent être laissés sans ciliés dans le rumen. De telles expériences ont été menées par plusieurs scientifiques dans différents pays. Le résultat était clair. En l'absence de contact entre les jeunes (prélevés à la mère pendant la période d'allaitement) et les ruminants avec des ciliés dans le rumen, les animaux grandissent stériles par rapport aux ciliés. Cependant, même un contact de courte durée avec des animaux porteurs de ciliés (une mangeoire commune, un seau commun pour boire, un pâturage commun) suffit pour que des ciliés apparaissent dans le rumen des animaux stériles.
Ci-dessus se trouvaient les résultats d'expériences sur le contenu de ruminants, complètement dépourvus de ciliés dans le rumen et le filet. Ceci est réalisé, comme nous l'avons vu, par un isolement précoce des jeunes. Des expériences ont été menées sur des moutons et des chèvres.
De cette façon, il a été possible de mener des observations d'animaux "sans infuseur" pendant une période de temps significative (plus d'un an). Comment l'absence de ciliés dans le rumen affecte-t-elle la vie de l'hôte ? L'absence de ciliés affecte-t-elle l'hôte négativement ou positivement ? Pour répondre à cette question, les expériences suivantes ont été réalisées sur des chèvres. Des chevreaux jumeaux (de la même portée et du même sexe) ont été prélevés afin d'avoir plus de matériel similaire. Ensuite, l'un des jumeaux de ce couple a été élevé sans ciliés dans le rumen (isolement précoce), tandis que l'autre, dès le début de l'alimentation des aliments végétaux, a été abondamment infecté par de nombreux types de ciliés. Tous deux recevaient exactement le même régime alimentaire et étaient élevés dans les mêmes conditions. La seule différence entre eux était la présence ou l'absence de ciliés. Sur plusieurs paires de chevreaux ainsi étudiés, aucune différence n'a été constatée au cours du développement des deux membres de chaque paire ("infuseur" et "non-infuseur"). Ainsi, on peut affirmer que les ciliés vivant dans le rumen et le filet n'ont pas d'effet marqué sur les fonctions vitales de l'animal hôte.
Les résultats ci-dessus des expériences ne permettent cependant pas d'affirmer que les ciliés du rumen sont complètement indifférents au propriétaire. Ces expériences ont été réalisées avec un régime alimentaire normal de l'hôte. Il est possible que dans d'autres conditions, avec un régime alimentaire différent (par exemple, avec une alimentation insuffisante), il soit possible de révéler l'effet sur l'hôte de la faune des protozoaires habitant le rumen.
Diverses suggestions ont été faites dans la littérature sur l'éventuelle influence positive de la faune protozoaire du rumen sur les processus digestifs de l'hôte. Il a été souligné que plusieurs millions de ciliés, nageant activement dans le rumen et écrasant les tissus végétaux, contribuent à la fermentation et à la digestion des masses alimentaires situées dans les parties antérieures du tube digestif. Un nombre important de ciliés, qui pénètrent dans la caillette avec le chewing-gum, sont digérés et la protéine, qui constitue une partie importante du corps des ciliés, est absorbée. Les infusoires peuvent donc être une source supplémentaire de protéines pour l'hôte. Il a également été suggéré que les ciliés contribuent à la digestion des fibres, qui constituent l'essentiel de l'alimentation des ruminants, et à leur transfert vers un état plus digeste.
Toutes ces hypothèses ne sont pas prouvées et des objections ont été soulevées contre certaines d'entre elles. Il a été souligné, par exemple, que les ciliés construisent le protoplasme de leur corps à partir de protéines qui pénètrent dans le rumen avec la nourriture de l'hôte. Absorbant les protéines végétales, ils les transforment en protéines animales de leur corps, qui sont ensuite digérées dans la caillette. On ne sait pas si cela procure un avantage à l'hôte. Toutes ces questions présentent un grand intérêt pratique, puisque nous parlons de la digestion des ruminants - les principaux objets de l'élevage. Des recherches plus approfondies sur le rôle des ciliés du rumen dans la digestion des ruminants sont hautement souhaitables.
Les ofrioscolécides de ruminants ont, en règle générale, une large spécificité. En termes d'espèces, la population de la cicatrice et du filet de bovins, ovins et caprins est très proche l'une de l'autre. Si nous comparons la composition spécifique du rumen des antilopes africaines avec celle des bovins, ici aussi, environ 40% du nombre total d'espèces seront communes. Cependant, il existe de nombreuses espèces d'ofrioscolécides que l'on ne trouve que chez les antilopes ou uniquement chez les cerfs. Ainsi, dans le contexte de la large spécificité générale des ofrioscolécides, on peut parler de types distincts, plus étroitement spécifiques.
Infusoires des intestins des équidés
Passons maintenant à une brève connaissance des ciliés habitant le grand et le caecum des équidés.
En termes d'espèces, cette faune, comme la faune du rumen des ruminants, est également très diversifiée. Actuellement, environ 100 espèces de ciliés ont été décrites qui vivent dans le gros intestin des animaux de la famille équine. Les ciliés retrouvés ici au sens de leur appartenance à des groupes systématiques différents sont plus diversifiés que les ciliés du rumen des ruminants.

Dans les intestins des chevaux, il existe de nombreuses espèces de ciliés appartenant à l'ordre des isociliaires, c'est-à-dire des ciliés dans lesquels l'appareil ciliaire ne forme pas de membranella ou de cirrhose près de la zone buccale (Fig. 113, 1).
Commandez l'entodyniomorphe(Entodiniomorpha) est également richement représentée dans les intestins du cheval. Alors qu'une seule famille d'endodiniomorphes (famille des ofrioscolécides) se retrouve dans le rumen des ruminants, des représentants de trois familles vivent dans les intestins d'un cheval, mais nous ne nous attarderons pas ici sur les caractéristiques dont nous nous limitons ici à quelques dessins de espèce de cheval typique (Fig. 113) .
Des études détaillées menées par A. Strelkov ont montré que les différents types de ciliés sont loin d'être uniformément répartis le long du gros intestin du cheval. Il y a deux groupes d'espèces différents, deux faunes, pour ainsi dire. L'un d'eux habite le caecum et la partie abdominale du gros côlon (les sections initiales du gros intestin), et l'autre habite la partie dorsale du gros côlon et du petit côlon. Ces deux complexes d'espèces sont assez nettement délimités. Il y a peu d'espèces communes à ces deux sections - moins d'une douzaine.
, 
Il est intéressant de noter que parmi les nombreuses espèces de ciliés habitant le gros intestin des équidés, il existe des représentants du même genre apparentés aux ciliés suceurs. Comme nous l'avons vu plus haut, ciliés suceurs(Suctoria) sont des organismes sessiles typiques vivant librement avec une façon très spéciale de se nourrir à l'aide de tentacules (Fig. 103). Un des succion de l'accouchement adapté à un habitat aussi inhabituel que le gros intestin d'un cheval, par exemple, plusieurs espèces allantose(Allantosome). Ces animaux très particuliers (Fig. 114) n'ont pas de pédoncule, les cils sont absents, les tentacules en massue épaissies aux extrémités sont bien développées.
À l'aide de tentacules, les allantosomes adhèrent à différents types de ciliés et les aspirent. Souvent, la proie est plusieurs fois plus grande que le prédateur.
La question de la nature des relations entre les ciliés du gros intestin des équidés et leurs hôtes reste encore floue. Le nombre de ciliés peut être aussi élevé, et parfois même plus, que dans le rumen des ruminants. Il existe des données montrant que la quantité de ciliés dans le gros intestin d'un cheval peut atteindre 3 millions dans 1 cm3. La signification symbiotique suggérée par certains scientifiques est encore moins probable que pour les ciliés du rumen.
L'opinion la plus probable est qu'ils causent des dommages à l'hôte, absorbant une quantité importante de nourriture. Une partie des ciliés est extraite avec des masses fécales, et ainsi les substances organiques (y compris les protéines) qui composent leur corps restent inutilisées par l'hôte.
La question des voies d'infection des équidés par des ciliés habitant le gros intestin n'a pas encore été résolue.

Balantidia capture une variété de particules alimentaires du contenu du gros intestin. Surtout volontiers, il se nourrit de grains d'amidon. Si la balantidia vit dans la lumière du côlon humain, elle se nourrit du contenu de l'intestin et n'a aucun effet nocif. Il s'agit d'un "chariot" typique, que nous avons déjà rencontré lors de l'examen de l'amibe dysentérique. Cependant, la balantidia est moins susceptible que l'amibe dysentérique de rester un "locataire inoffensif".
À l'heure actuelle, les spécialistes ont bien développé diverses méthodes, permettant la culture de balantidia dans un environnement artificiel - en dehors de l'organisme hôte.

Comme on peut le voir sur la figure, troglodptella est l'un des endodiniomorphes complexes. Elle, en plus de la zone périorale de la cirrhose (à l'extrémité antérieure du corps), a trois autres zones de cirrhi bien développés, recouvrant annulairement le corps du cilié. Les trogloditells ont un appareil squelettique bien développé composé d'hydrates de carbone, couvrant presque toute l'extrémité antérieure du corps. Les tailles de ces ciliés particuliers sont assez importantes. En longueur, ils atteignent 200-280 microns.
ASTOMATES ciliés sans bouche

Les formations squelettiques de soutien se développent principalement à l'extrémité antérieure du corps, qui doit subir des contraintes mécaniques et surmonter des obstacles, poussant à travers la lumière intestinale entre les particules alimentaires. Espèces genre radiophrie(Radiophrya) à l'extrémité antérieure d'un côté du corps (qui est conventionnellement considéré comme le côté abdominal) il y a de très fortes côtes élastiques (spicules) situées dans la couche superficielle de l'ectoplasme (Fig. 117, B, D, E) . Espèces Genre Menilella(Mesnilella) il existe également des rayons de soutien (spicules) qui, pour la majeure partie de leur longueur, se situent dans les couches profondes du cytoplasme (dans l'endoplasme, fig. 117, A). Des formations de support disposées de la même manière sont également développées dans des espèces de certains autres genres Astomat.
La reproduction asexuée chez certains astomates ciliés se déroule d'une manière particulière. Au lieu de la division transversale en deux, caractéristique de la plupart des ciliés, de nombreux astomates ont une division inégale (bourgeonnement). Dans le même temps, les reins qui se séparent à l'extrémité postérieure restent associés à l'individu mère pendant un certain temps (Fig. 117, B). En conséquence, des chaînes sont obtenues, constituées d'individus antérieurs grands et postérieurs plus petits (reins). À l'avenir, les reins se séparent progressivement de la chaîne et passent à une existence indépendante. Cette forme particulière de reproduction est répandue, par exemple, dans la radiophrie déjà connue de nous. Les chaînes de certains astomes résultant du bourgeonnement ressemblent apparence chaînes de ténias. Ici encore, nous rencontrons le phénomène de convergence.
L'appareil nucléaire de l'astomat a une structure caractéristique des ciliés : un macronoyau, le plus souvent en forme de ruban (Fig. 117), et un micronoyau. Les vacuoles contractiles sont généralement bien développées. La plupart des espèces ont plusieurs (parfois plus d'une douzaine) vacuoles contractiles disposées en une rangée longitudinale.
Une étude de la distribution des espèces d'Astomat par différents types d'hôtes montre que la plupart des espèces d'Astomat sont confinées à des espèces hôtes strictement définies. La plupart des astomates se caractérisent par une spécificité étroite : une seule espèce animale peut leur servir d'hôte.
Malgré le grand nombre d'études consacrées à l'étude des ciliés astomatiques, un aspect très important de leur biologie reste totalement flou : comment s'effectue la transmission de ces ciliés d'un individu hôte à un autre ? Il n'a jamais été possible d'observer la formation de kystes chez ces ciliés.
Par conséquent, il est suggéré que l'infection se produise activement - stades mobiles.
INFUSOIRES DE L'INTESTIN D'OURSIN DE MER
Les oursins sont très nombreux dans la zone côtière de nos mers du nord (Barents) et d'Extrême-Orient (la mer du Japon, la côte pacifique des îles Kouriles). Majorité oursins se nourrit d'aliments végétaux, principalement des algues, qu'ils grattent sur les objets sous-marins avec des "dents" acérées spéciales entourant l'ouverture de la bouche. Dans les intestins de ces hérissons herbivores se trouve une riche faune de ciliés. Souvent, ils se développent ici en masse et le contenu des intestins de l'oursin au microscope est presque aussi "grouillant" d'infusoires que le contenu du rumen des ruminants. Il faut dire qu'en plus de profondes différences dans les conditions de vie des ciliés des intestins de l'oursin et du rumen du ruminant, il existe quelques similitudes. Elles résident dans le fait qu'ici et là les ciliés vivent dans un milieu très riche en restes végétaux. Actuellement, plus de 50 espèces de ciliés sont connues pour vivre dans les intestins des oursins, qui ne se trouvent que dans la zone côtière, où les oursins se nourrissent d'algues. Aux grandes profondeurs, là où les algues ne poussent plus, il n'y a pas de ciliés chez les oursins.

Selon le mode de vie et la nature de la nutrition, la plupart des ciliés des intestins des oursins sont herbivores. Ils se nourrissent d'algues, qui remplissent en grande quantité les intestins de l'hôte. Certaines espèces sont assez "tatillonnes" dans leur choix de nourriture. Par example, strobilidium(Strobilidium, fig. 118, A) se nourrit presque exclusivement de grosses diatomées. Il y a aussi des prédateurs qui mangent des représentants d'autres, plus petite espèce ciliés.
Dans les ciliés des intestins des oursins, contrairement à l'astomat, il n'y a pas de confinement strict à certains types d'hôtes. Ils vivent dans une grande variété d'oursins mangeurs d'algues.
Les voies d'infection des oursins par les ciliés n'ont pas été étudiées. Cependant, ici avec un degré de probabilité élevé, on peut supposer qu'il se produit sous des formes actives flottantes. Le fait est que les ciliés des intestins des oursins peuvent vivre longtemps (plusieurs heures) dans l'eau de mer. Pourtant, ils se sont déjà tellement adaptés à la vie dans les intestins des hérissons qu'en dehors de leur corps, dans l'eau de mer, tôt ou tard ils meurent.
En terminant la connaissance des ciliés, il convient de souligner une fois de plus qu'ils représentent une riche en espèces, un groupe (classe) étendu et prospère du monde animal. Restant au niveau de l'organisation cellulaire, les infusoires ont atteint, par rapport aux autres classes de protozoaires, la plus grande complexité de structure et de fonctions.
Un rôle particulièrement important dans ce développement progressif (évolution) a probablement été joué par la transformation de l'appareil nucléaire et l'émergence du dualisme nucléaire (non-équivalence qualitative des noyaux). La richesse du macronoyau en substances nucléiques est associée à des processus métaboliques actifs, à des processus vigoureux de synthèse de protéines dans le cytoplasme et les noyaux.
Conclusion
Nous sommes arrivés à la fin de notre examen de la structure et du mode de vie d'un type étendu de monde animal - protozoaires. Leur trait caractéristique, comme cela a été souligné à plusieurs reprises ci-dessus, est l'unicellularité. En termes de structure, les protozoaires sont des cellules. Cependant, ils sont incomparables avec les cellules qui composent le corps des organismes multicellulaires, car ils sont eux-mêmes des organismes. Ainsi, les protozoaires sont des organismes au niveau d'organisation cellulaire. Certains protozoaires très organisés, possédant de nombreux noyaux, semblent déjà dépasser les limites morphologiques de la structure cellulaire, ce qui donne à certains scientifiques des raisons de qualifier ces protozoaires de "supracellulaires". Cela change peu le fond des choses, car une organisation unicellulaire est encore typique des Protozoaires.
Dans les limites de l'unicellularité, les protozoaires ont parcouru un long chemin de développement évolutif et ont donné une grande variété de formes adaptées aux conditions de vie les plus diverses. Au cœur de la tige généalogique des protozoaires se trouvent deux classes : les sarcodes et les flagellés. La question de savoir laquelle de ces classes est la plus primitive est encore débattue en science. D'une part, les représentants inférieurs des Sarcodidae (ameba) ont la structure la plus primitive. Mais les flagellés présentent la plus grande plasticité du type de métabolisme et se situent en quelque sorte à la frontière entre les mondes animal et végétal. Dans le cycle de vie de certains sarcodes (par exemple, les foraminifères), il existe des stades flagellaires (gamètes), ce qui indique leur relation avec les flagellés. Il est évident que ni les sarcodes modernes ni les flagellés modernes ne peuvent constituer le groupe initial de l'évolution du monde animal, car ils ont eux-mêmes parcouru un long chemin dans le développement historique et ont développé de nombreuses adaptations aux conditions de vie modernes sur Terre. Probablement, ces deux classes de protozoaires modernes doivent être considérées comme deux troncs en évolution, issus de formes anciennes qui n'ont pas survécu à ce jour, qui ont vécu à l'aube du développement de la vie sur notre planète.
Dans l'évolution ultérieure des protozoaires, des changements de nature différente se sont produits. Certains d'entre eux ont conduit à une augmentation générale du niveau d'organisation, à une augmentation de l'activité et à l'intensité des processus de la vie. Parmi ces transformations phylogénétiques (évolutives), il faut, par exemple, inclure le développement des organites du mouvement et de la capture alimentaire, qui a atteint un haut niveau de perfection dans la classe des ciliés. Il est incontestable que les cils sont des organites correspondant (homologues) aux flagelles. Alors que chez les flagellés, à quelques exceptions près, le nombre de flagelles est faible, chez les ciliés, le nombre de cils a atteint plusieurs milliers. Le développement de l'appareil ciliaire a fortement augmenté l'activité des protozoaires, rendu les formes de leur relation avec l'environnement plus diverses et complexes, les formes de réactions aux stimuli externes. La présence d'un appareil ciliaire différencié a sans aucun doute été l'une des principales raisons de l'évolution progressive de la classe des ciliés, où une grande variété de formes adaptées à différents habitats est apparue.
Le développement de l'appareil ciliaire des ciliés est un exemple de ce type de changement évolutif, nommé par Acad. Aromorphoses de Severtsov. Les aromorphoses se caractérisent par une augmentation générale de l'organisation, le développement d'adaptations d'une grande importance. Une augmentation de l'organisation est comprise comme des changements qui provoquent une augmentation de l'activité vitale de l'organisme; ils sont associés à la différenciation fonctionnelle de ses parties, conduisant à des formes de communication plus diverses entre l'organisme et l'environnement. Le développement de l'appareil ciliaire des ciliés fait précisément référence à ce type de transformations structurelles en cours d'évolution. Il s'agit d'une aromorphose typique.
Chez les protozoaires, comme l'a souligné V. A. Dogel, les modifications du type d'aromorphoses sont généralement associées à une augmentation du nombre d'organites. La polymérisation des organites se produit. Le développement de l'appareil ciliaire chez les ciliés est un exemple typique de tels changements. Le deuxième exemple d'aromorphose dans l'évolution des ciliés est leur appareil nucléaire. Nous avons examiné ci-dessus les caractéristiques structurelles du noyau des ciliés. Le dualisme nucléaire des ciliés (présence d'un micronoyau et d'un macronoyau) s'accompagne d'une augmentation du nombre de chromosomes dans le macronoyau (phénomène de polyploïdie). Comme les chromosomes sont associés aux principaux processus de synthèse dans la cellule, principalement à la synthèse des protéines, ce processus conduit à une augmentation générale de l'intensité des principales fonctions vitales. Ici aussi, une polymérisation a eu lieu, affectant les complexes chromosomiques du noyau.
ciliés- l'un des groupes de protozoaires les plus nombreux et les plus progressifs, provient des flagellés. Ceci est démontré par la similitude morphologique complète de leurs organoïdes de mouvement. Ce stade d'évolution était associé à deux grandes aromorphoses: l'une affectait les organites du mouvement, la seconde - l'appareil nucléaire. Ces deux types de changements sont liés, car ils entraînent tous deux une augmentation de l'activité vitale et une complication des formes d'interrelations avec l'environnement extérieur.
Parallèlement aux aromorphoses, il existe un autre type de changement évolutif, exprimé dans le développement d'adaptations (adaptation) à certaines conditions d'existence bien définies. Ce type de changement évolutif a été appelé idioadaptation par Severtsov. Dans l'évolution des protozoaires, ce type de changement a joué un rôle très important. Ci-dessus, en considérant différentes classes protozoaires, de nombreux exemples de changements idioadaptatifs sont donnés. Adaptations à un mode de vie planctonique dans différents groupes de protozoaires, adaptations à la vie dans le sable chez les ciliés, formation de coquilles protectrices d'oocystes chez les coccidies, et bien plus encore - ce sont toutes des idioadaptations qui ont joué un rôle important dans l'émergence et le développement de groupes individuels, mais ne sont pas associés à des changements progressifs généraux dans l'organisation .
Les adaptations à divers habitats spécifiques chez les protozoaires sont très diverses. Ils ont assuré la large distribution de ce type dans une grande variété d'habitats, ce qui a été discuté en détail ci-dessus, lors de la description des classes individuelles.
Grande encyclopédie médicale
Infusoria Tetrahymena thermophila ... Wikipédia
- (Infusoria) une classe de protozoaires (Protozoa) les plus développés. Les principales caractéristiques d'I.: la présence de cils (pour le mouvement et la nutrition), deux types de noyaux (macronoyau polyploïde et micronoyau diploïde, de structure et ... ... Grande Encyclopédie soviétique
Ou animaux liquides (Infusoria) une classe d'animaux protozoaires, dotés de tresses ou de cils chatoyants, remplacés parfois à l'âge adulte par des excroissances suceuses cylindriques, avec une certaine forme de corps, pour la plupart avec ... ... Dictionnaire encyclopédique F. Brockhaus et I.A. Efron
Taxonomie de la classification ifusoria et systématisation des organismes appartenant au type des ciliés, conformément au concept général de taxonomie des protozoaires en particulier, et des animaux en général Sommaire 1 Systématique 1.1 Classique ... Wikipedia
Ou animaux liquides (Infusoria) une classe d'animaux protozoaires, équipés de tresses ou de cils chatoyants, qui à l'état adulte sont parfois remplacés par des excroissances suceuses cylindriques, avec une certaine forme de corps, pour la plupart avec ... ... Encyclopédie de Brockhaus et Efron
Le type de ciliés est aussi communément appelé cilié - les organes de mouvement de ces protozoaires sont cils. La cellule ciliée a deux noyaux, ils sont appelés petit et grand. Le premier régule le processus de reproduction et le second est responsable des processus de nutrition, de mouvement et de respiration.
Les caractéristiques de la vie de ce type doivent être considérées sur l'exemple des chaussures ciliées.
Mouvement et respiration
L'infusoire-chaussure, dont la longueur est d'environ 0,5 mm, choisit les plans d'eau comme habitat. La forme du corps du plus simple est facile à deviner par son nom - elle ressemble à une chaussure. La vitesse de déplacement est d'environ 2,5 mm par seconde.
La présence d'une coque extérieure élastique offre une forme corporelle stable.
Dans le cytoplasme, qui est adjacent à la coquille, il y a des fibres de soutien, leur développement est une garantie de la préservation de la forme constante du cilié.
À la surface des ciliés, il y a 15 000 cils, à leur base il y a un corps basal. Le mouvement se produit à l'aide des vibrations des cils : ils produisent environ 30 coups par seconde, poussant ainsi la chaussure ciliée vers l'avant.
Il respire à travers la surface du corps.
La nutrition
Une caractéristique des ciliés est la présence bouche de cellule, près de laquelle se trouvent des cils particulièrement longs et denses. La bouche cellulaire continue avec le pharynx cellulaire: les cils poussent l'eau et la nourriture ciliée - les bactéries à l'intérieur.
Les infusoires détectent les produits chimiques, ce qui libère un groupe de bactéries. Ainsi, elle cherche une proie.
La nourriture se retrouve ensuite dans la vacuole digestive où elle est digérée. De là, il suit déjà dans le cytoplasme.
Sélection
La sélection s'effectue à l'aide deux vacuoles contractiles, l'un est situé à l'avant et l'autre à l'arrière. Les vacuoles sont constituées d'un réservoir et de canaux.
Le liquide remplit les canaux, puis suit le réservoir central, puis sort des infusoires. Le processus de contraction des vacuoles prend 10 à 20 secondes.
la reproduction
Les infusoires se reproduisent de manière asexuée - elles sont divisées en deux. Sa caractéristique est la division à travers le corps.
Les noyaux des ciliés sont divisés en deux parties : les ciliés nouvellement formés ont un petit et un gros noyau. Les ciliés filles possèdent des parties d'organoïdes et les parties manquantes sont formées indépendamment. La reproduction a lieu plusieurs fois par jour.
Pour les ciliés-chaussures, la reproduction sexuée est également possible, mais dans ce cas, il n'y a pas d'augmentation du nombre d'individus. Temporairement, les protozoaires sont connectés, formant un pont de connexion à partir du cytoplasme.
Dans chaque individu, le gros noyau disparaît et les petits sont divisés deux fois - quatre noyaux apparaissent. De ceux-ci, il ne reste qu'un seul noyau, qui se divise également. Un individu contient deux noyaux, puis un échange de noyaux se produit - l'un des noyaux se déplace vers un autre individu.
Là, il fusionne avec le noyau qui reste, et ainsi un noyau petit et plus grand se forme dans chacun des individus. Ce processus, appelé conjugaison, est nécessaire au renouvellement du matériel génétique entre les individus.
Types de ciliés
Les ciliés sont des protozoaires organisés de manière complexe, il en existe environ 7 000 espèces.
1. Variété de sarcodes.
2. Variété de flagelles.
Les représentants du deuxième sous-type se déplacent à l'aide de flagelles, ce qui se reflète dans le nom de ce groupe.
- Flagellés. Le vert Euglena est un représentant typique de ce groupe. Ce protozoaire a une forme fusiforme, recouverte d'une membrane élastique dense. À l'extrémité antérieure du corps se trouve un flagelle, près de la base duquel se trouve un œil sensible à la lumière - un stigmate et une vacuole contractile. Plus près de l'extrémité postérieure, dans l'épaisseur du cytoplasme, se trouve un noyau qui contrôle tous les processus vitaux du corps. La couleur verte d'Euglena est due à de nombreux chloroplastes. À la lumière, ce flagellé effectue la photosynthèse, mais étant longtemps dans les zones non éclairées du réservoir, il passe à se nourrir des produits de désintégration de substances organiques complexes, en les extrayant de l'environnement. Ainsi, l'euglène verte peut combiner à la fois un métabolisme de type autotrophe et hétérotrophe. L'existence d'organismes avec un tel type de nutrition mixte indique la relation entre les mondes animal et végétal. La plupart des flagelles
3. Variété de sporozoaires et de ciliés.
Le type cilié comprend les protozoaires, qui se distinguent par l'organisation la plus complexe parmi les animaux unicellulaires. Un représentant typique de ce groupe est la chaussure d'infusoire - un habitant commun de l'eau douce. Son corps est recouvert d'une coquille dense - pellicule et a donc une forme relativement constante. Un trait caractéristique de la structure est la présence de cils, couvrant uniformément tout le corps de la chaussure. Les mouvements des cils sont coordonnés grâce au réseau de fibres contractiles situées dans la couche superficielle du cytoplasme. La deuxième caractéristique est la présence de deux noyaux, gros (macronoyau) et petit (micronoyau). Les noyaux diffèrent également sur le plan fonctionnel: un gros régule le métabolisme et un petit participe au processus sexuel (conjugaison). L'infusoire-chaussure se nourrit de bactéries, des algues unicellulaires, qui sont ajustées par des cils à la bouche cellulaire, qui se trouve au fond de la cavité pré-orale. Après avoir traversé le pharynx cellulaire, qui se termine dans le cytoplasme, les particules alimentaires sont enfermées dans des vacuoles digestives, où elles sont décomposées par l'action d'enzymes. Les résidus non digérés sont rejetés à travers la poudre. Dans le corps de la chaussure, il y a deux vacuoles contractiles complexes pulsant alternativement. Les chaussures se reproduisent de manière asexuée, en se divisant en deux. La reproduction asexuée multiple est remplacée dans la chaussure par le processus sexuel - la conjugaison, au cours de laquelle deux chaussures se rejoignent et échangent du matériel génétique. Après cela, les chaussures divergent et recommencent bientôt la reproduction asexuée. La signification biologique de la conjugaison réside dans la combinaison dans un même organisme des propriétés héréditaires de deux individus. Cela augmente sa viabilité, qui se traduit par une meilleure adaptabilité aux conditions environnementales.
Vous n'avez pas trouvé ce que vous cherchiez ? Utilisez la recherche
Sur cette page, du matériel sur les sujets :
- brièvement sur les flagellés, les pseudopodes, les sporozoaires et les ciliés
- sarcodes et sporozoaires
- variété de flagelles
- résumé du rapport sur les pantoufles d'infusoires
- quelle est la différence entre les ciliés et les espèces de sarcode
L'un des représentants les plus connus du ciliaire est la chaussure ciliée. Il vit, en règle générale, dans l'eau d'une direction permanente, ainsi que dans des réservoirs de type eau douce, où le courant se distingue par l'exception de l'affirmation de soi. Son habitat doit nécessairement contenir de la matière organique en décomposition. Il conviendrait d'examiner en détail tous les aspects de la vie de ce représentant de la faune.
Représentants du cil

A noter que les ciliés sont un type dont le nom vient du mot "teinture" (traduit de Latin). Cela peut s'expliquer par le fait que les premiers représentants des protozoaires ont été trouvés précisément dans les teintures à base de plantes. Au fil du temps, le développement de ce type a commencé à prendre rapidement de l'ampleur. Ainsi, déjà aujourd'hui en biologie, environ 6 à 7 000 espèces sont connues, y compris le type de ciliés. Si nous nous appuyons sur les données des années 1980, on peut alors affirmer que le type en question contient deux classes dans sa structure : les ciliés ciliés (a trois superordres) et les ciliés suceurs. En lien avec ces informations, on peut conclure que la diversité des organismes vivants est très large, ce qui est d'un réel intérêt.
Type d'Infusoires : représentants

Les représentants éminents de ce type sont les ciliés-balantidies et les ciliés-chaussures. Caractéristiques distinctives Ces animaux sont le revêtement de la pellicule avec des cils, qui sont utilisés pour le mouvement, la protection des ciliés par des organes spécialement conçus, les trichocystes (situés dans l'ectoplasme de la coquille), ainsi que la présence dans la cellule de deux noyaux (végétatif et génératif). De plus, la cavité buccale sur le corps du cilié forme un entonnoir buccal, qui a tendance à passer dans la bouche cellulaire menant au pharynx. C'est là que se créent les vacuoles digestives qui servent directement à la digestion des aliments. Mais les composants non digérés sont éliminés du corps par la poudre. Caractéristiques du type de ciliés très polyvalent, mais les points principaux sont discutés ci-dessus. La seule chose à ajouter est que les deux ciliés sont situés dans des parties opposées du corps. C'est par leur fonctionnement que l'excès d'eau ou de produits métaboliques est excrété par le corps.
Chaussure Infusoire

Afin de considérer qualitativement la structure et le mode de vie d'organismes aussi intéressants d'une structure unicellulaire, il serait conseillé de se référer à l'exemple correspondant. Cela nécessite des chaussures à infusoires, très répandues dans les réservoirs d'eau douce. Ils peuvent facilement être élevés dans des conteneurs ordinaires (par exemple, dans des aquariums), la baie de foin de prairie est la plus facile eau fraiche, car dans les teintures de ce type, en règle générale, un grand nombre d'espèces de protozoaires se développent, y compris des ciliés-chaussures. Ainsi, à travers un microscope, vous pouvez pratiquement étudier toutes les informations fournies dans l'article.
Caractéristiques des ciliés-chaussures
Comme indiqué ci-dessus, les ciliés sont un type qui comprend de nombreux éléments, dont le plus intéressant est la chaussure ciliée. C'est une longueur d'un demi-millimètre, dotée d'une forme de fuseau. Il convient de noter que visuellement cet organisme ressemble à une chaussure, d'où, par conséquent, un nom aussi intrigant. La chaussure d'infusoire est constamment en mouvement et nage avec une extrémité émoussée vers l'avant. Fait intéressant, la vitesse de son mouvement atteint souvent 2,5 mm par seconde, ce qui est très bien pour un représentant de ce type. À la surface du corps des ciliés-chaussures, on peut observer des cils qui servent d'organites moteurs. Comme tous les ciliés, l'organisme en question a deux noyaux dans sa structure : le grand est responsable des processus nutritionnels, respiratoires, moteurs et métaboliques, et le petit participe à l'aspect sexuel.
Le corps des ciliés-chaussures

La structure du corps des ciliés-chaussures est très complexe. Le revêtement extérieur de ce représentant est une fine coque élastique. Il est capable de maintenir la forme correcte du corps du corps tout au long de la vie. Des assistants fidèles dans ce domaine sont des fibres de soutien parfaitement développées situées dans la couche cytoplasmique, qui est étroitement adjacente à la membrane. La surface du corps de la chaussure ciliée est dotée d'un grand nombre (environ 15 000) de cils, qui fluctuent quelles que soient les circonstances extérieures. A la base de chacun d'eux se trouve un corps basal. Les cils bougent environ 30 fois par seconde pour pousser le corps vers l'avant. Il est important de noter que les mouvements ondulatoires de ces instruments sont très coordonnés, ce qui permet aux infusoires de tourner lentement et magnifiquement autour de l'axe longitudinal de son corps en cours de mouvement.
Ciliés - un type d'intérêt défini
Pour une compréhension absolue de toutes les caractéristiques des chaussures ciliées, il est conseillé de considérer les principaux processus de sa vie. Donc, cela revient à manger des bactéries et des algues. Le corps de l'organisme est doté d'un évidement appelé bouche cellulaire et passant dans le pharynx, au fond duquel les aliments pénètrent directement dans la vacuole. Là, il est digéré pendant environ une heure, faisant la transition d'un environnement acide à un environnement alcalin dans le processus. Les vacuoles se déplacent dans le corps du cilié à travers le flux du cytoplasme, et les résidus non digérés sortent à l'arrière du corps à travers la poudre.
La respiration des ciliés-chaussures s'effectue au moyen de l'oxygène pénétrant dans le cytoplasme à travers le tégument du corps. Et les processus excréteurs se produisent à travers deux vacuoles contractiles. Quant à l'irritabilité des organismes, les ciliés-chaussures ont tendance à s'assembler en complexes bactériens en réponse à l'action de substances sécrétées par les bactéries. Et ils flottent loin d'un tel irritant comme le sel de table.
la reproduction

Le cilié de la chaussure peut se reproduire de deux manières. La reproduction asexuée s'est généralisée, selon laquelle les noyaux sont divisés en deux parties. A la suite de cette opération, chaque cilié contient 2 noyaux (grand et petit). La reproduction sexuée est appropriée lorsqu'il y a des carences nutritionnelles ou un changement du régime de température du corps de l'animal. Il convient de noter qu'après cela, les infusoires peuvent se transformer en kyste. Mais avec le type de reproduction sexuelle, une augmentation du nombre d'individus est exclue. Ainsi, deux ciliés sont connectés l'un à l'autre pendant un certain temps, à la suite de quoi la coquille est dissoute et un pont de liaison est formé entre les animaux. Il est important que le gros noyau de chacun d'eux disparaisse sans laisser de trace et que le petit passe deux fois par le processus de fission. Ainsi, dans chaque cilié, 4 noyaux filles sont formés, après quoi trois d'entre eux sont détruits et le quatrième est à nouveau divisé. Ce processus sexuel s'appelle la conjugaison. Et sa durée peut aller jusqu'à 12 heures.
L'infusoire-chaussure vit dans de petits réservoirs stagnants. Cet animal unicellulaire, de 0,5 mm de long, a un corps en forme de fuseau, ressemblant vaguement à une chaussure. Les ciliés sont constamment en mouvement, nageant avec une extrémité émoussée vers l'avant. La vitesse de déplacement de cet animal atteint 2,5 mm par seconde. À la surface du corps, ils ont des organites de mouvement - des cils. Il y a deux noyaux dans la cellule : un gros noyau est responsable de la nutrition, de la respiration, du mouvement, du métabolisme ; le petit noyau est impliqué dans le processus sexuel.
La structure des infusoires de la chaussure
L'organisme des ciliés est plus compliqué. La fine coquille élastique qui recouvre l'extérieur du cilié maintient une forme constante de son corps. Ceci est également facilité par des fibrilles de soutien bien développées, qui sont situées dans la couche de cytoplasme adjacente à la coquille. Environ 15 000 cils oscillants sont situés à la surface du corps du cilié. A la base de chaque cil se trouve un corps basal. Le mouvement de chaque cil consiste en un coup sec dans une direction et un retour plus lent et plus doux à sa position d'origine. Les cils vibrent environ 30 fois par seconde et, comme des rames, poussent les infusoires vers l'avant. Le mouvement ondulatoire des cils est coordonné. Lorsque la chaussure ciliée nage, elle tourne lentement autour de l'axe longitudinal du corps.
Processus vitaux
La nutrition
La pantoufle et certains autres ciliés libres se nourrissent de bactéries et d'algues.
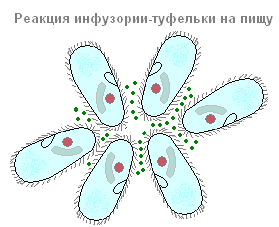
La réaction des ciliés-chaussures à la nourriture
Coquille élastique mince, ( membrane cellulaire) recouvrant le cilié de l'extérieur, conserve une forme corporelle constante. Environ 15 000 cils sont situés à la surface du corps. Sur le corps, il y a un évidement - une bouche cellulaire, qui passe dans un pharynx cellulaire. Au bas du pharynx, les aliments pénètrent dans la vacuole digestive. Dans la vacuole digestive, les aliments sont digérés en une heure, d'abord avec une réaction acide puis alcaline. Les vacuoles digestives se déplacent dans le corps des ciliés par le courant du cytoplasme. Les résidus non digérés sont rejetés à l'extrémité postérieure du corps par une structure spéciale - la poudre, située derrière l'ouverture de la bouche.
Haleine
La respiration se fait par le tégument du corps. L'oxygène pénètre dans le cytoplasme à travers toute la surface du corps et oxyde les substances organiques complexes, à la suite desquelles elles se transforment en eau, en dioxyde de carbone et en certains autres composés. En même temps, de l'énergie est libérée, ce qui est nécessaire à la vie de l'animal. Le dioxyde de carbone est éliminé sur toute la surface du corps lors de la respiration.
Sélection
Dans le corps des chaussures ciliées, il y a deux vacuoles contractiles, situées aux extrémités antérieure et postérieure du corps. Ils recueillent l'eau avec des substances dissoutes formées lors de l'oxydation de substances organiques complexes. Ayant atteint la valeur limite, les vacuoles contractiles se rapprochent de la surface du corps et leur contenu se déverse. Chez les animaux unicellulaires d'eau douce, l'excès d'eau est éliminé par des vacuoles contractiles, qui pénètrent constamment dans leur corps depuis l'environnement.
Irritabilité
Les chaussures ciliées se rassemblent en grappes de bactéries en réponse à l'action des substances qu'elles sécrètent, mais s'éloignent d'un irritant tel que le sel de table.
L'irritabilité est une propriété de tous les organismes vivants pour répondre aux actions des stimuli - lumière, chaleur, humidité, produits chimiques, influences mécaniques. En raison de l'irritabilité, les animaux unicellulaires évitent les conditions défavorables, trouvent de la nourriture, les individus de leur année.
la reproduction
asexué
Les ciliés se reproduisent généralement de manière asexuée en se divisant en deux. Les noyaux sont divisés en deux parties et chaque nouveau cilié contient un grand et un petit noyau. Chacune des deux filles reçoit une partie des organites, tandis que les autres se forment à nouveau.
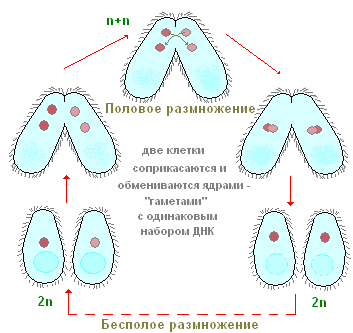
Reproduction de ciliés-chaussures
sexuel
Avec un manque de nourriture ou un changement de température, les ciliés vont à la reproduction sexuée, puis peuvent se transformer en kyste.
Pendant le processus sexuel, une augmentation du nombre d'individus ne se produit pas. Deux ciliés sont temporairement connectés l'un à l'autre. Au point de contact, la coquille se dissout et un pont de liaison se forme entre les animaux. Le gros noyau de chaque cilié disparaît. Le petit noyau se divise deux fois. Dans chaque cilié, quatre noyaux filles sont formés. Trois d'entre eux sont détruits et le quatrième est à nouveau divisé. En conséquence, deux cœurs restent dans chacun. L'échange nucléaire se produit le long du pont cytoplasmique et là, il fusionne avec le noyau restant. Les noyaux nouvellement formés forment un grand et un petit noyau, et les ciliés divergent. Ce processus sexuel s'appelle la conjugaison. Cela dure environ 12 heures. Le processus sexuel conduit au renouvellement, à l'échange entre les individus et à la redistribution du matériel héréditaire (génétique), ce qui augmente la vitalité des organismes.
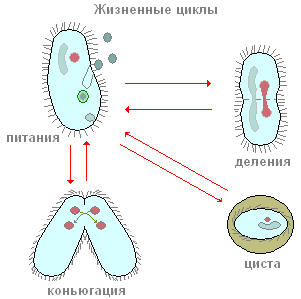
Cycle de vie des ciliés-chaussures
Nous recommandons également
 Héros pionniers de la Grande Guerre patriotique Héros de la présentation des pionniers de la guerre patriotique
Héros pionniers de la Grande Guerre patriotique Héros de la présentation des pionniers de la guerre patriotique
 Présentation "Formation de la posture chez les enfants d'âge préscolaire Hygiène de la présentation de la posture correcte pour les enfants
Présentation "Formation de la posture chez les enfants d'âge préscolaire Hygiène de la présentation de la posture correcte pour les enfants
 Sciences du corps humain
Sciences du corps humain
 Présentation "historique et perspectives du développement de la robotique"
Présentation "historique et perspectives du développement de la robotique"
 La valeur de la lutte de la Russie avec les Polovtsy
La valeur de la lutte de la Russie avec les Polovtsy
 L'Asie et l'Afrique après la Seconde Guerre mondiale
L'Asie et l'Afrique après la Seconde Guerre mondiale