Compiti a casa. Ciabatta Infusori
Rispetto ad altri gruppi di protozoi, i ciliati hanno la struttura più complessa, associata alla diversità e complessità delle loro funzioni.
Da dove viene il nome "pantofola infusoria"? Non sarete sorpresi se osservate al microscopio un ciliato vivo, e nemmeno la sua immagine (Fig. 85).
In effetti, la forma del corpo di questo ciliato ricorda un'elegante scarpa da donna.
La scarpa infusoria è in continuo movimento piuttosto veloce. La sua velocità (a temperatura ambiente) è di circa 2,0-2,5 mm/sec. È un sacco di velocità per un animale così piccolo! Dopotutto, questo significa che in un secondo la scarpa percorre una distanza che supera di 10-15 volte la lunghezza del suo corpo. La traiettoria della scarpa è piuttosto complicata. Muove l'avantreno dritto in avanti
SCARPA INFUSORIUM (PARAMECIUM CAUDATUM)
Per conoscere la struttura e il modo di vivere di questi interessanti organismi unicellulari, passiamo prima a un esempio caratteristico. Prendiamo i ciliati a scarpa (specie del genere Paramecium), che sono diffusi nei bacini d'acqua dolce poco profondi. Questi ciliati sono molto facili da allevare in piccoli acquari, se riempite lo stagno con normale fieno di prato. In tali tinture si sviluppano molti diversi tipi di protozoi e quasi sempre si sviluppano ciliati a scarpa. Con l'aiuto di un normale microscopio didattico, puoi vedere molto di ciò che verrà discusso ulteriormente.
Tra i più semplici scarpe ciliate sono organismi abbastanza grandi. La loro lunghezza del corpo è di circa 1/6-1/3 mm. e ruota a destra lungo l'asse longitudinale del corpo.
Un movimento così attivo della scarpa dipende dal lavoro di un gran numero delle migliori appendici simili a capelli: le ciglia che coprono l'intero corpo del ciliato. Il numero di ciglia in un individuo dei ciliati della scarpa è 10-15 mila!
Ogni cilio fa movimenti simili a una pagaia molto frequenti - a temperatura ambiente fino a 30 battiti al secondo. Durante il contraccolpo, il ciglio viene tenuto in posizione raddrizzata. Quando ritorna alla sua posizione originale (quando si scende), si muove 3-5 volte più lentamente e descrive un semicerchio.
Quando una scarpa nuota, si riassumono i movimenti di numerose ciglia che ricoprono il suo corpo. Le azioni delle singole ciglia sono coordinate, risultando nelle corrette vibrazioni ondulate di tutte le ciglia. L'onda di oscillazione inizia all'estremità anteriore del corpo e si propaga all'indietro. Allo stesso tempo, 2-3 onde di contrazione passano lungo il corpo della scarpa. Pertanto, l'intero apparato ciliare del ciliato è, per così dire, un unico insieme fisiologico funzionale, le azioni delle singole unità strutturali di cui (ciglia) sono strettamente collegate (coordinate) tra loro.
La struttura di ogni singolo ciglio della scarpa, come dimostrato da studi al microscopio elettronico, è molto complessa.
La direzione e la velocità di movimento della scarpa non sono quantità costanti e immutabili. La scarpa, come tutti gli organismi viventi (lo abbiamo già visto con l'esempio dell'ameba), reagisce ad un cambiamento ambiente esterno cambio di direzione del movimento.
Un cambiamento nella direzione del movimento dei protozoi sotto l'influenza di vari stimoli è chiamato taxi. Nei ciliati è facile osservare vari taxi. Se in una goccia dove le scarpe galleggiano, metti una sostanza che le influenzi negativamente (ad esempio un cristallo sale da tavola), quindi le scarpe si allontanano (come se scappassero) da questo fattore per loro sfavorevole (Fig. 86).

Davanti a noi c'è un esempio di taxi negativo su un effetto chimico (chemiotassi negativa). Puoi osservare la scarpa e la chemiotassi positiva. Se, ad esempio, una goccia d'acqua in cui nuotano i ciliati è coperta da un vetrino e viene lasciata una bolla di anidride carbonica (CO2), la maggior parte dei ciliati andrà in questa bolla e si depositerà attorno ad essa in un squillo.
Il fenomeno dei taxi si manifesta molto chiaramente nelle scarpe sotto l'influenza di una corrente elettrica. Se una debole corrente elettrica viene fatta passare attraverso il liquido in cui galleggiano le scarpe, si può osservare la seguente immagine: tutti i ciliati orientano il loro asse longitudinale parallelamente alla linea di corrente, quindi, come a comando, si spostano verso il catodo, in l'area di cui formano un denso ammasso. Il movimento dei ciliati, determinato dalla direzione della corrente elettrica, è chiamato galvanotassi. Vari taxi nei ciliati possono essere rilevati sotto l'influenza di un'ampia varietà di fattori ambientali.
L'intero corpo citoplasmatico del ciliato è nettamente suddiviso in 2 strati: quello esterno è più chiaro (ectoplasma) e quello interno è più scuro e granulare (endoplasma), un ruolo importante nel mantenimento della costanza della forma corporea dei ciliati.
Nello strato esterno (nell'ectoplasma) del corpo di una scarpa vivente sono chiaramente visibili numerose aste corte, poste perpendicolarmente alla superficie (Fig. 85, 7). Queste formazioni sono chiamate tricocisti. La loro funzione è molto interessante ed è connessa alla protezione dei più semplici. Con un'irritazione meccanica, chimica o qualche altra forte irritazione, le tricocisti vengono espulse con forza, trasformandosi in sottili fili lunghi che colpiscono un predatore che attacca la scarpa. Le tricocisti sono una potente difesa. Sono disposti regolarmente tra le ciglia, in modo che il numero di tricocisti corrisponda approssimativamente al numero di ciglia. Al posto delle tricocisti usate ("sparate"), ne si sviluppano di nuove nell'ectoplasma della scarpa.
Da un lato, all'incirca al centro del corpo (Fig. 85, 5), la scarpa presenta una depressione piuttosto profonda. Questa è la cavità orale, o peristoma. Le ciglia si trovano lungo le pareti del peristoma e lungo la superficie del corpo. Sono sviluppati qui molto più potentemente che sul resto della superficie del corpo. Queste ciglia ravvicinate sono disposte in due gruppi. La funzione di queste ciglia altamente differenziate è associata non al movimento, ma alla nutrizione (Fig. 87).

Come e cosa mangiano le scarpe, come si digeriscono?
Le scarpe sono tra i ciliati, il cui alimento principale sono i batteri. Insieme ai batteri, possono anche ingerire qualsiasi altra particella sospesa nell'acqua, indipendentemente dal loro valore nutritivo. Le ciglia periorali creano un flusso continuo d'acqua con particelle sospese nella direzione dell'apertura orale, che si trova in profondità nel peristoma. Piccole particelle di cibo (il più delle volte batteri) penetrano attraverso la bocca in una piccola faringe tubolare e si accumulano sul fondo di essa, al confine con l'endoplasma. L'apertura della bocca è sempre aperta. Forse non sarà un errore dire che la scarpa ciliata è uno degli animali più voraci: si nutre continuamente. Questo processo si interrompe solo in determinati momenti della vita associati alla riproduzione e al processo sessuale.
Il grumo di cibo accumulato sul fondo della faringe si stacca quindi dal fondo della faringe e, insieme a una piccola quantità di liquido, entra nell'endoplasma, formando un vacuolo digestivo. Quest'ultimo non rimane nel sito della sua formazione, ma, cadendo nelle correnti dell'endoplasma, compie un percorso piuttosto complesso e regolare nel corpo della scarpa, chiamato ciclosi del vacuolo digestivo (Fig. 88). Durante questo viaggio piuttosto lungo (a temperatura ambiente, che richiede circa un'ora) del vacuolo digestivo, si verificano al suo interno una serie di cambiamenti associati alla digestione del cibo in esso contenuto.
Qui, come nelle amebe e in alcuni flagellati, si verifica la tipica digestione intracellulare. Dall'endoplasma che circonda il vacuolo digestivo, entrano gli enzimi digestivi che agiscono sulle particelle di cibo. I prodotti della digestione vengono assorbiti attraverso il vacuolo digestivo nell'endoplasma.
Nel corso della ciclosi del vacuolo digestivo, vengono sostituite diverse fasi della digestione. Nei primi istanti dopo la formazione di un vacuolo, il liquido di riempimento differisce poco dal liquido dell'ambiente. Presto gli enzimi digestivi iniziano a entrare nel vacuolo dall'endoplasma e la reazione dell'ambiente al suo interno diventa fortemente acida. Questo è facile da rilevare aggiungendo al cibo un indicatore il cui colore cambia a seconda della reazione (acida, neutra o alcalina) dell'ambiente. In questo ambiente acido avvengono le prime fasi della digestione. Quindi l'immagine cambia e la reazione all'interno dei vacuoli digestivi diventa leggermente alcalina. In queste condizioni, ci sono prossimi passi digestione intracellulare. La fase acida è solitamente più breve della fase alcalina; dura circa 1/6-1/4 dell'intera permanenza del vacuolo digestivo nel corpo del ciliato. Tuttavia, il rapporto tra fasi acide e alcaline può variare entro limiti abbastanza ampi, a seconda della natura dell'alimento.
Il percorso del vacuolo digestivo nell'endoplasma termina con il fatto che si avvicina alla superficie del corpo e attraverso la pellicola il suo contenuto, costituito da residui di cibo liquido e non digerito, viene espulso - si verifica la defecazione. Questo processo, a differenza delle amebe, in cui la defecazione può avvenire ovunque, nelle scarpe, come negli altri ciliati, è strettamente confinato in una specifica area del corpo situata sul lato ventrale (l'addome è convenzionalmente chiamato superficie dell'animale su cui è posto il recesso periorale), approssimativamente a metà strada tra il peristoma e l'estremità posteriore del corpo.
Pertanto, la digestione intracellulare è un processo complesso costituito da diverse fasi che si sostituiscono successivamente l'una con l'altra.

I calcoli mostrano che in circa 30-45 minuti, un volume di fluido pari al volume del corpo ciliato viene escreto dalla scarpa attraverso i vacuoli contrattili. Pertanto, a causa dell'attività dei vacuoli contrattili, viene effettuato un flusso continuo di acqua attraverso il corpo del ciliato, che entra dall'esterno attraverso l'apertura della bocca (insieme ai vacuoli digestivi), nonché osmoticamente direttamente attraverso la pellicola. I vacuoli contrattili svolgono un ruolo importante nella regolazione del flusso d'acqua che passa attraverso il corpo del ciliato e nella regolazione della pressione osmotica. Questo processo procede qui in linea di principio allo stesso modo delle amebe, solo la struttura del vacuolo contrattile è molto più complicata.
Per molti anni, tra gli scienziati che studiano i protozoi, c'è stata una disputa sulla questione se ci siano strutture nel citoplasma associate alla comparsa di un vacuolo contrattile o se si formi ogni volta di nuovo. Su un ciliato vivo non si possono osservare strutture speciali che avrebbero preceduto la sua formazione. Dopo che si verifica la contrazione del vacuolo - sistole, nel citoplasma nel sito dell'ex vacuolo non sono visibili strutture. Quindi riappaiono una bolla trasparente o canali adduttori, che iniziano ad aumentare di dimensioni. Tuttavia, non viene trovata alcuna connessione tra il vacuolo appena emergente e il vacuolo precedentemente esistente. Sembra che non vi sia continuità tra i cicli successivi del vacuolo contrattile e qualsiasi nuovo vacuolo contrattile si formi di nuovo nel citoplasma. Tuttavia, metodi di ricerca speciali hanno dimostrato che in realtà non è così. L'uso della microscopia elettronica, che fornisce un ingrandimento molto elevato (fino a 100mila volte), ha mostrato in modo convincente che il ciliato ha un citoplasma particolarmente differenziato nella zona in cui si formano i vacuoli contrattili, costituiti da un intreccio dei tubi più sottili. Pertanto, si è scoperto che il vacuolo contrattile non si presenta nel citoplasma su " posto vuoto”, ma sulla base del precedente organoide cellulare speciale, la cui funzione è la formazione di un vacuolo contrattile.
Come tutti i protozoi, i ciliati hanno un nucleo cellulare. Tuttavia, in termini di struttura dell'apparato nucleare, i ciliati differiscono nettamente da tutti gli altri gruppi di protozoi.
L'apparato nucleare dei ciliati è caratterizzato dal suo dualismo. Ciò significa che i ciliati hanno due diversi tipi di nuclei: nuclei grandi o macronuclei e nuclei piccoli o micronuclei. Vediamo quale struttura ha l'apparato nucleare nei ciliati della scarpa (Fig. 85).
Al centro del corpo del ciliato (a livello del peristoma) è posto un grande nucleo massiccio di forma ovoidale oa forma di fagiolo. Questo è il macronucleo. Nelle immediate vicinanze vi è un secondo nucleo molte volte più piccolo, di solito abbastanza vicino al macronucleo. Questo è un micronucleo. La differenza tra questi due nuclei non è solo nelle dimensioni, è più significativa, influenzando profondamente la loro struttura.
Il macronucleo, rispetto al micronucleo, è molto più ricco di una speciale sostanza nucleare (cromatina, o più precisamente acido desossiribonucleico, abbreviato DNA), che fa parte dei cromosomi.
Studi recenti hanno dimostrato che il macronucleo ha diverse decine (e in alcuni ciliati e centinaia) di volte un largo numero cromosomi rispetto ai micronuclei. Il macronucleo è un tipo molto particolare di nuclei multicromosomici (poliploidi). Pertanto, la differenza tra micro e macronuclei influisce sulla loro composizione cromosomica, che determina la maggiore o minore ricchezza della loro sostanza nucleare: la cromatina.
In uno dei tipi più comuni di ciliati - scarpe(Paramecium caudatum) - ha un macronucleo (abbreviato Ma) e un micronucleo (abbreviato Mi). Questa struttura dell'apparato nucleare è caratteristica di molti ciliati. Altri potrebbero avere diversi Ma e Mi. Ma una caratteristica di tutti i ciliati è la differenziazione dei nuclei in due gruppi qualitativamente differenti, in Ma e Mi, o, in altre parole, il fenomeno del dualismo nucleare.
Come si riproducono i ciliati? Torniamo come esempio alla scarpa infusoria. Se pianti una singola copia della scarpa in una piccola nave (microacquario), in un giorno ci saranno due e spesso quattro ciliati. Come succede? Dopo un certo periodo di nuoto attivo e alimentazione, il ciliato è leggermente allungato in lunghezza. Quindi, esattamente al centro del corpo, compare una costrizione trasversale sempre più profonda (Fig. 90). Alla fine, i ciliati, per così dire, sono allacciati a metà e da un individuo si ottengono due individui, inizialmente leggermente più piccoli dell'individuo genitore. L'intero processo di fissione dura circa un'ora a temperatura ambiente. Lo studio dei processi interni mostra che anche prima che appaia la costrizione trasversale, inizia il processo di fissione dell'apparato nucleare. Mi viene condiviso per primo e solo dopo è Ma. Non ci fermeremo qui considerazione dettagliata processi di fissione nucleare e notiamo solo che Mi è diviso per mitosi, mentre la divisione di Ma in apparenza ricorda la fissione nucleare diretta - amitosi. Questo processo di riproduzione asessuata dei ciliati della scarpa, come si vede, è simile alla riproduzione asessuata di amebe e flagellati. Al contrario, i ciliati nel processo di riproduzione asessuata si dividono sempre trasversalmente, mentre nei flagellati il piano di divisione è parallelo all'asse longitudinale del corpo.
Durante la divisione avviene una profonda ristrutturazione interna del corpo dei ciliati. Si formano due nuovi peristomi, due faringe e due aperture della bocca. Allo stesso tempo, viene cronometrata la divisione dei nuclei basali delle ciglia, grazie alla quale si formano nuove ciglia. Se il numero di ciglia non aumenta durante la riproduzione, come risultato di ogni divisione, gli individui figli riceverebbero circa la metà del numero di ciglia dell'individuo madre, il che porterebbe alla completa "calvizie" dei ciliati. In realtà questo non accade.

Di tanto in tanto, la maggior parte dei ciliati, comprese le scarpe, hanno una forma speciale ed estremamente peculiare del processo sessuale, che si chiama coniugazione. Non analizzeremo qui in dettaglio tutti i complessi cambiamenti nucleari che accompagnano questo processo, ma noteremo solo i più importanti. La coniugazione procede come segue (Fig. 91), due ciliati si avvicinano, sono strettamente applicati l'uno all'altro dai lati ventrali, e in questa forma nuotano abbastanza a lungo insieme (alla scarpa per circa 12 ore a temperatura ambiente). I coniugati quindi si separano. Cosa succede nel corpo dei ciliati durante la coniugazione? L'essenza di questi processi è la seguente (Fig. 91). Il grande nucleo (macronucleo) collassa e gradualmente si dissolve nel citoplasma. I micronuclei prima si dividono, alcuni dei nuclei formati a seguito della fissione vengono distrutti (vedi Fig. 91). Ciascuno dei coniuganti conserva due nuclei. Uno di questi nuclei rimane al suo posto nell'individuo in cui si è formato (il nucleo stazionario), mentre l'altro si muove attivamente nel partner di coniugazione (il nucleo migrante) e si fonde con il suo nucleo stazionario. Pertanto, in ciascuno dei coniuganti in questa fase c'è un nucleo formato come risultato della fusione dei nuclei stazionari e migratori. Questo nucleo complesso è chiamato synkaryon. La formazione di un synkaryon non è altro che un processo di fecondazione. E negli organismi multicellulari, il momento essenziale della fecondazione è la fusione dei nuclei delle cellule germinali. Nei ciliati non si formano cellule germinali, ci sono solo nuclei sessuali, che si fondono tra loro. Pertanto, si verifica una fertilizzazione incrociata reciproca.
Poco dopo la formazione del synkaryon, i coniuganti si separano. Secondo la struttura del loro apparato nucleare, in questa fase differiscono ancora in modo molto significativo dai soliti cosiddetti ciliati neutri (non coniugati), poiché hanno un solo nucleo ciascuno. In futuro, a causa del synkarion, viene ripristinato il normale apparato nucleare. Il synkaryon è diviso (una o più volte). Parte dei prodotti di questa divisione, attraverso complesse trasformazioni legate all'aumento del numero di cromosomi e all'arricchimento della cromatina, si trasforma in macronuclei. Altri mantengono la struttura caratteristica dei micronuclei. In questo modo viene ripristinato l'apparato nucleare caratteristico e tipico dei ciliati, dopodiché i ciliati iniziano la riproduzione asessuata per fissione.
Pertanto, il processo di coniugazione comprende due momenti biologici essenziali: la fecondazione e il ripristino di un nuovo macronucleo dovuto al synkaryon.
Qual è il significato biologico della coniugazione, che ruolo gioca nella vita dei ciliati? Non possiamo chiamarla riproduzione, perché non c'è aumento del numero di individui. Le domande di cui sopra sono servite da materiale per numerosi studi sperimentali condotti in molti paesi. Il risultato principale di questi studi è il seguente. In primo luogo, la coniugazione, come qualsiasi altro processo sessuale, in cui due principi ereditari (paterno e materno) si uniscono in un organismo, porta ad un aumento della variabilità ereditaria, della diversità ereditaria. Un aumento della variabilità ereditaria aumenta le capacità di adattamento dell'organismo alle condizioni ambientali. Il secondo aspetto biologicamente importante della coniugazione è lo sviluppo di un nuovo macronucleo dovuto ai prodotti di fissione del synkaryon e, allo stesso tempo, la distruzione del vecchio. I dati sperimentali mostrano che è il macronucleo a svolgere un ruolo estremamente importante nella vita dei ciliati. Controlla tutti i principali processi vitali e ne determina il più importante: la formazione (sintesi) di una proteina che costituisce la parte principale del protoplasma di una cellula vivente. Con la riproduzione asessuata prolungata per divisione, si verifica un peculiare processo di "invecchiamento" del macronucleo e allo stesso tempo dell'intera cellula: l'attività del processo metabolico diminuisce, la velocità di divisione diminuisce. Dopo la coniugazione (durante la quale, come abbiamo visto, il vecchio macronucleo viene distrutto), si ripristinano il tasso metabolico e il tasso di divisione. Poiché il processo di fecondazione avviene durante la coniugazione, che nella maggior parte degli altri organismi è associato alla riproduzione e alla comparsa di una nuova generazione, nei ciliati l'individuo formatosi dopo la coniugazione può anche essere considerato come una nuova generazione sessuale, che nasce qui, in quanto erano, a causa del "ringiovanimento" del vecchio.
Sull'esempio delle scarpe ciliate, abbiamo incontrato un tipico rappresentante di un'ampia classe di ciliati. Tuttavia, questa classe è caratterizzata da una straordinaria diversità di specie sia nella struttura che nello stile di vita. Diamo un'occhiata più da vicino ad alcune delle forme più caratteristiche e interessanti.
Nei ciliati, le ciglia della scarpa coprono uniformemente l'intera superficie del corpo. Questa è una caratteristica della struttura (Holotricha). Molti ciliati sono caratterizzati da una diversa natura dello sviluppo della copertura ciliare. Il fatto è che le ciglia dei ciliati sono in grado, se combinate insieme, di formare complessi più complessi. Ad esempio, si osserva spesso che le ciglia situate in una o due file vicine l'una all'altra si uniscono (si attaccano) insieme, formando un piatto che, come le ciglia, è in grado di battere. Tali formazioni contrattili lamellari sono dette membranelle (se sono corte) o membrane (se sono lunghe). In altri casi, le ciglia sono unite insieme, situate in un fascio stretto. Queste formazioni - cirri - assomigliano a una spazzola, i cui singoli peli sono incollati insieme. Varie formazioni ciliare complesse sono caratteristiche di molti ciliati. Molto spesso la copertura ciliare non si sviluppa in modo uniforme, ma solo in alcune parti del corpo.
INFUSORIA trombettista (STENTOR POLIMORFO)
Nelle acque dolci, specie di grandi bei ciliati appartenenti a specie di trombettieri(Stentore). Questo nome è abbastanza coerente con la forma del corpo di questi animali, che assomiglia davvero a una pipa (Fig. 92), ampiamente aperta a un'estremità. Alla prima conoscenza con i trombettisti dal vivo, si può notare una caratteristica che non è caratteristica di una scarpa. Alla minima irritazione, anche meccanica (per esempio, picchiettando con una matita sul vetro, dove c'è una goccia d'acqua con i trombettieri), il loro corpo si contrae bruscamente e molto rapidamente (in una frazione di secondo), assumendo un aspetto quasi regolare forma sferica. Poi, piuttosto lentamente (il tempo si misura in secondi), il trombettista si raddrizza, assumendo la sua forma caratteristica. Questa capacità del trombettista di contrarsi rapidamente è dovuta alla presenza di speciali fibre muscolari poste lungo il corpo e nell'ectoplasma. Pertanto, un sistema muscolare può svilupparsi anche in un organismo unicellulare.

Esistono specie nel genere dei trombettieri, alcune delle quali sono caratterizzate da un colore piuttosto brillante. Molto comune nelle acque dolci trombettista blu(Stentor coeruleus), che è di un blu brillante. Questa colorazione del trombettista è dovuta al fatto che i più piccoli granelli di pigmento blu si trovano nel suo ectoplasma.
Un'altra specie di trombettista (Stentor polymorphus) è spesso di colore verde. Il motivo di questa colorazione è abbastanza diverso. Colore verde a causa del fatto che nell'endoplasma del ciliato vivono e si moltiplicano piccole alghe verdi unicellulari, che conferiscono al corpo del trombettista un colore caratteristico. Stentor polymorphus è un tipico esempio di convivenza reciprocamente vantaggiosa - simbiosi. Il trombettista e le alghe sono in un rapporto simbiotico tra loro: il trombettista protegge le alghe che vivono nel suo corpo e fornisce loro l'anidride carbonica formata a seguito della respirazione; dal canto loro, le alghe forniscono al trombettista ossigeno, rilasciato nel processo di fotosintesi. Apparentemente, parte delle alghe viene digerita dai ciliati, essendo cibo per il trombettista.
I trombettieri nuotano lentamente nell'acqua con l'estremità larga in avanti. Ma possono anche essere temporaneamente attaccati al substrato dall'estremità posteriore stretta del corpo, su cui si forma una piccola ventosa.
Nel corpo del trombettista si può distinguere la regione del tronco che si espande da dietro in avanti e un ampio campo periorale (peristomale) situato quasi perpendicolarmente ad esso. Questo campo ricorda un imbuto piatto asimmetrico, su un bordo del quale c'è una rientranza: una faringe che porta all'endoplasma dei ciliati. Il corpo del trombettista è ricoperto da file longitudinali di ciglia corte. Lungo il bordo del campo peristomale, c'è una zona periorale (adorale) di membranella fortemente sviluppata in un cerchio (Fig. 92). Questa zona è costituita da un gran numero di singole placche ciliate, ciascuna delle quali, a sua volta, è composta da molte ciglia incollate tra loro, disposte in due file ravvicinate.
Nella regione dell'apertura orale, le membrane periorali sono avvolte verso la faringe, formando una spirale sinistrorsa. Il flusso d'acqua, causato dall'oscillazione della membrana periorale, è diretto verso l'apertura della bocca (nella profondità dell'imbuto formato dall'estremità anteriore del corpo). Insieme all'acqua, anche le particelle di cibo sospese nell'acqua entrano nella faringe. Gli oggetti alimentari del trombettista sono più diversi di quelli della scarpetta. Insieme ai batteri, mangia piccoli protozoi (ad esempio flagellati), alghe unicellulari, ecc.
Il trombettista ha un vacuolo contrattile ben sviluppato, che ha una struttura particolare. Il serbatoio centrale si trova nel terzo anteriore del corpo, leggermente al di sotto dell'apertura della bocca. Da esso partono due lunghi canali adduttori. Uno di questi va dal serbatoio all'estremità posteriore del corpo, il secondo si trova nella regione del campo peristomale parallelo alla zona periorale della membranella.
L'infusoria trombettista è un oggetto prediletto per le ricerche sperimentali sulla rigenerazione. Numerosi esperimenti hanno dimostrato l'elevata capacità rigenerativa dei trombettisti. Un ciliato con un bisturi sottile può essere tagliato in molte parti e ognuna di esse attraverso poco tempo(diverse ore, a volte un giorno o più) si trasformerà in un trombettiere proporzionalmente costruito, ma piccolo, che poi, a seguito di un'alimentazione vigorosa, raggiunge le dimensioni tipiche di questa specie. Per completare i processi di recupero, il pezzo rigenerante deve contenere almeno un segmento di un macronucleo perlato.
Il trombettista, come abbiamo visto, ha ciglia diverse: da un lato, sono corte, che coprono l'intero corpo, e dall'altro, c'è una zona quasi orale di membranelle. In accordo con questo tratto caratteristico della struttura, fu denominato il distacco di ciliati, a cui appartiene il trombettista ciliati ciliati(Eterotrico).
INFUSORIA BURSARIA (BURSARIA TRUNCATELLA)
Il secondo rappresentante interessante dei ciliati ciliari si trova spesso nelle acque dolci borsari(Bursaria truncatella, Fig. 93). Questo è un gigante tra i ciliati: le sue dimensioni possono raggiungere i 2 mm, le dimensioni più comuni sono 0,5-1,0 mm. Bursaria è chiaramente visibile ad occhio nudo. Secondo il suo nome, la borsa ha la forma di una borsa, spalancata all'estremità anteriore (bursa è una parola latina, tradotta in russo significa "borsa", "borsa") e leggermente espansa all'estremità posteriore. L'intero corpo del ciliato è ricoperto da file longitudinali di ciglia corte. Il loro battito provoca un movimento in avanti piuttosto lento dell'animale. La borsa di mare nuota come se "rotolasse" da un lato all'altro.

Dall'estremità anteriore nelle profondità del corpo (circa 2/3 della sua lunghezza) sporge una depressione periorale - un peristoma. Sul lato ventrale comunica con l'ambiente esterno attraverso una stretta fenditura; sul lato dorsale la cavità peristoma non comunica con l'ambiente esterno. Se osservi la sezione trasversale del terzo superiore del corpo della borsa (Fig. 93, B), puoi vedere che la cavità del peristoma occupa la maggior parte del corpo, mentre il citoplasma lo circonda sotto forma di un bordo.
All'estremità anteriore del corpo, a sinistra, ha origine una zona di membranella periorale (adorale), molto fortemente sviluppata nella borsaria (Fig. 93, 4). Scende nella profondità della cavità peristoma, svoltando a sinistra. La zona adorale si estende nella parte più profonda del peristoma. Non ci sono altre formazioni ciliare nella cavità peristomale oltre alle membrane periorali, fatta eccezione per la striscia ciliare che corre lungo il lato ventrale della cavità peristomalica (Fig. 93, 10). All'interno parete di fondo la cavità peristomale ha una stretta fessura per quasi tutta la sua lunghezza (Fig. 93, 7), i cui bordi sono solitamente strettamente adiacenti l'uno all'altro. Questa è l'apertura della bocca. I suoi bordi si allontanano solo al momento di mangiare.
I Bursaria non hanno una ristretta specializzazione alimentare, ma sono principalmente predatori. Quando avanzano, incontrano vari piccoli animali. Grazie al lavoro delle membrane della zona quasi orale, la preda viene trascinata con forza nella vasta cavità peristomale, da dove non può più nuotare fuori. Gli oggetti alimentari vengono premuti contro la parete dorsale della cavità peristomale e penetrano nell'endoplasma attraverso la fessura orale in espansione. Le borserie sono molto voraci, possono ingoiare oggetti piuttosto grandi: ad esempio, il loro cibo preferito sono i ciliati delle scarpe. Bursaria è in grado di ingoiare 6-7 scarpe di fila. Di conseguenza, nell'endoplasma della borsa si formano vacuoli digestivi molto grandi.
L'apparato nucleare della borsa è piuttosto complicato. Hanno un lungo macronucleo a forma di salsiccia e un numero elevato (fino a circa 30) di piccoli micronuclei sparsi casualmente nell'endoplasma del ciliato.
Le borserie sono tra le poche specie di ciliati d'acqua dolce privi di un vacuolo contrattile. Non è ancora del tutto chiaro come avvenga l'osmoregolazione in questo grande ciliato. Sotto l'ectoplasma della borsa in diverse parti del corpo, si possono osservare varie forme e dimensioni di bolle fluide - vacuoli, che cambiano il loro volume. Apparentemente questi forma irregolare vacuoli e corrispondono nella loro funzione ai vacuoli contrattili di altri ciliati.

È interessante osservare le fasi successive della riproduzione asessuata di Bursaria. Nelle fasi iniziali della divisione si ha una completa riduzione dell'intera cavità del peristoma e della zona quasi orale della membranella (Fig. 94). Si conserva solo il ciglio esterno. Gli infusori hanno la forma di un uovo. Successivamente, il corpo viene allacciato con una scanalatura trasversale in due metà. In ciascuno dei ciliati figli risultanti, attraverso trasformazioni piuttosto complesse, si sviluppa una tipica zona peristoma e periorale di membranelle. L'intero processo di divisione della borsa procede rapidamente e richiede poco più di un'ora.
È molto facile osservare un altro importante processo vitale in borsa, il cui esordio è associato a condizioni sfavorevoli per i ciliati, il processo di formazione di cisti (cistazione). Questo fenomeno è caratteristico, ad esempio, dell'ameba. Ma si scopre che anche protozoi così organizzati come i ciliati sono in grado di passare in uno stato inattivo. Se la cultura in cui vivono i bursaria non viene alimentata o raffreddata in tempo, l'incistazione di massa inizierà tra poche ore. Questo processo procede come segue. I borseridi, così come prima della divisione, perdono la zona peristoma e periorale delle membrane. Quindi diventano completamente sferici, dopo di che distinguono un doppio guscio di forma caratteristica (Fig. 94, D).
Nello stato delle cisti, la borsa può durare mesi. Quando si verificano condizioni favorevoli, il guscio della cisti esplode, la borsa ne esce, sviluppa un peristoma e passa a una vita attiva.
STYLONICHIA MITILO
Un apparato ciliare molto complesso e variamente differenziato ha ciliati correlati distacco del tratto gastrointestinale(Hypotricha), di cui numerose specie vivono sia in acqua dolce che marina. Uno dei rappresentanti più comuni e frequenti di questo gruppo interessante può essere chiamato stilonichia(Stylonichia mytilus). Questo è un ciliato piuttosto grande (lunghezza fino a 0,3 mm), che vive sul fondo dei bacini d'acqua dolce, su vegetazione acquatica(Fig. 95). A differenza della pantofola, del trombettiere e della borsa, la stilonychia manca di una copertura ciliare continua e l'intero apparato ciliare è rappresentato da un numero limitato di formazioni ciliare rigorosamente definite.

Il corpo di Stilonychia (come la maggior parte degli altri ciliati ventrali) è fortemente appiattito nella direzione dorso-addominale e i suoi lati dorsale e ventrale, le estremità anteriore e posteriore sono chiaramente distinguibili. Il corpo è leggermente allargato anteriormente, ristretto posteriormente. Quando si esamina l'animale dal lato ventrale, si vede chiaramente che nel terzo anteriore a sinistra c'è un'apertura pennata e orale disposta in modo complesso.
Sul lato dorsale, le ciglia si trovano abbastanza raramente, che non sono in grado di battere. Possono piuttosto essere chiamati setole elastiche sottili. Sono immobili e non hanno nulla a che fare con la funzione del movimento. A queste ciglia viene solitamente assegnata una funzione tattile e sensibile.
Tutte le formazioni ciliari associate al movimento e alla cattura del cibo sono concentrate sul lato ventrale dell'animale (Fig. 95). C'è un piccolo numero di spesse formazioni simili a dita situate in diversi gruppi. Questi sono cirri addominali. Ognuno di essi è una complessa formazione ciliare, il risultato di una stretta connessione (che si attaccano insieme) di molte dozzine di singole ciglia. Pertanto, i cirre sono come spazzole, i cui singoli peli sono strettamente uniti e collegati tra loro.
Con l'aiuto dei cirri, l'animale si muove abbastanza rapidamente, "corre" lungo il substrato. Oltre a "strisciare" e "correre" sul substrato, la stilonychia è in grado di produrre salti piuttosto acuti e forti, staccandosi immediatamente dal substrato. Questi movimenti bruschi vengono eseguiti con l'aiuto di due potenti cirri di coda (Fig. 95), che non prendono parte al solito "strisciare".
Due file di cirre marginali (marginali) si trovano lungo il bordo del corpo a destra ea sinistra. Dal margine destro dell'animale corrono lungo tutto il corpo, mentre dal margine sinistro raggiungono solo la regione del peristoma. Queste formazioni ciliari servono a spingere l'animale quando si stacca dal substrato e nuota liberamente nell'acqua.
Vediamo, quindi, che il vario e specializzato apparato ciliare della stilonichia le consente di eseguire movimenti molto diversi, in contrasto, ad esempio, con il semplice scivolamento in acqua, come una scarpa o un trombettista.
Complesso è anche l'apparato ciliare associato alla funzione della nutrizione. Abbiamo già visto che il recesso quasi orale (peristoma), in fondo al quale si trova l'apertura della bocca che porta alla faringe, si trova nella metà anteriore dell'animale a sinistra. Lungo il margine sinistro, a partire dall'estremità molto anteriore del corpo, è presente una zona fortemente sviluppata di membranole periorali (adorali). Con il loro battito dirigono il flusso dell'acqua verso l'apertura della bocca. Inoltre, nella regione del recesso peristomale, ci sono altre tre membrane contrattili (membrane), che si estendono nella faringe con le loro estremità interne, e un certo numero di speciali ciglia periorali (Fig. 95). L'intero complesso apparato serve a catturare e dirigere il cibo nell'apertura della bocca.
Stilonychia è uno dei protozoi con una gamma molto ampia di oggetti alimentari. Può essere giustamente definito un onnivoro. Può mangiare, come una scarpa, batteri. Tra i suoi oggetti alimentari ci sono flagellati, alghe unicellulari (spesso diatomee). Infine, Stilonychia può anche essere un predatore, attaccando altre specie più piccole di ciliati e divorandole.
Stilonychia ha un vacuolo contrattile. È costituito da un serbatoio centrale situato all'estremità posteriore sinistra del peristoma e da un canale adduttore diretto all'indietro.
L'apparato nucleare, come sempre nei ciliati, è costituito da un macronucleo e da un micronucleo.
Il macronucleo è composto da due metà collegate da una sottile costrizione; ci sono due micronuclei, si trovano direttamente vicino a entrambe le metà di Ma.
Stilonichia, in parte borsaria, trombettista: sono tutti ciliati con una vasta gamma di oggetti alimentari. La capacità di assorbire vari alimenti è caratteristica della maggior parte dei ciliati. Tuttavia, tra loro si possono trovare anche specie strettamente specializzate in relazione alla natura del cibo.
PREDATORI INFUSORI
Tra i ciliati ci sono predatori molto "scelti" riguardo alle loro prede. Un buon esempio sono gli infusori. didinia(Didinium nasutum). Il didinio è un ciliato relativamente piccolo, con una lunghezza media di circa 0,1-0,15 mm. L'estremità anteriore è allungata a forma di proboscide, all'estremità della quale è posta l'apertura della bocca. L'apparato ciliare è rappresentato da due corolle di ciglia (Fig. 96). Didinius nuota velocemente nell'acqua, cambiando spesso direzione. L'alimento preferito della didinia sono i ciliati a scarpa. In questo caso, il predatore è più piccolo della sua preda. Didinius penetra nella preda con una proboscide, quindi, allargando gradualmente la bocca aprendosi sempre di più, ingoia la scarpa intera! Nella proboscide c'è un apparato speciale, cosiddetto bastoncino. È costituito da una serie di bastoncini elastici e forti situati nel citoplasma lungo la periferia della proboscide. Si ritiene che questo apparato aumenti la forza delle pareti della proboscide, che non scoppia quando ingoia una preda così enorme rispetto al didinium, come una scarpa. Didinio - esempio ultima risorsa predazione tra i protozoi. Se confrontiamo il didinio che ingoia la sua preda - le scarpe - con la predazione negli animali superiori, allora è difficile trovare esempi simili.

Didinius, ingoiando paramecia, ovviamente, si gonfia molto. Il processo di digestione è molto veloce, a temperatura ambiente ci vogliono solo due ore circa. Quindi i resti non digeriti vengono buttati fuori e il didinium inizia a cacciare un'altra vittima. Studi speciali hanno scoperto che la "dieta" quotidiana della didinia è di 12 scarpe - un appetito davvero colossale! Va tenuto presente che negli intervalli tra le prossime "cacce" la didinia a volte si divide. Con la mancanza di cibo, i didinia si incistano molto facilmente e altrettanto facilmente escono nuovamente dalle cisti.
ciliati erbivori
Molto meno comune della predazione, si trova tra i ciliati "puro vegetarianismo" - mangiare esclusivamente cibo vegetale. Uno dei pochi esempi di ciliati "vegetariani" può essere rappresentato genere passula(Nassula). Il loro oggetto alimentare sono le alghe blu-verdi filamentose (Fig. 97).

Penetrano nell'endoplasma attraverso la bocca, situata sul lato, quindi ruotano i ciliati in una spirale stretta, che viene gradualmente digerita. I pigmenti delle alghe entrano parzialmente nel citoplasma dei ciliati e lo colorano di un colore verde scuro brillante.
SUVOIKA (VORTICELLA NEBULIFERA)
Un gruppo di ciliati interessante e piuttosto ampio in termini di numero di specie sono forme sessili attaccate al substrato, che formano distacco di occhi tondi(Peritrica). I membri più comuni di questo gruppo sono suvoyki(specie del genere Vorticella).
Suvoyki assomigliano a un fiore elegante come una campana o un mughetto, seduto su un lungo stelo, che è attaccato al substrato con la sua estremità. Il suvoyka trascorre la maggior parte della sua vita in uno stato attaccato al substrato.

Considera la struttura del corpo dei ciliati. In specie diverse, le loro dimensioni variano in un intervallo abbastanza ampio (fino a circa 150 micron). Il disco orale (Fig. 98) si trova su una parte anteriore espansa del corpo, che è completamente priva di ciglia. L'apparato ciliare si trova solo lungo il bordo del disco orale (peristomale) (Fig. 98) in un solco speciale, al di fuori del quale si forma un rullo (labbro peristomale). Tre membrane ciliate corrono lungo il bordo del rullo, due delle quali si trovano verticalmente, una (esterna) è orizzontale. Formano un po' più di un giro completo dell'elica. Queste membrane sono in costante movimento tremolante, dirigendo il flusso d'acqua verso l'apertura della bocca. L'apparato orale inizia piuttosto in profondità con un imbuto ai margini del campo peristomale (Fig. 98), nella cui profondità è presente un'apertura orale che conduce a una breve faringe. I suvoy, come le scarpe, si nutrono di batteri. La loro bocca è costantemente aperta e c'è un flusso d'acqua continuo verso la bocca.
Un vacuolo contrattile senza canali adduttori si trova vicino all'apertura della bocca. Il macronucleo ha una forma nastriforme o salsiccia, strettamente adiacente ad esso c'è un unico piccolo micronucleo.
Suvoyka è in grado di accorciare bruscamente il gambo, che in una frazione di secondo viene attorcigliato con un cavatappi. Allo stesso tempo, anche il corpo del ciliato si contrae: il disco peristomale e le membrane vengono tirati verso l'interno e l'intera estremità anteriore si chiude.

La domanda sorge spontanea: poiché i suvoy sono attaccati al substrato, in che modo viene effettuato il loro insediamento nel bacino? Ciò avviene attraverso la formazione di uno stadio fluttuante, il vagabondo. All'estremità posteriore del corpo ciliato compare una corolla di ciglia (Fig. 99). Allo stesso tempo, il disco peristomale si ritrae verso l'interno e il ciliato si separa dal gambo. Il vagabondo risultante è in grado di nuotare per diverse ore. Gli eventi vengono quindi giocati ordine inverso: il ciliato si attacca al substrato con la sua estremità posteriore, il peduncolo cresce, la corolla posteriore delle ciglia si riduce, il disco peristomale si raddrizza all'estremità anteriore e le membrane adorali iniziano a lavorare. La formazione di vagabondi in suvoyka è spesso associata al processo di riproduzione asessuata. L'infusoria sul gambo si divide e uno degli individui figli (e talvolta entrambi) diventa un vagabondo e nuota via.
Molti tipi di suwok sono in grado di incistare in condizioni avverse.
Tra i ciliati sessili appartenenti al gruppo dei ciliati, solo relativamente poche specie, come i suvoe discussi sopra, sono forme viventi solitarie. La maggior parte delle specie qui incluse sono organismi coloniali.
Di solito la colonialità deriva da una riproduzione asessuata o vegetativa non completamente completata. Gli individui formatisi come risultato della riproduzione, in misura maggiore o minore, mantengono una connessione tra loro e formano insieme un'individualità organica di ordine superiore, unendo grandi quantità singoli individui, da cui prende il nome della colonia (abbiamo già incontrato esempi di organismi coloniali in classe dei flagellati.

Le colonie di ciliati tondi ciliati si formano a causa del fatto che gli individui separati non si trasformano in vagabondi, ma si tengono in contatto tra loro con l'aiuto di steli (Fig. 100). In questo caso il fusto principale della colonia, così come i suoi primi rami, non può essere attribuito a nessuno degli individui, ma appartiene all'intera colonia nel suo insieme. A volte la colonia è composta solo da un piccolo numero di individui, mentre in altre specie di ciliati il numero dei singoli individui della colonia può raggiungere diverse centinaia. Tuttavia, la crescita di qualsiasi colonia non è illimitata. Raggiunte le dimensioni caratteristiche di questa specie, la colonia cessa di crescere e gli individui formatisi per divisione sviluppano una corolla di ciglia, diventano vagabondi e nuotano via, dando origine a nuove colonie.
Le colonie di ciliati tondi ciliati sono di due tipi. In alcuni il fusto della colonia è irriducibile: quando irritato, solo i singoli individui della colonia si contraggono, attirando il pennato, ma l'intera colonia nel suo insieme non subisce modifiche (questo tipo di colonia comprende, ad esempio, i generi Epistylis, Opercularia). In altri (ad esempio il genere Carchesium) il peduncolo dell'intera colonia è in grado di contrarsi, poiché il citoplasma attraversa tutti i rami e quindi collega tra loro tutti gli individui della colonia. Quando tali colonie sono irritate, si restringono completamente. L'intera colonia in questo caso reagisce nel suo insieme, come individualità organica.
Tra tutti i ciliati rotondi coloniali, di particolare interesse è, forse, zootamnico(Zoothamnium arbuscula). Le colonie di questo ciliato si distinguono per una particolare regolarità della struttura. Inoltre, qui all'interno della colonia viene delineato un interessante fenomeno biologico di polimorfismo.

La colonia di zootamnia sembra un ombrello. Su uno stelo principale della colonia ci sono rami secondari (Fig. 101). La dimensione di una colonia adulta è di 2-3 mm, quindi sono chiaramente visibili ad occhio nudo. Zootamnii vivono in piccoli stagni Insieme a acqua pulita. Le loro colonie si trovano solitamente su piante sottomarine, il più delle volte su elodea (peste dell'acqua).
Gli steli di una colonia di zootamnia sono contrattili, poiché il citoplasma contrattile attraversa tutti i rami della colonia, ad eccezione della parte basale del peduncolo principale. Con una riduzione che avviene molto rapidamente e bruscamente, l'intera colonia si raccoglie in un grumo.

La zootamnia è caratterizzata da una disposizione rigorosamente regolare dei rami. Uno stelo principale è attaccato al substrato. Nove rami principali della colonia si dipartono dalla sua parte superiore in un piano perpendicolare al gambo, rigorosamente posizionati l'uno rispetto all'altro (Fig. 102, 6). I rami secondari si estendono da questi rami, sui quali siedono i singoli individui della colonia. Ogni ramo secondario può avere fino a 50 ciliati. Il numero totale di individui nella colonia raggiunge 2-3 mila individui.
La maggior parte degli individui della colonia nella loro struttura assomiglia a piccoli suvoik singoli, di dimensioni 40-60 micron. Ma oltre ai piccoli individui, che sono chiamati microzoidi, sulle colonie adulte, all'incirca nel mezzo dei rami principali, si sviluppano individui di tipo e dimensioni completamente diversi (Fig. 102, 5). Si tratta di grandi individui sferici con un diametro di 200-250 micron, che superano il volume del microzoide di cento o più volte in massa. Gli individui di grandi dimensioni sono chiamati macrozoidi.
Nella loro struttura differiscono significativamente dai piccoli individui della colonia. Il loro peristoma non è espresso: è attratto verso l'interno e non funziona. Fin dall'inizio del suo sviluppo da un microzoide, un macrozoo smette di assumere cibo da solo. Manca di vacuoli digestivi. La crescita del macrozoide è apparentemente effettuata a spese di sostanze che entrano attraverso i ponti citoplasmatici collegando tra loro tutti gli individui della colonia. Nella parte del corpo del macrozoo, con cui è attaccato al gambo, c'è un accumulo di grani speciali (granuli), che, come vedremo, svolgono un ruolo significativo nel suo ulteriore destino. Cosa sono questi grandi macrozoidi sferici, qual è il loro ruolo biologico nella vita di una colonia di zootamnia? L'osservazione mostra che i macrozoidi sono futuri vagabondi da cui si sviluppano nuove colonie. Raggiungere limite di dimensione, il macrozoide sviluppa una corolla di ciglia, si separa dalla colonia e nuota via. Allo stesso tempo, la sua forma cambia leggermente, da sferica diventa conica. Dopo qualche tempo, il vagabondo è sempre attaccato al substrato con il lato su cui si trova la granularità. Inizia immediatamente la formazione e la crescita del gambo e i granuli vengono spesi per la costruzione del gambo, che sono localizzati all'estremità posteriore del vagabondo. Man mano che lo stelo cresce, la granulosità scompare. Dopo che il gambo ha raggiunto la lunghezza finale caratteristica della zootamnia, inizia una serie di divisioni rapidamente successive, che portano alla formazione di una colonia. Queste divisioni sono fatte in una sequenza rigorosamente definita (Fig. 102).
Non ci soffermeremo sui dettagli di questo processo. Prestiamo attenzione solo al seguente interessante fenomeno. Durante le prime divisioni dei vagabondi di zootamnia, durante lo sviluppo di una colonia negli individui in formazione, il pennato e la bocca non funzionano. L'alimentazione inizia più tardi, quando la giovane colonia è già composta da 12-16 individui. Pertanto, tutte le prime fasi dello sviluppo di una colonia vengono eseguite esclusivamente a spese di quelle riserve che si sono formate nel corpo del macrozoide durante la sua crescita e sviluppo sulla colonia madre. C'è un'innegabile somiglianza tra lo sviluppo della zootamnia vagabondo e lo sviluppo dell'uovo negli animali multicellulari. Questa somiglianza si esprime nel fatto che lo sviluppo in entrambi i casi viene effettuato a scapito delle riserve precedentemente accumulate, senza la percezione del cibo dall'ambiente esterno.
Quando si studiano i ciliati rotondi sessili, sorge la domanda: come viene eseguita la forma del processo sessuale caratteristico dei ciliati: la coniugazione? Si scopre che in connessione con uno stile di vita sedentario, subisce alcuni cambiamenti significativi. All'inizio del processo sessuale, sulla colonia si formano piccoli vagabondi speciali. Muovendosi attivamente con l'aiuto di una corolla di ciglia, strisciano per qualche tempo lungo la colonia, per poi venire in coniugazione con grandi individui normali sedentari della colonia. Pertanto, qui si verifica la differenziazione dei coniuganti in due gruppi di individui: piccoli, mobili (microconiuganti) e più grandi, immobili (macroconiuganti). Questa differenziazione dei coniuganti in due categorie, di cui una (microconiuganti) è mobile, era un adattamento necessario a uno stile di vita sedentario. Senza questo, ovviamente, il normale corso del processo sessuale (coniugazione) non potrebbe essere assicurato.
CILITARIO SUCCHIANTE (SUCTORIA)
Un gruppo molto particolare in termini di modo in cui mangiano è rappresentato da succhiare i ciliati(Suctoria). Questi organismi, come il suvoyka e altri ciliati ciliati, sono sessili. Il numero di specie appartenenti a questo ordine è misurato da diverse dozzine. La forma del corpo dei ciliati succhiatori è molto varia. Alcune delle loro specie caratteristiche sono mostrate nella Figura 103. Alcune siedono sul substrato su steli più o meno lunghi, altre non hanno steli, in alcune il corpo si ramifica abbastanza forte, ecc. Tuttavia, nonostante la varietà di forme, tutti i ciliati succhiatori sono caratterizzati dalle due seguenti caratteristiche:
1) completa assenza (nelle forme adulte) dell'apparato ciliare,
2) la presenza di speciali appendici - tentacoli che servono a risucchiare la preda.

In diversi tipi di ciliati succhiatori, il numero di tentacoli non è lo stesso. Spesso sono riuniti in gruppi. Ad alto ingrandimento del microscopio, si può notare che all'estremità del tentacolo è presente un piccolo ispessimento a forma di mazza.
Come funzionano i tentacoli? Non è difficile rispondere a questa domanda osservando per qualche tempo i ciliati che succhiano. Se qualche piccolo protozoo (flagellato, infusoria) tocca il tentacolo del suctoria, allora si attaccherà immediatamente ad esso. Tutti i tentativi della vittima di staccarsi sono generalmente vani. Se continui ad osservare la preda attaccata ai tentacoli, puoi vedere che gradualmente inizia a diminuire di dimensioni. Il suo contenuto viene gradualmente "pompato" attraverso i tentacoli nell'endoplasma del ciliato succhiante fino a quando rimane solo una pellicola della vittima, che viene scartata. Pertanto, i tentacoli dei ciliati succhiatori sono completamente unici, in nessun altro posto nel mondo animale ci sono organi per intrappolare e allo stesso tempo succhiare il cibo (Fig. 103).
I ciliati succhiatori sono predatori immobili che non inseguono la preda, ma la catturano istantaneamente, se solo una preda negligente li tocca da sola.

Perché riferiamo questi organismi peculiari alla classe dei ciliati? A prima vista, non hanno nulla a che fare con loro. I seguenti fatti parlano dell'appartenenza di suctoria ai ciliati. In primo luogo, hanno un apparato nucleare tipico dei ciliati, costituito da un macronucleo e un micronucleo. In secondo luogo, durante la riproduzione, sviluppano ciglia che sono assenti negli individui "adulti". La riproduzione asessuale e, allo stesso tempo, il reinsediamento dei ciliati succhiatori avviene mediante la formazione di vagabondi, dotati di numerose corolle anulari di ciglia. La formazione di vagabondi in suctoria può avvenire in diversi modi. A volte si formano a seguito di una divisione (gemma) non del tutto uniforme, in cui ogni rene che si separa verso l'esterno riceve un segmento di macronucleo e un micronucleo (Fig. 104, L). Su un individuo materno possono formarsi più gemme figlie contemporaneamente (Fig. 104, 5). In altre specie (Fig. 104, D, E) si osserva un metodo molto particolare di "gemma interna". Allo stesso tempo, si forma una cavità all'interno del corpo della madre aspirante, in cui si forma il rene vagabondo. Esce attraverso appositi fori, attraverso i quali si “spreme” con nota difficoltà.
Questo sviluppo dell'embrione all'interno del corpo materno, e quindi l'atto della gravidanza, è un'interessante analogia della più semplice con ciò che accade negli organismi multicellulari superiori.
Nelle pagine precedenti sono stati considerati alcuni tipici rappresentanti a vita libera della classe dei ciliati, diversamente adattati alle diverse condizioni ambientali. È interessante affrontare il tema dell'adattamento dei ciliati alle condizioni di vita e, dall'altro, vedere quali sono le caratteristiche caratteristiche comuni ciliati che vivono in determinate condizioni ambientali nettamente definite.
Prendiamo ad esempio due habitat molto diversi: la vita nella composizione del plancton e la vita sul fondo nello spessore della sabbia.
INFUSORIA PLANCTONICA
Un numero abbastanza elevato di specie ciliate si trova sia nel plancton marino che in quello d'acqua dolce.
Le caratteristiche degli adattamenti alla vita nella colonna d'acqua sono particolarmente pronunciate nei radiolari. La principale linea di adattamento allo stile di vita planctonico è ridotta allo sviluppo di tali caratteristiche strutturali che contribuiscono all'impennata dell'organismo nella colonna d'acqua.

Una tipica famiglia planctonica, inoltre, quasi esclusivamente marina di ciliati è tintinnidi(Tintinnidae, Fig. 105, 5). Il numero totale di specie di tintinnidi finora conosciute è di circa 300. Si tratta di forme piccole, caratterizzate dal fatto che il corpo protoplasmatico del ciliato è posto in una casa trasparente, leggera e allo stesso tempo robusta, costituita da materia organica. Un disco sporge dalla casa, portando una corolla di ciglia, che sono in costante movimento tremolante. Nello stato di infusori in bilico nella colonna d'acqua, è principalmente supportato dal costante lavoro attivo dell'apparato ciliare. La casa, ovviamente, svolge la funzione di protezione della parte inferiore del corpo ciliato. Solo 2 specie di tintinnidi vivono in acqua dolce (senza contare 7 specie caratteristiche solo del lago Baikal).
I ciliati d'acqua dolce hanno altri adattamenti alla vita nel plancton. In molti di essi, il citoplasma è molto fortemente vacuolato (Loxodes, Condylostoma, Trachelius), in modo che assomigli alla schiuma. Ciò porta ad una significativa riduzione del peso specifico. Tutti i ciliati elencati, inoltre, hanno una copertura ciliare, grazie alla quale il corpo del ciliato, in termini di peso specifico, è solo leggermente più grande di peso specifico acqua, facilmente mantenibile in uno stato di "impennata". In alcune specie, la forma del corpo contribuisce ad aumentare la superficie specifica e facilita il volo in acqua. Ad esempio, alcuni ciliati planctonici del lago Baikal hanno la forma di un ombrello o di un paracadute (Liliomorpha, Fig. 105, 2). C'è un ciliato succhiante planctonico nel lago Baikal (Mucophrya pelagica, Fig. 105, 4), che differisce nettamente dai suoi parenti sessili. Questa specie è priva di fusto. Il suo corpo protoplasmatico è circondato da un'ampia guaina viscida, un adattamento che porta alla riduzione del peso. Spuntano lunghi tentacoli sottili che, insieme alla loro funzione diretta, probabilmente ne svolgono anche un'altra: un aumento della superficie specifica, che contribuisce a librarsi in acqua.
Infine, è necessario menzionare un'altra, per così dire, forma indiretta di adattamento dei ciliati alla vita nel plancton. Questo è l'attaccamento di piccoli ciliati ad altri organismi che conducono uno stile di vita planctonico. Sì, tra ciliati circonciliati(Peritricha) sono abbastanza numerose le specie che si attaccano ai copepodi planctonici. Questo è uno stile di vita normale e normale per questi tipi di ciliati.
Insieme ai ciliati circonciliari e tra succhiare(Suctoria) ci sono specie che si insediano su organismi planctonici.
CILIZI CHE VIVONO NELLA SABBIA
Spiagge sabbiose e secche rappresentano un habitat estremamente peculiare. Lungo la costa dei mari occupano vasti spazi e sono caratterizzati da una fauna peculiare.
Effettuato per l'anno scorso Numerosi studi in vari paesi hanno dimostrato che lo strato di molte sabbie marine è molto ricco di varie faune microscopiche o microscopiche. Tra le particelle di sabbia ci sono numerosi piccoli e minuscoli spazi pieni d'acqua. Si scopre che questi spazi sono riccamente popolati da organismi appartenenti ai più diversi gruppi del mondo animale. Qui vivono decine di specie di crostacei, anellidi, nematodi, soprattutto numerosi platelminti, alcuni molluschi, celenterati. In gran numero sono presenti anche protozoi, principalmente ciliati. Secondo i dati moderni, la composizione della fauna dei ciliati che abitano lo spessore delle sabbie marine comprende circa 250-300 specie. Se teniamo presente non solo i ciliati, ma anche altri gruppi di organismi che abitano lo spessore della sabbia, il numero totale delle loro specie sarà molto grande. L'intero insieme di animali che abitano lo spessore della sabbia, che vivono negli spazi più piccoli tra i granelli di sabbia, è chiamato fauna psammofila.
La ricchezza e la composizione delle specie della fauna psammofila è determinata da molti fattori. Tra questi, in particolare importanza ha le dimensioni di particelle di sabbia. Le sabbie a grana grossa hanno una fauna povera. Scarsa è anche la fauna delle sabbie limose a grana molto fine (con un diametro delle particelle inferiore a 0,1 mm), dove, ovviamente, gli spazi tra le particelle sono troppo piccoli perché gli animali vi possano abitare. Le sabbie più ricche di vita sono a grana media e fine.
Il secondo fattore che gioca un ruolo importante nello sviluppo della fauna psammofila è la ricchezza di sabbia nei resti organici e nella materia organica in decomposizione (il cosiddetto grado di saprobità). Le sabbie prive di materia organica sono povere di vita. D'altra parte, anche le sabbie sono quasi senza vita e sono molto ricche di materia organica, poiché il decadimento della materia organica porta all'esaurimento dell'ossigeno. Spesso a questo viene aggiunta la fermentazione anaerobica di idrogeno solforato.
La presenza di acido solfidrico libero è un fattore estremamente negativo per lo sviluppo della fauna.
Negli strati superficiali di sabbia si sviluppa talvolta una flora abbastanza ricca di alghe unicellulari (diatomee, peridiniums). Questo è un fattore che favorisce lo sviluppo della fauna psammofila, poiché molti piccoli animali (compresi i ciliati) si nutrono di alghe.
Infine, un fattore che ha un effetto molto negativo sulla fauna psammofila è il surf. Questo è abbastanza comprensibile, dal momento che il surf, lavando gli strati superiori di sabbia, uccide tutti gli esseri viventi qui. La fauna psammofila è la più ricca di calette riparate e ben riscaldate. Il flusso e il riflusso non impediscono lo sviluppo della fauna psammofila. Quando l'acqua esce temporaneamente con la bassa marea, esponendo la sabbia, allora nello spessore della sabbia, negli intervalli tra i granelli di sabbia, rimane, e questo non impedisce l'esistenza degli animali.
Nei ciliati, che fanno parte della fauna psammofila e appartengono a vari gruppi sistematici (ordini, famiglie), nel processo di evoluzione si sviluppano molte caratteristiche comuni, che sono adattamenti alle peculiari condizioni di esistenza tra le particelle di sabbia.

La figura 106 mostra alcune specie della fauna psammofila dei ciliati appartenenti a diversi ordini e famiglie. Ci sono molte somiglianze tra loro. Il corpo della maggior parte di loro è più o meno fortemente allungato in lunghezza, simile a un verme. Questo lo rende facile "spremere" nei fori più piccoli tra i granelli di sabbia. In moltissime specie (Fig. 106) l'allungamento del corpo è combinato con il suo appiattimento. L'apparato ciliare è sempre ben sviluppato, il che consente all'attivo, con una certa forza, di muoversi in spazi ristretti. Abbastanza spesso, le ciglia si sviluppano su un lato del corpo appiattito simile a un verme, il lato opposto è nudo. Questa caratteristica è probabilmente associata alla capacità, pronunciata nella maggior parte delle specie psammofile, di attaccarsi (attaccarsi) al substrato molto strettamente e molto saldamente attraverso l'apparato ciliare (fenomeno chiamato tigmotassi). Questa proprietà consente agli animali di rimanere al loro posto nei casi in cui si formano correnti d'acqua negli angusti spazi in cui vivono. In questo caso è probabilmente più vantaggioso che il lato opposto a quello su cui l'animale si è attaccato al substrato sia liscio.
Cosa mangiano i ciliati psammofili? Una parte significativa della "dieta" in molte specie sono le alghe, in particolare le diatomee. I batteri li servono in misura minore come cibo. Dipende anche in larga misura dal fatto che ci sono pochi batteri nelle sabbie che non sono fortemente inquinate. Infine, soprattutto tra i ciliati psammofili più grandi, esiste un numero considerevole di forme predatorie che si nutrono di altri ciliati appartenenti a specie più piccole. I ciliati psammofili sono distribuiti, a quanto pare, ovunque.
INFUSORIA APOSTOMATI

ciliati spirofria(Spirophrya subparasitica) allo stato incistato si trova spesso seduto su un piccolo gambo su piccoli crostacei marini planctonici (soprattutto spesso su crostacei del genere Idia). Mentre il crostaceo nuota attivamente nell'acqua di mare, la spirofria che si trova su di esso non subisce alcun cambiamento. Per l'ulteriore sviluppo dei ciliati, è necessario che il crostaceo venga mangiato da un polipo idroide marino, cosa che accade abbastanza spesso (Fig. 107). Non appena le cisti di spirofria, insieme al crostaceo, entrano nella cavità digestiva, ne escono subito dei piccoli ciliati, che iniziano a nutrirsi vigorosamente della pappa alimentare derivante dalla digestione del crostaceo ingerito. Entro un'ora, la dimensione degli infusori aumenta di 3-4 volte. Tuttavia, la riproduzione non avviene in questa fase. Davanti a noi c'è una tipica fase di crescita dei ciliati, chiamata trofonte. Dopo qualche tempo, insieme ai residui di cibo non digerito, il trofonte viene lanciato dal polipo nell'acqua di mare. Qui, nuotando attivamente, scende lungo il corpo del polipo fino alla suola, dove si attacca, essendo circondato da una ciste. Questo stadio di un grande ciliato incistato seduto su un polipo è chiamato tomont. Questa è la fase di allevamento. Tomont non si nutre, ma si divide rapidamente più volte di seguito (Fig. 107, 7). Il risultato è un intero gruppo di ciliati molto piccoli. Il loro numero dipende dalla grandezza del tomont, che a sua volta è determinata dalla grandezza del trofonte che gli ha dato origine. I piccoli ciliati formatisi a seguito della divisione del tomont (si chiamano tomiti o vagabondi) rappresentano lo stadio dell'insediamento.
Lasciano la cisti, nuotano velocemente (senza mangiare allo stesso tempo, ma utilizzando le riserve che hanno nel citoplasma). Se sono "abbastanza fortunati" da imbattersi in un copepode, si attaccano immediatamente ad esso e si incistano. Questa è la fase da cui abbiamo iniziato la nostra considerazione del ciclo.
Nella recensione ciclo vitale spirophria richiama l'attenzione sulla netta delimitazione di stadi che hanno un significato biologico diverso. Trophon è la fase di crescita. Cresce solo, accumula una grande quantità di citoplasma e tutti i tipi di sostanze di riserva a causa di un'alimentazione vigorosa e veloce. Il trofonte non è in grado di riprodursi. Il fenomeno inverso si osserva nel tomont: l'incapacità di nutrirsi e una riproduzione rapida e vigorosa. Dopo ogni divisione, non c'è crescita, e quindi la riproduzione del tomont è ridotta a un rapido decadimento in molti vagabondi. Infine, i vagabondi svolgono la loro funzione speciale e unica caratteristica: sono individui - coloni e distributori della specie. Non sono in grado di mangiare o riprodursi.
CICLO DI VITA DI Ichthyophthirius

Entro la fine del periodo di crescita, l'ichthyophthirius raggiunge dimensioni molto grandi rispetto ai vagabondi: 0,5-1 mm di diametro. Al raggiungimento del valore limite, i ciliati escono attivamente dai tessuti del pesce nell'acqua e nuotano lentamente per qualche tempo con l'aiuto dell'apparato ciliare che copre tutto il loro corpo. Presto, un grande ittioftirio si deposita su qualche oggetto sottomarino e secerne una ciste. Immediatamente dopo l'incistura, iniziano le successive divisioni dei ciliati: prima a metà, poi ogni individuo figlia viene nuovamente diviso in due, e così via fino a 10-11 volte. Di conseguenza, all'interno della cisti si formano fino a 2000 piccoli individui quasi arrotondati ricoperti di ciglia. All'interno della cisti, i vagabondi si muovono attivamente. Perforano il guscio e ne escono. I vagabondi che nuotano attivamente infettano i nuovi pesci.
La velocità di divisione dell'ichthyophthirius nelle cisti, così come la velocità della sua crescita nei tessuti dei pesci, dipende in larga misura dalla temperatura. Secondo studi di vari autori si riportano le seguenti cifre: a 26-27°C lo sviluppo dei vagabondi in una ciste dura 10-12 ore, a 15-16°C impiega 28-30 ore, a 4-5 °C dura 6 -7 giorni.
La lotta contro l'ittioftirio presenta notevoli difficoltà. Di primaria importanza qui sono le misure preventive volte a impedire ai vagabondi che nuotano liberamente di penetrare nei tessuti dei pesci. Per fare ciò, è utile eseguire frequenti trapianti di pesci malati in nuovi serbatoi o acquari, per creare condizioni di flusso, particolarmente efficaci nella lotta contro l'ittiofito.
ciliati tricodini

L'intero sistema di adattamento dei tricodini alla vita sulla superficie dell'ospite mira a non staccarsi dal corpo dell'ospite (che equivale quasi sempre alla morte), pur mantenendo la mobilità. Questi dispositivi sono molto perfetti. Il corpo della maggior parte delle tricodine ha la forma di un disco piuttosto piatto, a volte un cappuccio. Il lato rivolto verso il corpo dell'ospite è leggermente concavo, forma una ventosa di attacco. Lungo il bordo esterno della ventosa c'è una corolla di ciglia ben sviluppate, con l'aiuto delle quali il movimento (strisciamento) dei ciliati avviene principalmente sulla superficie del corpo del pesce. Questa corolla corrisponde alla corolla che si trova nei vagabondi ciliati ciliati tondi sessili discussi sopra. Pertanto, la trichodina può essere paragonata a un vagabondo. Sulla superficie addominale (sulla ventosa), i tricodini hanno un apparato di supporto e attaccamento molto complesso, che aiuta a trattenere i ciliati sull'ospite. Senza entrare nei dettagli della sua struttura, notiamo che la sua base è un anello di configurazione complessa, costituito da segmenti separati portanti denti esterni ed interni (Fig. 109, B). Questo anello forma una base elastica e allo stesso tempo robusta della superficie addominale, che funge da ventosa. Diversi tipi di tricodin si differenziano tra loro per il numero di segmenti che formano l'anello e per la configurazione dei ganci esterno ed interno.
Sul lato del corpo della tricodina opposto al disco è presente un apparato pennato e orale. La sua struttura è più o meno tipica di ciliati circonciliati. Le membrane adorabili contorte in senso orario conducono a una rientranza in fondo alla quale si trova la bocca. L'apparato nucleare della tricodina è tipicamente disposto per i ciliati: un macronucleo a forma di nastro e un micronucleo situato accanto ad esso. C'è un vacuolo contrattile.
I tricodini sono ampiamente distribuiti in serbatoi di tutti i tipi. Soprattutto si trovano su avannotti di diverse specie di pesci. Con la riproduzione di massa, vengono applicati i tricodini grande danno pesce, soprattutto se le masse coprono le branchie. Questo interrompe la normale respirazione del pesce.
Per purificare il pesce dal tricodina, si consiglia di fare bagni terapeutici da una soluzione al 2% di cloruro di sodio o una soluzione allo 0,01% di permanganato di potassio (per gli avannotti - per 10-20 minuti).
INFUSORI DEL TRATTO INTESTINALE DEGLI UNGATES
Dalla cicatrice attraverso la rete, il cibo viene ruttato nella cavità orale, dove viene ulteriormente masticato (gomma da masticare). La massa di cibo masticato ingerita nuovamente attraverso un apposito tubo formato dalle pieghe dell'esofago non va più alla cicatrice, ma al libro e da lì all'abomaso, dove viene esposta ai succhi digestivi del ruminante. Nel caglio, in condizioni di reazione acida e presenza di enzimi digestivi, muoiono i ciliati. Arrivando lì con la gomma da masticare, vengono digeriti.
Il numero di protozoi nel rumine (oltre che nella rete) può raggiungere valori colossali. Se prendi una goccia del contenuto della cicatrice e la esamini al microscopio (se riscaldata, poiché i ciliati si fermano a temperatura ambiente), i ciliati letteralmente sciamano nel campo visivo. È difficile anche nella cultura ottenere una tale massa di ciliati. Il numero di ciliati in 1 cm3 del contenuto della cicatrice raggiunge il milione e spesso di più. In termini di intero volume della cicatrice, questo fornisce cifre davvero astronomiche! La ricchezza del contenuto del rumine con i ciliati dipende in larga misura dalla natura del cibo del ruminante. Se il cibo è ricco di fibre e povero di carboidrati e proteine (erba, paglia), allora ci sono relativamente pochi ciliati nel rumine. Quando i carboidrati e le proteine (crusca) vengono aggiunti alla dieta, il numero di ciliati aumenta notevolmente e raggiunge numeri enormi. Va tenuto presente che c'è un deflusso costante di ciliati. Unendosi con la gomma da masticare nell'abomaso, muoiono. Alto livello il numero dei ciliati è mantenuto dalla loro vigorosa riproduzione.
Anche gli ungulati dalle dita dispari (cavallo, asino, zebra) hanno un gran numero di ciliati nel tratto digerente, ma la loro localizzazione nell'ospite è diversa. Gli ungulati dispari non hanno uno stomaco complesso, a causa del quale è assente la possibilità di sviluppare protozoi nelle sezioni anteriori del tubo digerente. Ma negli equidi, il grosso e il cieco sono molto ben sviluppati, che di solito sono ostruiti da masse di cibo e svolgono un ruolo essenziale nella digestione. In questa sezione dell'intestino, così come nel rumine e nella maglia dei ruminanti, si sviluppa una fauna molto ricca di protozoi, principalmente ciliati, la maggior parte dei quali appartiene anche all'ordine degli endodiniomorfi. Tuttavia, in termini di composizione delle specie, la fauna del rumine dei ruminanti e la fauna dell'intestino crasso degli equidi non coincidono.
CILIZI DEL TRATTO INTESTINALE DEI RUMINANTI
Di grande interesse sono i ciliati famiglia ofrioscolecida(Ophryoscolecidae), relativo a ordine endodiniomorfo. Una caratteristica di questo distacco è l'assenza di una copertura ciliare continua. Formazioni ciliari complesse - cirri - si trovano all'estremità anteriore del corpo dei ciliati nella regione dell'apertura della bocca. A questi elementi di base dell'apparato ciliare si possono aggiungere ulteriori gruppi di cirre, localizzate all'estremità anteriore o posteriore del corpo. Il numero totale di specie di ciliati della famiglia ofrioscolecid è di circa 120.

La figura 110 mostra alcuni dei più tipici ofrioscolecidi del rumine dei ruminanti. I ciliati del genere Entodinium (Entodinium, Fig. 110, L) sono disposti in modo molto semplice. All'estremità anteriore del loro corpo c'è una zona periorale di cirri. L'estremità anteriore del corpo, su cui si trova l'apertura della bocca, può essere tirata verso l'interno. Ectoplasma ed endoplasma sono nettamente delimitati. Il tubo anale è ben visibile all'estremità posteriore, che serve a rimuovere i residui di cibo non digerito. Struttura un po' più complessa anoplodinia(Anoplodinio, Fig. 110, B). Hanno due zone dell'apparato ciliare: cirri periorali e cirri dorsali. Entrambi si trovano nella parte anteriore. All'estremità posteriore del corpo della specie mostrata nella figura, ci sono lunghe escrescenze affilate - questo è abbastanza tipico per molte specie di ofrioscolecidi. È stato suggerito che queste escrescenze contribuiscano alla "spinta" dei ciliati tra le particelle vegetali che riempiono la cicatrice.
tipi Genere Eudiplodinia(Eudiplodinium, Fig. 110, B) sono simili a anoplodinia, ma, a differenza di loro, hanno una placca di base scheletrica situata sul bordo destro lungo la faringe. Questa placca scheletrica è costituita da una sostanza simile in natura chimica alla fibra, cioè alla sostanza che compone i gusci delle cellule vegetali.
In genere polyplastron(Polyplastron, Fig. 110, D, E) c'è un'ulteriore complicazione dello scheletro. La struttura di questi ciliati è vicina all'eudiplodinia. Le differenze si riducono principalmente al fatto che invece di una placca scheletrica, questi ciliati ne hanno cinque. Due di loro, il più grande, si trovano sul lato destro e tre, più piccoli, si trovano sul lato sinistro del ciliato. La seconda caratteristica del polyplastron è un aumento del numero di vacuoli contrattili. L'entodinia ha un vacuolo contrattile, l'anoplodinia e l'eudiplodinia hanno due vacuoli contrattili e il poliplastron ne ha circa una dozzina.
In epidinio(Epidinium, Fig. 110), che hanno uno scheletro di carboidrati ben sviluppato situato sul lato destro del corpo, la zona dorsale del cirro si sposta dall'estremità anteriore al lato dorsale. Le spine spesso si sviluppano all'estremità posteriore dei ciliati di questo genere.
La struttura più complessa rivela genere Ofrioscolex(Ophryoscolex), da cui prende il nome l'intera famiglia dei ciliati (Fig. 110, E). Hanno una zona dorsale del cirro ben sviluppata, che copre circa 2/3 della circonferenza del corpo e delle placche scheletriche. All'estremità posteriore si formano numerose spine, di cui una di solito è particolarmente lunga.
Conoscenza di alcuni rappresentanti tipici ofrioscolecid mostra che all'interno di questa famiglia si è verificata una significativa complicazione dell'organizzazione (dall'entodinia all'ofrioscolex).
Oltre ai ciliati famiglia ofrioscolecida, nel rumine dei ruminanti, si trovano in piccole quantità rappresentanti del già noto a noi. distacco dei ciliati isociliari. Sono rappresentati da un piccolo numero di specie. Il loro corpo è uniformemente coperto da file longitudinali di ciglia, gli elementi scheletrici sono assenti. Nella massa totale della popolazione ciliata del rumine, non svolgono un ruolo significativo, e quindi non ci soffermeremo su di loro qui.
Cosa e come mangiano i ciliati dei rioscolecidi? Questo problema è stato studiato in dettaglio da molti scienziati, in particolare dal professor V. A. Dogel.

Il cibo ofrioscolecid è piuttosto vario e si osserva una certa specializzazione in specie diverse. Le specie più piccole del genere Entodynia si nutrono di batteri, granuli di amido, funghi e altre piccole particelle. Moltissimi ofrioscolecidi medi e grandi assorbono particelle di tessuti vegetali, che costituiscono la maggior parte del contenuto del rumine. L'endoplasma di alcune specie è letteralmente intasato di particelle vegetali. Puoi vedere come i ciliati si avventano su frammenti di tessuti vegetali, li fanno letteralmente a pezzi e poi li inghiottono, spesso attorcigliandoli a spirale nel loro corpo (Fig. 111, 4). A volte è necessario osservare tali immagini (Fig. 111, 2), quando il corpo del ciliato stesso è deformato a causa di grandi particelle ingerite.
Nell'ofrioscolecid si osserva talvolta la predazione. Le specie più grandi divorano quelle più piccole. La predazione (Fig. 112) è combinata con la capacità della stessa specie di nutrirsi di particelle vegetali.

In che modo i ciliati entrano nel rumine di un ruminante? Quali sono le vie di infezione da ofrioscolecides? Si scopre che i ruminanti appena nati non hanno ancora ciliati nel rumine. Sono assenti anche mentre l'animale si nutre di latte. Ma non appena il ruminante passa al cibo vegetale, i ciliati compaiono immediatamente nella cicatrice e nella rete, il cui numero sta crescendo rapidamente. Da dove vengono? Per molto tempo si è ritenuto che il rumine ciliato formasse una sorta di stadi di riposo (molto probabilmente cisti), che sono ampiamente dispersi in natura e, quando ingeriti, danno origine a stadi attivi di ciliati. Ulteriori studi hanno dimostrato che i ciliati dei ruminanti non hanno fasi di riposo. È stato possibile dimostrare che l'infezione si verifica con ciliati mobili attivi che penetrano nella cavità orale durante l'eruttazione. Se si esamina al microscopio preso da cavità orale gomma da masticare, ma ha sempre un gran numero di ciliati attivamente fluttuanti. Queste forme attive possono facilmente penetrare nella bocca e ulteriormente nel rumine di altri ruminanti da un comune vaso da bere, insieme a erba, fieno (che può contenere saliva con ciliati), ecc. Questa via di infezione è stata dimostrata sperimentalmente.
Se non ci sono fasi di riposo negli ofrioscolecidi, è ovviamente facile ottenere animali "senza infuso" isolandoli quando si stanno ancora nutrendo di latte. Se non è consentito il contatto diretto tra giovani in crescita e ruminanti con ciliati, i giovani animali possono essere lasciati senza ciliati nel rumine. Tali esperimenti sono stati condotti da diversi scienziati in diversi paesi. Il risultato è stato chiaro. In assenza di contatto tra animali giovani (latte prelevato dalla madre durante il periodo dell'alimentazione) e ruminanti con ciliati nel rumine, gli animali crescono sterili rispetto ai ciliati. Tuttavia, anche il contatto a breve termine con animali con ciliati (una comune mangiatoia, un comune secchio per abbeverarsi, un comune pascolo) è sufficiente affinché i ciliati appaiano nel rumine degli animali sterili.
Sopra c'erano i risultati di esperimenti sul contenuto dei ruminanti, completamente privi di ciliati nel rumine e nella rete. Ciò si ottiene, come abbiamo visto, mediante l'isolamento precoce dei giovani. Sono stati condotti esperimenti su pecore e capre.
In questo modo è stato possibile condurre osservazioni di animali "infusorless" per un periodo di tempo significativo (oltre un anno). In che modo l'assenza di ciliati nel rumine influisce sulla vita dell'ospite? L'assenza di ciliati influisce negativamente o positivamente sull'ospite? Per rispondere a questa domanda, sono stati effettuati i seguenti esperimenti sulle capre. Gemelli (della stessa cucciolata e dello stesso sesso) sono stati presi per avere più materiale simile. Quindi uno dei gemelli di questa coppia è stato allevato senza ciliati nel rumine (isolamento precoce), mentre l'altro fin dall'inizio dell'alimentazione di cibi vegetali è stato abbondantemente infettato da molti tipi di ciliati. Entrambi hanno ricevuto esattamente la stessa dieta e sono stati allevati nelle stesse condizioni. L'unica differenza tra loro era la presenza o l'assenza di ciliati. Su diverse coppie di bambini studiate in questo modo, non sono state riscontrate differenze nel corso dello sviluppo di entrambi i membri di ciascuna coppia ("infusore" e "non infusore"). Pertanto, si può sostenere che i ciliati che vivono nel rumine e nella rete non hanno alcun effetto netto sulle funzioni vitali dell'animale ospite.
I suddetti risultati degli esperimenti non consentono, tuttavia, di affermare che i ciliati del rumine siano del tutto indifferenti al proprietario. Questi esperimenti sono stati effettuati con una dieta normale dell'ospite. È possibile che in altre condizioni, con una dieta diversa (ad esempio con alimentazione insufficiente), sia possibile rivelare l'effetto sull'ospite della fauna dei protozoi che abitano il rumine.
In letteratura sono stati formulati vari suggerimenti sulla possibile influenza positiva della fauna protozoica del rumine sui processi digestivi dell'ospite. È stato sottolineato che molti milioni di ciliati, nuotando attivamente nel rumine e schiacciando i tessuti vegetali, contribuiscono alla fermentazione e alla digestione delle masse alimentari situate nelle sezioni anteriori del tubo digerente. Un numero significativo di ciliati, che entrano nell'abomaso insieme alla gomma da masticare, viene digerito e la proteina, che costituisce una parte significativa del corpo dei ciliati, viene assorbita. Gli infusori, quindi, possono essere un'ulteriore fonte di proteine per l'ospite. È stato anche suggerito che i ciliati contribuiscano alla digestione delle fibre, che costituiscono la maggior parte del cibo dei ruminanti, e al loro trasferimento a uno stato più digeribile.
Tutte queste ipotesi non sono provate e alcune di esse sono state sollevate obiezioni. È stato fatto notare, ad esempio, che i ciliati costruiscono il protoplasma del loro corpo dalle proteine che entrano nel rumine con il cibo dell'ospite. Assorbendo le proteine vegetali, le convertono nella proteina animale del loro corpo, che viene poi digerita nell'abomaso. Non è chiaro se ciò offra vantaggi all'host. Tutti questi problemi sono di grande interesse pratico, dal momento che stiamo parlando della digestione dei ruminanti, i principali oggetti della zootecnia. Sono altamente auspicabili ulteriori ricerche sul ruolo dei ciliati ruminali nella digestione dei ruminanti.
Gli ofrioscolecidi dei ruminanti hanno, di regola, un'ampia specificità. In termini di specie, la popolazione della cicatrice e della rete di bovini, ovini e caprini è molto vicina tra loro. Se confrontiamo la composizione delle specie del rumine delle antilopi africane con il bestiame, anche qui sarà comune circa il 40% del numero totale di specie. Tuttavia, ci sono molte specie di ofrioscolecid che si trovano solo nelle antilopi o solo nei cervi. Pertanto, sullo sfondo dell'ampia specificità generale dei rioscolecidi, si può parlare di tipi separati e più strettamente specifici di essi.
Infusori degli intestini degli equidi
Passiamo ora a una breve conoscenza dei ciliati che abitano il grande e cieco degli equidi.
In termini di specie, anche questa fauna, come la fauna del rumine dei ruminanti, è molto diversificata. Attualmente sono state descritte circa 100 specie di ciliati che vivono nell'intestino crasso degli animali della famiglia degli equini. I ciliati che si trovano qui nel senso della loro appartenenza a diversi gruppi sistematici sono più diversi dei ciliati del rumine dei ruminanti.

Nell'intestino dei cavalli sono presenti alcune specie di ciliati appartenenti all'ordine degli isociliari, cioè ciliati in cui l'apparato ciliare non forma membranelle o cirrosi vicino alla zona orale (Fig. 113, 1).
Ordine entodiniomorfo(Entodiniomorpha) è anche riccamente rappresentato negli intestini del cavallo. Mentre nel rumine dei ruminanti si trova una sola famiglia di endodiniomorfi (famiglia ofrioscolecid), nell'intestino del cavallo vivono rappresentanti di tre famiglie, su cui però non ci soffermeremo, limitandoci a pochi disegni di specie equine tipiche ( Fig. 113) .
Studi dettagliati di A. Strelkov hanno mostrato che diversi tipi di ciliati sono tutt'altro che distribuiti uniformemente lungo l'intestino crasso del cavallo. Ci sono due diversi gruppi di specie, due faune, per così dire. Uno di essi abita il cieco e la parte addominale del colon crasso (le sezioni iniziali dell'intestino crasso), e l'altro abita la parte dorsale del colon grosso e del colon piccolo. Questi due complessi di specie sono piuttosto nettamente delimitati. Ci sono poche specie comuni a queste due sezioni - meno di una dozzina.
, 
È interessante notare che tra le numerose specie di ciliati che abitano l'intestino crasso degli equidi, ci sono rappresentanti di un genere legato ai ciliati succhiatori. Come abbiamo visto sopra, succhiare i ciliati(Suctoria) sono tipici organismi sessili a vita libera con un modo molto speciale di alimentarsi con l'aiuto dei tentacoli (Fig. 103). Uno di aspirazione del parto adattato a un habitat apparentemente insolito come l'intestino crasso di un cavallo, ad esempio diverse specie allantosi(Allantosoma). Questi animali molto particolari (Fig. 114) non hanno un gambo, le ciglia sono assenti, i tentacoli a forma di clava ispessiti alle estremità sono ben sviluppati.
Con l'aiuto dei tentacoli, gli allantosomi si attaccano a diversi tipi di ciliati e li succhiano. Spesso la preda è molte volte più grande del predatore.
La questione della natura della relazione tra i ciliati dell'intestino crasso degli equidi e i loro ospiti non è ancora chiara. Il numero di ciliati può essere tanto elevato, e talvolta anche superiore, che nel rumine dei ruminanti. Ci sono dati che mostrano che la quantità di ciliati nell'intestino crasso di un cavallo può raggiungere i 3 milioni in 1 cm3. Il significato simbiotico suggerito da alcuni scienziati è persino meno probabile che per i ciliati del rumine.
L'opinione più probabile è che causino qualche danno all'ospite, assorbendo una quantità significativa di cibo. Parte dei ciliati viene estratta con masse fecali, e quindi le sostanze organiche (comprese le proteine) che compongono il loro corpo rimangono inutilizzate dall'ospite.
La questione delle modalità di infezione degli equidi con i ciliati che abitano l'intestino crasso non è stata ancora risolta.

Balantidia cattura una varietà di particelle di cibo dal contenuto dell'intestino crasso. Soprattutto volentieri, si nutre di chicchi di amido. Se balantidia vive nel lume del colon umano, si nutre del contenuto dell'intestino e non ha alcun effetto dannoso. Questa è una tipica "carrozza", che abbiamo già incontrato considerando l'ameba della dissenteria. Tuttavia, è meno probabile che balantidia dell'ameba dissenterica rimanga un "inquilino innocuo".
Al momento, gli specialisti sono ben sviluppati vari metodi, consentendo la coltivazione di balantidia in un ambiente artificiale - esterno all'organismo ospite.

Come si può vedere dalla figura, la troglodptella è uno degli endodiniomorfi complessi. Lei, oltre alla zona periorale della cirrosi (all'estremità anteriore del corpo), ha altre tre zone di cirri ben sviluppati, che coprono anulare il corpo del ciliato. I trogloditelli hanno un apparato scheletrico ben sviluppato costituito da carboidrati, che copre quasi l'intera estremità anteriore del corpo. Le dimensioni di questi ciliati peculiari sono piuttosto significative. In lunghezza, raggiungono i 200-280 micron.
Ciliati senza bocca ASTOMATS

Le formazioni scheletriche di supporto si sviluppano principalmente all'estremità anteriore del corpo, che deve subire stress meccanico e superare ostacoli, spingendo attraverso il lume intestinale tra le particelle di cibo. Specie genere radiofria(Radiophrya) all'estremità anteriore su un lato del corpo (che è convenzionalmente considerato il lato addominale) ci sono costole elastiche molto forti (spicole) che giacciono nello strato superficiale dell'ectoplasma (Fig. 117, B, D, E) . Specie Genere Menilella(Mesnilella) sono presenti anche raggi di supporto (spicole), che per la maggior parte della loro lunghezza giacciono negli strati più profondi del citoplasma (nell'endoplasma, Fig. 117, A). Formazioni di supporto disposte in modo simile si sviluppano anche in specie di altri generi Astomat.
La riproduzione asessuata in alcuni ciliati astomatici procede in modo peculiare. Invece della divisione trasversale in due, caratteristica della maggior parte dei ciliati, molti astomati hanno una divisione irregolare (gemma). Allo stesso tempo, i reni che si separano all'estremità posteriore rimangono per qualche tempo associati all'individuo madre (Fig. 117, B). Di conseguenza, si ottengono catene, costituite da individui grandi anteriori e posteriori più piccoli (reni). In futuro, i reni si separano gradualmente dalla catena e passano a un'esistenza indipendente. Questa peculiare forma di riproduzione è diffusa, ad esempio, nella radiofria a noi già nota. Le catene di alcuni astomi risultanti dal germogliamento somigliano aspetto esteriore catene di tenie. Anche qui incontriamo il fenomeno della convergenza.
L'apparato nucleare dell'astoma ha una struttura caratteristica dei ciliati: un macronucleo, il più delle volte a forma di nastro (Fig. 117), e un micronucleo. I vacuoli contrattili sono generalmente ben sviluppati. La maggior parte delle specie ha diversi (a volte più di una dozzina) vacuoli contrattili disposti in una fila longitudinale.
Uno studio sulla distribuzione delle specie Astomat per diversi tipi di ospiti mostra che la maggior parte delle specie Astomat è confinata a specie ospiti rigorosamente definite. La maggior parte degli astomat sono caratterizzati da una specificità ristretta: solo una specie di animale può fungere da ospite per loro.
Nonostante il gran numero di studi dedicati allo studio dei ciliati astomatici, un aspetto molto importante della loro biologia rimane del tutto oscuro: come avviene la trasmissione di questi ciliati da un individuo ospite all'altro? Non è mai stato possibile osservare la formazione di cisti in questi ciliati.
Pertanto, si suggerisce che l'infezione si verifichi attivamente - fasi mobili.
INFUSORI DELL'INTESTINO DI RICCI DI MARE
I ricci di mare sono molto numerosi nella zona costiera dei nostri mari del nord (Barents) e dell'Estremo Oriente (il Mar del Giappone, la costa del Pacifico delle Isole Curili). Maggioranza ricci di mare si nutre di alimenti vegetali, principalmente alghe, che raschia via oggetti sottomarini con speciali "denti" affilati che circondano l'apertura della bocca. Negli intestini di questi ricci erbivori è presente una ricca fauna di ciliati. Spesso si sviluppano qui in quantità massicce e il contenuto dell'intestino del riccio di mare al microscopio è "brulicante" di infusori quasi quanto il contenuto del rumine dei ruminanti. C'è da dire che, oltre a profonde differenze nelle condizioni di vita dei ciliati degli intestini del riccio di mare e del rumine del ruminante, ci sono alcune somiglianze. Stanno nel fatto che sia qua che là i ciliati vivono in un ambiente molto ricco di resti vegetali. Attualmente, oltre 50 specie di ciliati vivono nell'intestino dei ricci di mare, che si trovano solo nella zona costiera, dove i ricci si nutrono di alghe. A grandi profondità, dove le alghe non crescono più, non ci sono ciliati nei ricci di mare.

Secondo lo stile di vita e la natura dell'alimentazione, la maggior parte dei ciliati dell'intestino dei ricci di mare sono erbivori. Si nutrono di alghe, che in grandi quantità riempiono l'intestino dell'ospite. Alcune specie sono piuttosto "pignolo" nella scelta del cibo. Per esempio, strobilidio(Strobilidium, Fig. 118, A) si nutre quasi esclusivamente di grandi diatomee. Ci sono anche predatori che mangiano rappresentanti di altri, di più piccole specie ciliati.
Nei ciliati dell'intestino dei ricci di mare, a differenza dell'astomat, non esiste un rigoroso confinamento a determinati tipi di ospiti. Vivono in un'ampia varietà di ricci di mare che si nutrono di alghe.
Le modalità di infezione dei ricci di mare con i ciliati non sono state studiate. Tuttavia, qui con un alto grado di probabilità si può presumere che si verifichi in forme attive fluttuanti. Il fatto è che i ciliati dell'intestino dei ricci di mare possono vivere a lungo (molte ore) nell'acqua di mare. Tuttavia, si sono già adattati così tanto alla vita nell'intestino dei ricci che fuori dal loro corpo, nell'acqua di mare, prima o poi muoiono.
Terminata la conoscenza dei ciliati, va sottolineato ancora una volta che rappresentano un gruppo (classe) ricco di specie, un gruppo (classe) esteso e prospero del mondo animale. Rimanendo a livello di organizzazione cellulare, gli infusori hanno raggiunto, rispetto ad altre classi di protozoi, la massima complessità strutturale e funzionale.
Un ruolo particolarmente significativo in questo progressivo sviluppo (evoluzione) è stato probabilmente svolto dalla trasformazione dell'apparato nucleare e dall'emergere del dualismo nucleare (non equivalenza qualitativa dei nuclei). La ricchezza del macronucleo in sostanze nucleiche è associata a processi metabolici attivi, a vigorosi processi di sintesi di proteine nel citoplasma e nei nuclei.
Conclusione
Siamo giunti alla fine della nostra rassegna della struttura e dello stile di vita di un vasto tipo di mondo animale - protozoi. La loro caratteristica, come è stato più volte sottolineato sopra, è l'unicellularità. In termini di struttura, i protozoi sono cellule. Tuttavia, sono incomparabili con le cellule che compongono il corpo degli organismi multicellulari, perché essi stessi sono organismi. Pertanto, i protozoi sono organismi a livello di organizzazione cellulare. Alcuni protozoi altamente organizzati, che possiedono molti nuclei, sembrano già andare oltre i limiti morfologici della struttura cellulare, il che dà ragione ad alcuni scienziati di chiamare tali protozoi "sopracellulari". Questo cambia poco l'essenza della questione, perché un'organizzazione unicellulare è ancora tipica dei protozoi.
Entro i limiti dell'unicellularità, i protozoi hanno percorso una lunga strada di sviluppo evolutivo e hanno dato un'enorme varietà di forme adattate alle più diverse condizioni di vita. Nel cuore del tronco genealogico dei protozoi ci sono due classi: sarcodi e flagellati. La questione di quale di queste classi sia più primitiva è ancora oggetto di dibattito nella scienza. Da un lato, i rappresentanti inferiori dei Sarcodidae (ameba) hanno la struttura più primitiva. Ma i flagellati mostrano la massima plasticità del tipo di metabolismo e si trovano, per così dire, al confine tra il mondo animale e quello vegetale. Nel ciclo vitale di alcuni sarcodi (ad esempio i foraminiferi) sono presenti stadi flagellari (gameti), che indicano la loro relazione con i flagellati. È ovvio che né i moderni sarcodi né i moderni flagellati possono essere il gruppo iniziale dell'evoluzione del mondo animale, perché essi stessi hanno fatto molta strada nello sviluppo storico e hanno sviluppato numerosi adattamenti alle moderne condizioni di vita sulla Terra. Probabilmente, entrambe queste classi di protozoi moderni dovrebbero essere considerate come due tronchi in evoluzione, originati da forme antiche che non sono sopravvissute fino ad oggi, vissute agli albori dello sviluppo della vita sul nostro pianeta.
Nell'ulteriore evoluzione dei protozoi si sono verificati cambiamenti di natura diversa. Alcuni di loro hanno portato a un aumento generale del livello di organizzazione, un aumento dell'attività e dell'intensità dei processi vitali. Tra tali trasformazioni filogenetiche (evolutive), si dovrebbe, ad esempio, includere lo sviluppo degli organelli di movimento e la cattura del cibo, che ha raggiunto un alto livello di perfezione nella classe dei ciliati. È indiscutibile che le ciglia sono organelli corrispondenti (omologhi) ai flagelli. Mentre nei flagellati, con poche eccezioni, il numero dei flagelli è piccolo, nei ciliati il numero delle ciglia ha raggiunto molte migliaia. Lo sviluppo dell'apparato ciliare aumentò notevolmente l'attività dei protozoi, rese più diverse e complesse le forme del loro rapporto con l'ambiente, le forme di reazione agli stimoli esterni. La presenza di un apparato ciliare differenziato, indubbiamente, fu una delle ragioni principali della progressiva evoluzione nella classe dei ciliati, dove sorse un'ampia varietà di forme adattate ai diversi habitat.
Lo sviluppo dell'apparato ciliare dei ciliati è un esempio di tali cambiamenti evolutivi, che sono stati nominati da Acad. Severtsov aromorfosi. Le aromorfosi sono caratterizzate da un generale aumento dell'organizzazione, dallo sviluppo di adattamenti di ampio significato. Un aumento dell'organizzazione è inteso come cambiamenti che causano un aumento dell'attività vitale dell'organismo; sono associati alla differenziazione funzionale delle sue parti, portando a forme più diverse di comunicazione tra l'organismo e l'ambiente. Lo sviluppo dell'apparato ciliare dei ciliati si riferisce proprio a questo tipo di trasformazioni strutturali nel processo evolutivo. Questa è una tipica aromorfosi.
Nei protozoi, come sottolineato da V. A. Dogel, i cambiamenti nel tipo di aromorfosi sono solitamente associati ad un aumento del numero di organelli. Si verifica la polimerizzazione degli organelli. Lo sviluppo dell'apparato ciliare nei ciliati è un tipico esempio di tali cambiamenti. Il secondo esempio di aromorfosi nell'evoluzione dei ciliati è il loro apparato nucleare. Abbiamo esaminato sopra le caratteristiche strutturali del nucleo dei ciliati. Il dualismo nucleare dei ciliati (la presenza di un micronucleo e di un macronucleo) è stato accompagnato da un aumento del numero di cromosomi nel macronucleo (fenomeno della poliploidia). Poiché i cromosomi sono associati ai principali processi sintetici nella cellula, principalmente alla sintesi delle proteine, questo processo ha portato ad un aumento generale dell'intensità delle principali funzioni vitali. Anche qui avveniva la polimerizzazione, che interessava i complessi cromosomici del nucleo.
ciliati- uno dei gruppi di protozoi più numerosi e progressivi, deriva dai flagellati. Ciò è evidenziato dalla completa somiglianza morfologica dei loro organoidi di movimento. Questa fase dell'evoluzione era associata a due grandi aromorfosi: una di esse interessava gli organelli del movimento, la seconda - l'apparato nucleare. Entrambi questi tipi di cambiamenti sono interconnessi, poiché entrambi portano ad un aumento dell'attività vitale e alla complicazione di forme di interrelazione con l'ambiente esterno.
Insieme alle aromorfosi, c'è un altro tipo di cambiamento evolutivo, espresso nello sviluppo di adattamenti (adattamento) a determinate condizioni di esistenza ben definite. Questo tipo di cambiamento evolutivo è stato chiamato idioadattamento da Severtsov. Nell'evoluzione dei protozoi, questo tipo di cambiamento ha giocato un ruolo molto importante. Sopra, quando si considera classi diverse protozoi, vengono forniti numerosi esempi di cambiamenti idioadattivi. Adattamenti a uno stile di vita planctonico in diversi gruppi di protozoi, adattamenti alla vita nella sabbia nei ciliati, formazione di gusci protettivi di oocisti nei coccidi e molto altro: tutti questi sono idioadattamenti che hanno giocato un ruolo importante nell'emergenza e nello sviluppo di singoli gruppi, ma non sono associati a cambiamenti progressivi generali nell'organizzazione.
Gli adattamenti a vari habitat specifici nei protozoi sono molto diversi. Hanno assicurato l'ampia distribuzione di questo tipo in un'ampia varietà di habitat, di cui si è discusso in dettaglio sopra, quando si descrivono le singole classi.
Grande enciclopedia medica
Infusoria Tetrahymena thermophila ... Wikipedia
- (Infusoria) una classe dei protozoi più sviluppati (Protozoa). Le caratteristiche principali di I.: la presenza di ciglia (per il movimento e la nutrizione), due tipi di nuclei (macronucleo poliploide e micronucleo diploide, diversi per struttura e ... ... Grande enciclopedia sovietica
Oppure animali liquidi (Infusoria) una classe di animali protozoi, dotati di trecce o ciglia luccicanti, talvolta sostituite in età adulta da escrescenze cilindriche succhianti, con una certa forma corporea, per la maggior parte con ... ... dizionario enciclopedico F. Brockhaus e I.A. Efron
Tassonomia dell'ifusoria Classificazione e sistematizzazione degli organismi appartenenti alla tipologia dei Ciliati, secondo il concetto generale di tassonomia dei protozoi in particolare, e degli animali in generale Contenuti 1 Sistematica 1.1 Classica ... Wikipedia
Oppure animali liquidi (Infusoria) una classe di animali protozoi, dotati di trecce o ciglia luccicanti, che nello stato adulto sono talvolta sostituite da escrescenze cilindriche succhianti, con una certa forma corporea, per la maggior parte con ... ... Enciclopedia di Brockhaus e Efron
Il tipo di ciliati è anche comunemente chiamato ciliato - gli organi di movimento di questi protozoi sono ciglia. La cellula ciliata ha due nuclei, sono chiamati piccoli e grandi. Il primo regola il processo di riproduzione e il secondo è responsabile dei processi di nutrizione, movimento e respirazione.
Le caratteristiche della vita di questo tipo dovrebbero essere considerate sull'esempio delle scarpe ciliate.
Movimento e respiro
La scarpa infusoria, la cui lunghezza è di circa 0,5 mm, sceglie i corpi idrici come suo habitat. La forma del corpo dei più semplici è facile da indovinare dal nome: ricorda una scarpa. La velocità di movimento è di circa 2,5 mm al secondo.
La presenza di un guscio elastico esterno fornisce una forma del corpo stabile.
Nel citoplasma, che è adiacente al guscio, ci sono fibre di supporto, il loro sviluppo è una garanzia della conservazione della forma costante del ciliato.
Sulla superficie dei ciliati ci sono 15 mila ciglia, alla loro base c'è un corpo basale. Il movimento avviene con l'aiuto delle vibrazioni delle ciglia: producono circa 30 colpi al secondo, spingendo così in avanti la scarpa ciliata.
Respira attraverso la superficie del corpo.
Cibo
Una caratteristica dei ciliati è la presenza bocca cellulare, vicino al quale ci sono ciglia particolarmente lunghe e dense. La bocca cellulare continua con la faringe cellulare: le ciglia spingono l'acqua e ciliano il cibo - i batteri al suo interno.
L'infusoria rileva le sostanze chimiche, che rilasciano un gruppo di batteri. Quindi, lei cerca la preda.
Il cibo finisce poi nel vacuolo digestivo dove viene digerito. Da qui segue già nel citoplasma.
Selezione
La selezione viene effettuata utilizzando due vacuoli contrattili, uno si trova nella parte anteriore e l'altro nella parte posteriore. I vacuoli sono costituiti da un serbatoio e canali.
Il liquido riempie i canali, quindi segue il serbatoio centrale e quindi esce dagli infusori. Il processo di contrazione dei vacuoli dura 10-20 secondi.
riproduzione
L'infusoria si riproduce in modo asessuato: è diviso in due. La sua caratteristica è la divisione in tutto il corpo.
I nuclei dei ciliati sono divisi in due parti: i ciliati di nuova formazione hanno un nucleo piccolo e uno grande. I ciliati figli possiedono parti di organoidi e quelli mancanti si formano indipendentemente. La riproduzione avviene più volte al giorno.
Per le scarpe-ciliati è possibile anche la riproduzione sessuata, ma in questo caso non c'è aumento del numero di individui. Temporaneamente, i protozoi sono collegati, formando un ponte di collegamento dal citoplasma.
In ogni individuo, il nucleo grande scompare e quelli piccoli vengono divisi due volte: compaiono quattro nuclei. Di questi rimane un solo nucleo, che pure si divide. Un individuo contiene due nuclei, quindi si verifica uno scambio di nuclei: uno dei nuclei si sposta su un altro individuo.
Lì si fonde con il nucleo che rimane, e così si forma un nucleo piccolo e più grande in ciascuno degli individui. Questo processo, chiamato coniugazione, è necessario per il rinnovo del materiale genetico tra individui.
Tipi di ciliati
I ciliati sono protozoi organizzati in modo complesso, ce ne sono circa 7.000 specie.
1. Varietà di sarcodi.
2. Varietà di flagelli.
I rappresentanti del secondo sottotipo si muovono con l'aiuto dei flagelli, che si riflette nel nome di questo gruppo.
- Flagellati. Euglena green è un tipico rappresentante di questo gruppo. Questo protozoo ha una forma fusiforme, ricoperto da una densa membrana elastica. All'estremità anteriore del corpo c'è un flagello, vicino alla base del quale c'è un occhio sensibile alla luce: uno stigma e un vacuolo contrattile. Più vicino all'estremità posteriore, nello spessore del citoplasma, c'è un nucleo che controlla tutti i processi vitali del corpo. Il colore verde di Euglena è dovuto a numerosi cloroplasti. Alla luce, questo flagellato si fotosintetizza, ma trovandosi per lungo tempo in zone non illuminate del bacino, passa a nutrirsi dei prodotti di decomposizione di sostanze organiche complesse, estraendole dall'ambiente. Pertanto, l'euglena verde può combinare sia un tipo di metabolismo autotrofico che eterotrofico. L'esistenza di organismi con un tipo di alimentazione così misto indica la relazione tra il mondo animale e quello vegetale. La maggior parte dei flagelli
3. Varietà di sporozoi e ciliati.
Il tipo Ciliates comprende i protozoi, che si distinguono per l'organizzazione più complessa tra gli animali unicellulari. Un tipico rappresentante di questo gruppo è la scarpa infusoria, un comune abitante dell'acqua dolce. Il suo corpo è ricoperto da un denso guscio - pellicola e quindi ha una forma relativamente costante. Una caratteristica della struttura è la presenza di ciglia, che ricoprono uniformemente l'intero corpo della scarpa. I movimenti delle ciglia sono coordinati grazie alla rete di fibre contrattili situata nello strato superficiale del citoplasma. La seconda caratteristica è la presenza di due nuclei, grande (macronucleo) e piccolo (micronucleo). Anche i nuclei differiscono funzionalmente: uno grande regola il metabolismo e uno piccolo partecipa al processo sessuale (coniugazione). La scarpa infusoria si nutre di batteri, alghe unicellulari, che vengono adattate dalle ciglia alla bocca cellulare, che si trova sul fondo della cavità preorale. Dopo aver attraversato la faringe cellulare, che termina nel citoplasma, le particelle di cibo sono racchiuse in vacuoli digestivi, dove vengono scomposte dall'azione degli enzimi. I residui non digeriti vengono espulsi attraverso la polvere. Nel corpo della scarpa sono presenti due vacuoli contrattili complessi che pulsano alternativamente. Le scarpe si riproducono asessualmente, dividendo a metà. La riproduzione asessuale multipla è sostituita nella scarpa dal processo sessuale - coniugazione, durante il quale due scarpe si uniscono e si scambiano materiale genetico. Successivamente, le scarpe divergono e presto ricominciano la riproduzione asessuata. Il significato biologico della coniugazione risiede nella combinazione in un organismo delle proprietà ereditarie di due individui. Ciò ne aumenta la fattibilità, che si esprime in una migliore adattabilità alle condizioni ambientali.
Non hai trovato quello che stavi cercando? Usa la ricerca
In questa pagina, materiale sugli argomenti:
- brevemente su flagellati, pseudopodi, sporozoi e ciliati
- sarcodi e sporozoi
- varietà di flagelli
- riassunto del rapporto della pantofola infusoria
- qual è la differenza tra le specie ciliate e sarcode
Uno dei più noti rappresentanti del ciliare è la scarpa ciliata. Vive, di regola, in acque con una direzione stagnante, così come in bacini d'acqua dolce, dove la corrente si distingue per l'eccezione dell'assertività. Il suo habitat deve necessariamente contenere materia organica in decomposizione. Sarebbe opportuno considerare in dettaglio tutti gli aspetti della vita di questo rappresentante della fauna.
Rappresentanti delle ciglia

Va notato che i ciliati sono un tipo il cui nome deriva dalla parola "tintura" (tradotto da latino). Ciò può essere spiegato dal fatto che i primi rappresentanti dei protozoi sono stati trovati proprio nelle tinture a base di erbe. Nel tempo, lo sviluppo di questo tipo ha iniziato a prendere rapidamente slancio. Pertanto, già oggi in biologia sono note circa 6-7 mila specie, che include il tipo di Ciliati. Se ci basiamo sui dati degli anni '80, allora si può sostenere che il tipo in questione contiene due classi nella sua struttura: Ciliated ciliates (ha tre superordini) e Sucking ciliates. In connessione con queste informazioni, possiamo concludere che la diversità degli organismi viventi è molto ampia, il che è di vero interesse.
Tipo di infusori: rappresentanti

Rappresentanti di spicco di questo tipo sono ciliati-balantidi e ciliati-scarpa. Caratteristiche distintive Questi animali sono la copertura della pellicola con le ciglia, che servono per il movimento, la protezione dei ciliati attraverso organi appositamente progettati, le tricocisti (situate nell'ectoplasma della conchiglia), nonché la presenza nella cellula di due nuclei (vegetativi e generativa). Inoltre, la cavità orale sul corpo del ciliato forma un imbuto orale, che tende a passare nella bocca cellulare che porta alla faringe. È lì che si creano i vacuoli digestivi, che servono direttamente alla digestione del cibo. Ma i componenti non digeriti vengono rimossi dal corpo attraverso la polvere. Caratteristiche del tipo di ciliati molto versatile, ma i punti principali sono discussi sopra. L'unica cosa da aggiungere è che i due ciliati si trovano in parti opposte del corpo. È attraverso il loro funzionamento che l'acqua in eccesso oi prodotti metabolici vengono espulsi dal corpo.
Scarpa per infusori

Per considerare qualitativamente la struttura e lo stile di vita di organismi così interessanti di struttura unicellulare, sarebbe opportuno fare riferimento all'esempio corrispondente. Ciò richiede scarpe-infusori, diffuse nei bacini d'acqua dolce. Possono essere facilmente allevati in normali contenitori (ad esempio negli acquari), la baia di fieno di prato è la più semplice acqua dolce, perché in tinture di questo tipo, di regola, si sviluppano moltissime specie di protozoi, comprese le scarpe ciliate. Quindi, attraverso un microscopio, puoi praticamente studiare tutte le informazioni fornite nell'articolo.
Caratteristiche dei ciliati-scarpe
Come notato sopra, i ciliati sono un tipo che include molti elementi, il più interessante dei quali è la scarpa ciliata. Questa è la cui lunghezza è di mezzo millimetro, dotata di una forma a fuso. Va notato che visivamente questo organismo assomiglia a una scarpa, quindi, di conseguenza, un nome così intrigante. La scarpa per infusori è costantemente in uno stato di movimento e nuota con un'estremità smussata in avanti. È interessante notare che la velocità del suo movimento raggiunge spesso i 2,5 mm al secondo, il che è molto buono per un rappresentante di questo tipo. Sulla superficie del corpo delle scarpe ciliate si possono osservare ciglia che fungono da organelli motori. Come tutti i ciliati, l'organismo in questione ha due nuclei nella sua struttura: quello grande è responsabile dei processi nutritivi, respiratori, motori e metabolici, e quello piccolo partecipa all'aspetto sessuale.
Il corpo dei ciliati-scarpe

La struttura del corpo delle scarpe ciliate è molto complessa. Il rivestimento esterno di questo rappresentante è un sottile guscio elastico. È in grado di mantenere la forma corretta del corpo del corpo per tutta la vita. I fedeli assistenti in questo sono fibre di supporto perfettamente sviluppate situate nello strato citoplasmatico, che è strettamente adiacente alla membrana. La superficie del corpo della scarpa ciliata è dotata di un numero enorme (circa 15.000) di ciglia, che oscillano indipendentemente dalle circostanze esterne. Alla base di ciascuno di essi c'è un corpo basale. Le ciglia si muovono circa 30 volte al secondo per spingere il corpo in avanti. È importante notare che i movimenti ondulatori di questi strumenti sono molto coordinati, il che consente agli infusori di ruotare lentamente e magnificamente attorno all'asse longitudinale del suo corpo durante il movimento.
Infusoria - un tipo sicuramente di interesse
Per una comprensione assoluta di tutte le caratteristiche delle scarpe-ciliati, è opportuno considerare i principali processi della sua vita. Quindi, si tratta di mangiare batteri e alghe. Il corpo dell'organismo è dotato di una cavità chiamata bocca cellulare e passa nella faringe, in fondo alla quale il cibo entra direttamente nel vacuolo. Lì viene digerito per circa un'ora, facendo il passaggio da un ambiente acido a uno alcalino nel processo. I vacuoli si muovono nel corpo del ciliato attraverso il flusso del citoplasma e i residui non digeriti escono nella parte posteriore del corpo attraverso la polvere.
La respirazione delle scarpe ciliate viene effettuata mediante l'ingresso di ossigeno nel citoplasma attraverso il tegumento del corpo. E i processi escretori avvengono attraverso due vacuoli contrattili. Per quanto riguarda l'irritabilità degli organismi, i ciliati-scarpe tendono ad assemblarsi in complessi batterici in risposta all'azione di sostanze secrete dai batteri. E volano via da un irritante come il sale da cucina.
riproduzione

Il ciliato della scarpa può riprodursi in due modi. La riproduzione asessuale è diventata più diffusa, secondo la quale i nuclei sono divisi in due parti. Come risultato di questa operazione, ogni ciliato contiene 2 nuclei (grande e piccolo). La riproduzione sessuale è appropriata quando ci sono alcune carenze nutrizionali o un cambiamento nel regime di temperatura del corpo dell'animale. Va notato che dopo questo, gli infusori possono trasformarsi in una cisti. Ma con il tipo di riproduzione sessuale si esclude un aumento del numero di individui. Quindi, due ciliati sono collegati tra loro per un certo periodo di tempo, a seguito del quale il guscio si dissolve e si forma un ponte di collegamento tra gli animali. È importante che il grande nucleo di ciascuno di essi scompaia senza lasciare traccia e quello piccolo subisca il processo di fissione due volte. Pertanto, in ogni ciliato si formano 4 nuclei figli, dopo di che tre di essi vengono distrutti e il quarto viene nuovamente diviso. Questo processo sessuale è chiamato coniugazione. E la sua durata può arrivare fino a 12 ore.
La scarpa infusoria vive in piccoli serbatoi stagnanti. Questo animale unicellulare, lungo 0,5 mm, ha un corpo a forma di fuso, vagamente simile a una scarpa. I ciliati sono costantemente in movimento, nuotando con l'estremità smussata in avanti. La velocità di movimento di questo animale raggiunge i 2,5 mm al secondo. Sulla superficie del corpo hanno organelli di movimento: le ciglia. Ci sono due nuclei nella cellula: un grande nucleo è responsabile della nutrizione, della respirazione, del movimento, del metabolismo; il piccolo nucleo è coinvolto nel processo sessuale.
La struttura degli infusori della scarpa
L'organismo dei ciliati è più complicato. Il sottile guscio elastico che ricopre l'esterno del ciliato mantiene una forma costante del suo corpo. Ciò è facilitato anche da fibrille di supporto ben sviluppate, che si trovano nello strato di citoplasma adiacente al guscio. Circa 15.000 ciglia oscillanti si trovano sulla superficie del corpo del ciliato. Alla base di ogni ciglio si trova un corpo basale. Il movimento di ciascuna ciglia consiste in un colpo netto in una direzione e in un ritorno più lento e regolare alla sua posizione originale. Le ciglia vibrano circa 30 volte al secondo e, come i remi, spingono in avanti gli infusori. Il movimento ondulatorio delle ciglia è coordinato. Quando la scarpa ciliata nuota, ruota lentamente attorno all'asse longitudinale del corpo.
Processi vitali
Cibo
La pantofola e alcuni altri ciliati a vita libera si nutrono di batteri e alghe.
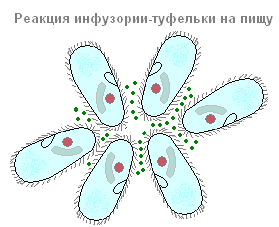
La reazione dei ciliati-scarpe al cibo
Guscio elastico sottile, ( membrana cellulare) coprendo il ciliato dall'esterno, mantiene una forma corporea costante. Circa 15 mila ciglia si trovano sulla superficie del corpo. Sul corpo c'è una rientranza: una bocca cellulare, che passa in una faringe cellulare. Nella parte inferiore della faringe, il cibo entra nel vacuolo digestivo. Nel vacuolo digestivo, il cibo viene digerito entro un'ora, prima con una reazione acida e poi con una alcalina. I vacuoli digestivi si muovono nel corpo dei ciliati dalla corrente del citoplasma. I residui non digeriti vengono espulsi all'estremità posteriore del corpo attraverso una struttura speciale - polvere, situata dietro l'apertura della bocca.
Respiro
La respirazione avviene attraverso il tegumento del corpo. L'ossigeno entra nel citoplasma attraverso l'intera superficie del corpo e ossida sostanze organiche complesse, a seguito delle quali si trasformano in acqua, anidride carbonica e alcuni altri composti. Allo stesso tempo, viene rilasciata energia, necessaria per la vita dell'animale. L'anidride carbonica viene rimossa attraverso l'intera superficie del corpo durante la respirazione.
Selezione
Nel corpo delle scarpe ciliate ci sono due vacuoli contrattili, che si trovano alle estremità anteriore e posteriore del corpo. Raccolgono l'acqua con le sostanze disciolte formate durante l'ossidazione di sostanze organiche complesse. Raggiunto il valore limite, i vacuoli contrattili si avvicinano alla superficie del corpo e il loro contenuto fuoriesce. Negli animali unicellulari d'acqua dolce, l'acqua in eccesso viene rimossa attraverso i vacuoli contrattili, che penetrano costantemente nel loro corpo dall'ambiente.
Irritabilità
Le scarpe ciliate si riuniscono in grappoli di batteri in risposta all'azione delle sostanze che secernono, ma nuotano lontano da un irritante come il sale da cucina.
L'irritabilità è una proprietà di tutti gli organismi viventi di rispondere alle azioni degli stimoli: luce, calore, umidità, sostanze chimiche, influenze meccaniche. Grazie all'irritabilità, gli animali unicellulari evitano condizioni avverse, trovano cibo, individui del loro anno.
riproduzione
asessuale
I ciliati di solito si riproducono asessualmente dividendo in due. I nuclei sono divisi in due parti e ogni nuovo ciliato contiene un nucleo grande e uno piccolo. Ognuna delle due figlie riceve parte degli organelli, mentre le altre si formano nuovamente.
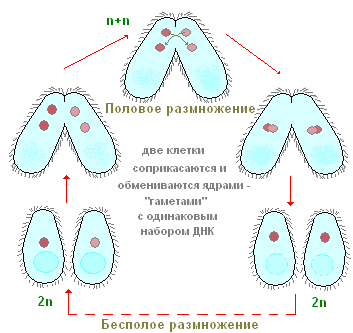
Riproduzione di ciliati-scarpe
sessuale
Con una mancanza di cibo o un cambiamento di temperatura, i ciliati vanno alla riproduzione sessuale e quindi possono trasformarsi in una cisti.
Durante il processo sessuale, non si verifica un aumento del numero di individui. Due ciliati sono temporaneamente collegati tra loro. Nel punto di contatto, il guscio si dissolve e si forma un ponte di collegamento tra gli animali. Il grande nucleo di ogni ciliato scompare. Il piccolo nucleo si divide due volte. In ogni ciliato si formano quattro nuclei figli. Tre di loro vengono distrutti e il quarto viene nuovamente diviso. Di conseguenza, in ciascuno rimangono due core. Lo scambio nucleare avviene lungo il ponte citoplasmatico e lì si fonde con il nucleo rimanente. I nuclei appena formati formano un nucleo grande e uno piccolo e i ciliati divergono. Questo processo sessuale è chiamato coniugazione. Dura circa 12 ore. Il processo sessuale porta al rinnovamento, allo scambio tra individui e alla ridistribuzione del materiale ereditario (genetico), che aumenta la vitalità degli organismi.
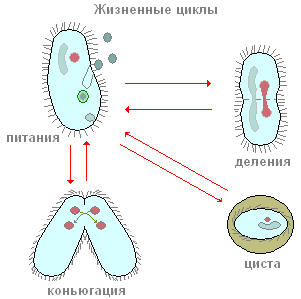
Ciclo di vita dei ciliati-scarpe
Consigliamo anche
 La guerra più lunga nella storia dell'umanità: storia, fatti interessanti Teorie sull'origine delle guerre
La guerra più lunga nella storia dell'umanità: storia, fatti interessanti Teorie sull'origine delle guerre
 Caramelle da masticare ROSHEN Ape pazza
Caramelle da masticare ROSHEN Ape pazza
 Picodi: Tutti gli sconti in un unico posto!
Picodi: Tutti gli sconti in un unico posto!
 Cosa è importante che un contabile sappia su una valutazione speciale dei lavori?
Cosa è importante che un contabile sappia su una valutazione speciale dei lavori?
 P \ 'piani quinquennali in URSS Tutti i piani quinquennali dell'URSS
P \ 'piani quinquennali in URSS Tutti i piani quinquennali dell'URSS
 Statistiche spaventose dall'assedio di Leningrado
Statistiche spaventose dall'assedio di Leningrado