Ո՞րն է տառադարձման գործընթացը: ՌՆԹ-ն ստանում է ժառանգական տեղեկատվություն
Տառադարձման նախաձեռնում
Տառադարձման երկարացում
Այն պահը, երբ ՌՆԹ պոլիմերազն անցնում է տրանսկրիպցիայի մեկնարկից դեպի երկարացում, ճշգրիտ որոշված չէ: Երեք հիմնական կենսաքիմիական իրադարձությունները բնութագրում են այս անցումը Escherichia coli ՌՆԹ պոլիմերազի դեպքում. պոլիմերազ, ներառում է աճող ՌՆԹ շղթան և տառադարձված ԴՆԹ-ն։ Նույն երեւույթները բնորոշ են նաեւ էուկարիոտ ՌՆԹ պոլիմերազներին։ Անցումը մեկնարկից դեպի երկարացում ուղեկցվում է ֆերմենտի, պրոմոտորի, տրանսկրիպցիան մեկնարկող գործոնների միջև կապերի խզմամբ, իսկ որոշ դեպքերում՝ ՌՆԹ պոլիմերազի անցումով երկարացման կոմպետենտության վիճակի (օրինակ՝ CTD տիրույթի ֆոսֆորիլացում ՌՆԹ պոլիմերազ II): Երկարացման փուլն ավարտվում է աճող տառադարձման թողարկումից հետո, և ֆերմենտը տարանջատվում է կաղապարից (վերջացում):
Երկարացումն իրականացվում է հիմնական երկարացման գործոնների օգնությամբ, որոնք անհրաժեշտ են, որպեսզի գործընթացը ժամանակից շուտ չդադարի։
Վերջերս ի հայտ եկան ապացույցներ, որոնք ցույց են տալիս, որ կարգավորիչ գործոնները կարող են նաև կարգավորել երկարացումը: Երկարացման գործընթացում ՌՆԹ պոլիմերազը կանգ է առնում գենի որոշ հատվածներում։ Սա հատկապես հստակ երևում է սուբստրատների ցածր կոնցենտրացիաների դեպքում: Մատրիցայի որոշ հատվածներում նկատվում են ՌՆԹ պոլիմերազի առաջխաղացման երկար ձգձգումներ, այսպես կոչված: դադարներ են նկատվում նույնիսկ սուբստրատի օպտիմալ կոնցենտրացիաների դեպքում: Այս դադարների տեւողությունը կարելի է վերահսկել երկարացման գործոններով։
Ավարտ
Բակտերիաներն ունեն տառադարձման դադարեցման երկու մեխանիզմ.
- rho-կախյալ մեխանիզմ, որի դեպքում Rho (rho) սպիտակուցը ապակայունացնում է ջրածնային կապերը ԴՆԹ-ի կաղապարի և mRNA-ի միջև՝ ազատելով ՌՆԹ-ի մոլեկուլը:
- rho-independent, որի դեպքում տրանսկրիպցիան դադարում է, երբ նոր սինթեզված ՌՆԹ մոլեկուլը ձևավորում է ցողունային հանգույց, որին հաջորդում են մի քանի ուրացիլներ (...UUUU), ինչը հանգեցնում է ՌՆԹ-ի մոլեկուլի անջատմանը ԴՆԹ-ի կաղապարից։
Էուկարիոտներում տրանսկրիպցիայի ավարտը քիչ է ուսումնասիրված: Այն ավարտվում է ՌՆԹ-ի կտրումով, որից հետո ֆերմենտն իր 3» ծայրին ավելացնում է մի քանի ադենին (...AAAA), որոնց թիվը որոշում է տվյալ տառադարձության կայունությունը։
Տառադարձման գործարաններ
Կան մի շարք փորձարարական տվյալներ, որոնք ցույց են տալիս, որ տրանսկրիպցիան տեղի է ունենում, այսպես կոչված, տրանսկրիպցիոն գործարաններում. հսկայական, ըստ որոշ գնահատականների, մինչև 10 Da համալիրներ, որոնք պարունակում են մոտ 8 ՌՆԹ պոլիմերազ II և բաղադրիչներ հետագա մշակման և միացման, ինչպես նաև ուղղման համար: նոր սինթեզված արտագրության. Բջջային միջուկում մշտական փոխանակում է լուծվող և ակտիվացված ՌՆԹ պոլիմերազի լողավազանների միջև: Ակտիվ ՌՆԹ պոլիմերազը ներգրավված է նման բարդույթում, որն իր հերթին քրոմատինի խտացում կազմակերպող կառուցվածքային միավոր է։ Վերջին տվյալները ցույց են տալիս, որ տրանսկրիպցիոն գործարանները գոյություն ունեն նույնիսկ տառադարձման բացակայության դեպքում, դրանք ամրագրված են բջջում (դեռ պարզ չէ՝ փոխազդում են բջջի միջուկային մատրիցայի հետ, թե ոչ) և ներկայացնում են անկախ միջուկային ենթաբաժին։ ՌՆԹ I, II կամ III պոլիմերազ պարունակող տրանսկրիպցիոն գործարանային համալիրը վերլուծվել է զանգվածային սպեկտրոմետրիայի միջոցով:
Հակադարձ արտագրում
Հակադարձ արտագրման սխեմաՈրոշ վիրուսներ (օրինակ՝ ՄԻԱՎ-ը, որն առաջացնում է ՁԻԱՀ), ունեն ՌՆԹ-ն ԴՆԹ-ի արտագրելու հատկություն: ՄԻԱՎ-ն ունի ՌՆԹ գենոմ, որը ինտեգրված է ԴՆԹ-ին: Արդյունքում վիրուսի ԴՆԹ-ն կարող է զուգակցվել ընդունող բջջի գենոմի հետ։ ՌՆԹ-ից ԴՆԹ-ի սինթեզման համար պատասխանատու հիմնական ֆերմենտը կոչվում է հակադարձազա: Ռեվերսետազի գործառույթներից մեկը վիրուսի գենոմից կոմպլեմենտար ԴՆԹ (cDNA) ստեղծումն է: Կապակցված ռիբոնուկլեազ H ֆերմենտը կտրում է ՌՆԹ-ն, իսկ ռեվերսազը սինթեզում է cDNA-ն ԴՆԹ-ի կրկնակի պարույրից: cDNA-ն ինտեգրվում է հյուրընկալող բջիջի գենոմի մեջ ինտեգրազի միջոցով: Արդյունքը հյուրընկալ բջիջի կողմից վիրուսային սպիտակուցների սինթեզն է, որոնք ձևավորում են նոր վիրուսներ։ ՄԻԱՎ-ի դեպքում ծրագրավորված է նաև T-լիմֆոցիտների ապոպտոզը (բջջային մահը): Այլ դեպքերում բջիջը կարող է մնալ վիրուսներ տարածող:
Որոշ էուկարիոտ բջիջներ պարունակում են տելոմերազ ֆերմենտ, որը նույնպես ցուցադրում է հակադարձ տրանսկրիպցիոն ակտիվություն։ Նրա օգնությամբ սինթեզվում են ԴՆԹ-ում կրկնվող հաջորդականությունները։ Տելոմերազը հաճախ ակտիվանում է քաղցկեղի բջիջներում՝ անորոշ ժամանակով կրկնօրինակելու գենոմը՝ չկորցնելով սպիտակուցը կոդավորող ԴՆԹ-ի հաջորդականությունը:
Նշումներ
Վիքիմեդիա հիմնադրամ. 2010 թ.
Տեսեք, թե ինչ է «Տրանսկրիպցիան (կենսաբանություն)» այլ բառարաններում.
- (լատիներեն transcriptio, լիտ. վերագրում), ՌՆԹ մոլեկուլների կենսասինթեզ, ընդ. ԴՆԹ հատվածներ; գենետիկական ներդրման առաջին փուլը. տեղեկատվություն կենդանի բջիջներում: Այն իրականացվում է ԴՆԹ-կախյալ ՌՆԹ պոլիմերազ ֆերմենտի միջոցով, դեպի դրախտ ամենաշատ ուսումնասիրված... ... Կենսաբանական հանրագիտարանային բառարան
ԿենսաբանությունԿԵՆՍԱԲԱՆՈՒԹՅՈՒՆ (հունարեն bio life և logos բառից, վարդապետություն) կյանքի մասին գիտությունների ամբողջությունը Երկրի վրա նրա ձևերի, հատկությունների, կապերի և հարաբերությունների դրսևորումների ողջ բազմազանությամբ: Տերմինը առաջին անգամ առաջարկվել է միաժամանակ և ինքնուրույն 1802 թվականին... ... Իմացաբանության և գիտության փիլիսոփայության հանրագիտարան
Կյանքի գիտություն, որն իր մեջ ներառում է կենդանի էակների բնույթի, կառուցվածքի, ֆունկցիայի և վարքագծի մասին բոլոր գիտելիքները։ Կենսաբանությունը զբաղվում է ոչ միայն տարբեր օրգանիզմների ձևերի մեծ բազմազանությամբ, այլև դրանց էվոլյուցիայի, զարգացման և այն հարաբերությունների հետ, որոնք... ... Collier's Encyclopedia
ԿԵՆՍԱԲԱՆՈՒԹՅՈՒՆ- կյանքի մասին գիտությունների մի շարք Երկրի վրա նրա ձևերի, հատկությունների, կապերի և հարաբերությունների դրսևորումների ամբողջ բազմազանության մեջ: Տերմինը առաջին անգամ առաջարկվել է միաժամանակ և միմյանցից անկախ 1802 թվականին ականավոր ֆրանսիացի գիտնական Ջ.Բ. Լամարկը և գերմանացիները... Գիտության փիլիսոփայություն. Հիմնական տերմինների բառարան
I Տառադարձում (լատիներեն transcriptio rewriting-ից) բառերի և տեքստերի գրավոր վերարտադրություն՝ հաշվի առնելով դրանց արտասանությունը՝ օգտագործելով որոշակի գրաֆիկական համակարգ։ Գիտական եւ գործնական կարող են լինել Տ. Լեզվաբանության մեջ օգտագործվում է գիտական Տ.
- (լատիներեն transcriptio-ից, տառերի վերագրում), ՌՆԹ-ի կենսասինթեզ ԴՆԹ-ի մատրիցով; գենետիկական ներդրման առաջին փուլը. տեղեկատվությունը, ԴՆԹ-ի նուկլեոտիդային հաջորդականությունը կտրելու ժամանակ կարդացվում է ՌՆԹ նուկլեոտիդային հաջորդականության տեսքով (տես Գենետիկ կոդը) ... Քիմիական հանրագիտարան
Pre mRNA ցողունային հանգույցով: Հիմքերում ազոտի ատոմներն ընդգծված են կապույտով, թթվածնի ատոմները՝ մոլեկուլի ֆոսֆատային ողնաշարում՝ կարմիր:
Գիտություն, որի նպատակն է հասկանալ կյանքի երևույթների բնույթը՝ ուսումնասիրելով կենսաբանական առարկաները և համակարգերը մոլեկուլային մակարդակին մոտենալու և որոշ դեպքերում հասնելով այս սահմանին մակարդակի վրա։ Վերջնական նպատակն է...... Խորհրդային մեծ հանրագիտարան
Հակադարձ տրանսկրիպցիան միաշղթա ՌՆԹ կաղապարից կրկնակի շղթա ԴՆԹ-ի արտադրության գործընթացն է: Այս գործընթացը կոչվում է հակադարձ տառադարձում, քանի որ գենետիկական տեղեկատվության փոխանցումը տեղի է ունենում «հակադարձում», համեմատաբար ... ... Վիքիպեդիա
«Վիրուս» հարցումը վերահղված է այստեղ։ Տեսնել նաև այլ իմաստներ։ ? Վիրուսներ Ռոտավիրուս Գիտական դասակարգում Overkingdom ... Վիքիպեդիա
Գենետիկ կոդը վերծանելուց հետո հարց առաջացավ՝ ինչպե՞ս է ինֆորմացիան ԴՆԹ-ից տեղափոխվում սպիտակուց։ Կենսաքիմիական ուսումնասիրությունները ցույց են տվել, որ բջջի ԴՆԹ-ի հիմնական մասը տեղայնացված է միջուկում, մինչդեռ սպիտակուցի սինթեզը տեղի է ունենում ցիտոպլազմայում: ԴՆԹ-ի և սպիտակուցի սինթեզի այս տարածքային տարանջատումը հանգեցրեց միջնորդի որոնմանը: Քանի որ սպիտակուցի սինթեզը տեղի էր ունենում ռիբոսոմների մասնակցությամբ, ՌՆԹ-ն առաջ քաշվեց միջնորդի դեր խաղալու համար։ Ստեղծվել է դիագրամ, որը ցույց է տալիս բջջում գենետիկական տեղեկատվության հոսքի ուղղությունը.
ԴՆԹ → ՌՆԹ → սպիտակուց
Այն կոչվում է մոլեկուլային կենսաբանության կենտրոնական դոգմա: Ֆ. Կրիկը պնդեց, որ մակրոմոլեկուլների սինթեզն ըստ այս սխեմայի իրականացվում է մատրիցային սկզբունքով։ Այս պոստուլատի ճիշտությունն ապացուցելու համար երկար տարիներ պահանջվեցին։
Սկզբում ենթադրվում էր, որ ռիբոսոմային ՌՆԹ-ն («մեկ գեն - մեկ ռիբոսոմ - մեկ սպիտակուց») միջնորդի դեր է խաղում: Սակայն շուտով պարզ դարձավ, որ այս ենթադրությունն անհիմն էր։ Ցույց է տրվել, որ սպիտակուցների սինթեզի ընթացքում ռիբոսոմների թիվը չի փոխվում, այսինքն. նոր ՌՆԹ չի սինթեզվում և, հետևաբար, նոր տեղեկատվություն չի ստացվում: Շուտով ռիբոսոմների բաղադրության մեջ հայտնաբերվեց անկայուն ՌՆԹ-ի մասնաբաժին, որի մոլեկուլները մագնի կատիոնների օգնությամբ ազատորեն պահվում են ռիբոսոմի վրա։ Օգտագործելով մոլեկուլային հիբրիդացում՝ ցույց է տրվել, որ այս ՌՆԹ-ի մոլեկուլները ԴՆԹ-ի որոշակի հատվածների պատճեններն են։ Նա ստացել է անունը մատրիցա, կամ սուրհանդակ ՌՆԹ. Այն նախկինում նաև կոչվում էր սուրհանդակային ՌՆԹ և սուրհանդակ ՌՆԹ: Այս մոլեկուլների փոխլրացումը ԴՆԹ-ի որոշ հատվածներին ցույց է տալիս, որ դրանք սինթեզվել են ըստ ԴՆԹ-ի կաղապարի տեսակի:
Աստիճանաբար պարզվեց ԴՆԹ-ից դեպի սպիտակուց տեղեկատվության փոխանցման ողջ ճանապարհը։ Այն բաղկացած է երկու փուլից. արտագրություններԵվ հեռարձակումներ. Տրանսկրիպցիայի փուլում գենետիկական ինֆորմացիան ընթերցվում և ԴՆԹ-ից փոխանցվում է mRNA: Տառադարձման գործընթացը տեղի է ունենում երեք փուլով. ընդունելը, երկարացումԵվ ավարտ. Տեղեկատվությունը կարդացվում է միայն մեկ ԴՆԹ շղթայից (+ շղթայից), քանի որ, ելնելով գենետիկ կոդի հատկություններից, ԴՆԹ-ի լրացուցիչ բաժինները չեն կարող կոդավորել նույն սպիտակուցի կառուցվածքը՝ կոդի կոմպլեմենտար այլասերվածության բացակայության պատճառով: Տրանսկրիպցիան իրականացվում է ՌՆԹ պոլիմերազ ֆերմենտի միջոցով, որը բաղկացած է չորս ենթամիավորներից (ααββ») և չունի կոնկրետություն ԴՆԹ-ի աղբյուրի հետ կապված: Տրանսկրիպցիայի սկզբնական փուլում՝ սկիզբը, հինգերորդ ենթամիավորը, այսպես կոչված, s-գործոնը: , կցվում է ԴՆԹ-ի որոշակի հատվածը ճանաչող ֆերմենտին, պրոմոտորին: Պրոմոտորները չեն տառադարձվում: Նրանք ճանաչվում են s-գործոնով դրանցում հատուկ նուկլեոտիդային հաջորդականության առկայությամբ: Բակտերիալ պրոմոտորներում այն կոչվում է Պրիբնով: բլոկ և ունի TATAAT ձև (թեթև տատանումներով): ՌՆԹ պոլիմերազ ֆերմենտը կցվում է խթանողին: mRNA շղթայի աճը տեղի է ունենում մեկ ուղղությամբ, տրանսկրիպցիայի արագությունը ≈ 45-50 նուկլեոտիդ է վայրկյանում: Մեկնարկային փուլում, սինթեզվում է ընդամենը 8 նուկլեոտիդից բաղկացած կարճ շղթա, որից հետո s-գործոնն առանձնանում է ՌՆԹ պոլիմերազից և սկսվում է երկարացման փուլը: mRNA շղթայի երկարացումը կատարվում է տետրամեր սպիտակուցի միջոցով: Բաժին, որտեղից կարդացվում է տեղեկատվությունը. կոչվում է տրանսկրիպտոն: Այն ավարտվում է տերմինատորով` հատուկ նուկլեոտիդային հաջորդականությամբ, որը կատարում է կանգառի ազդանշանի դեր: Հասնելով տերմինատորին՝ ՌՆԹ պոլիմերազային ֆերմենտը դադարում է գործել և սպիտակուցի վերջացման գործոնների օգնությամբ առանձնանում է մատրիցից։

Բակտերիալ բջիջներում ստացված mRNA մոլեկուլները կարող են անմիջապես ծառայել որպես սպիտակուցների սինթեզի ձևանմուշներ, այսինքն. հեռարձակում. Նրանք միանում են ռիբոսոմներին, որոնց փոխադրող ՌՆԹ (tRNA) մոլեկուլները միաժամանակ փոխանցում են ամինաթթուներ։ Տրանսֆերային ՌՆԹ շղթաները բաղկացած են մոտավորապես 70 նուկլեոտիդներից: Միաշղթա tRNA մոլեկուլն ունի կոմպլեմենտար զուգավորման վայրեր, որոնք պարունակում են ակտիվ կենտրոններ. tRNA սինթետազ ֆերմենտի կողմից tRNA-ի ճանաչման վայր, որը համապատասխան ակտիվացված ամինաթթուն է կապում tRNA-ին; ակցեպտոր - այն վայրը, որին կցված է ամինաթթուն, և հակակոդոնային հանգույցը:

Հակակոդոնեռյակ է, որը լրացնում է mRNA մոլեկուլի համապատասխան կոդոնին։ Կոդոն-հակակոդոն փոխազդեցությունը հետևում է կոմպլեմենտար զուգավորման տեսակին, որի ընթացքում աճող սպիտակուցային շղթային ավելացվում է ամինաթթու: Տարբեր mRNA-ներում մեկնարկային կոդոնը AUG կոդոնն է, որը համապատասխանում է մեթիոնին ամինաթթվին։ Հետևաբար, tRNA-ն UAC հակակոդոնով, որը կապված է ակտիվացված ամինաթթվի մեթիոնինի հետ, առաջինն է մոտենում մատրիցին: Ֆերմենտները, որոնք ակտիվացնում են ամինաթթուները և միացնում դրանք tRNA-ին, կոչվում են aminoacyl-tRNA սինթետազներ: Սպիտակուցների կենսասինթեզի բոլոր փուլերը (սկիզբ, երկարացում, վերջացում) սպասարկվում են սպիտակուցների թարգմանության գործոններով։ Պրոկարիոտները յուրաքանչյուր փուլի համար ունեն դրանցից երեքը: mRNA կաղապարի վերջում կան անհեթեթ կոդոններ, որոնք չեն կարդացվում և նշում են թարգմանության ավարտը։
Բազմաթիվ օրգանիզմների գենոմում՝ բակտերիայից մինչև մարդ, հայտնաբերվել են գեներ և համապատասխան tRNA-ներ, որոնք իրականացնում են կոդոնների ոչ ստանդարտ ընթերցում։ Այս երեւույթը կոչվում է հեռարձակման անորոշություն.

Այն թույլ է տալիս խուսափել սխալների բացասական հետևանքներից, որոնք տեղի են ունենում mRNA մոլեկուլների կառուցվածքում տրանսկրիպցիայի ժամանակ: Այսպիսով, երբ mRNA մոլեկուլի ներսում հայտնվում են անհեթեթ կոդոններ, որոնք ունակ են վաղաժամ դադարեցնել տրանսկրիպցիայի գործընթացը, ակտիվանում է ճնշելու մեխանիզմը։ Այն բաղկացած է նրանից, որ բջջում հայտնվում է tRNA-ի անսովոր ձև՝ անհեթեթ կոդոնին լրացնող հակակոդոնով, որը սովորաբար չպետք է գոյություն ունենա: Դրա տեսքը արդյունք է գենի գործողության, որը փոխարինում է tRNA հակակոդոնի հիմքը, որն իր կազմով նման է անհեթեթ կոդոնին։ Այս փոխարինման արդյունքում անհեթեթ կոդոնը կարդացվում է որպես կանոնավոր նշանակալի կոդոն։ Նման մուտացիաները կոչվում են ճնշող մուտացիաներ, քանի որ նրանք ճնշում են սկզբնական մուտացիան, որը հանգեցրեց անհեթեթ կոդոնի:
Ածխածնի տեսքով կյանքը գոյություն ունի սպիտակուցի մոլեկուլների առկայության պատճառով: Իսկ բջջում սպիտակուցի կենսասինթեզը գեների արտահայտման միակ հնարավորությունն է։ Բայց այս գործընթացն իրականացնելու համար անհրաժեշտ է սկսել մի շարք գործընթացներ, որոնք կապված են գենետիկ տեղեկատվության «փաթեթավորման», ցանկալի գենի որոնման, այն կարդալու և վերարտադրելու հետ: Կենսաբանության մեջ «տրանսկրիպցիա» տերմինը մասնավորապես վերաբերում է գենից հաղորդագրություն փոխանցող ՌՆԹ տեղեկատվության փոխանցման գործընթացին: Սա կենսասինթեզի, այսինքն՝ գենետիկ տեղեկատվության անմիջական ներդրման սկիզբն է։
Գենետիկական տեղեկատվության պահպանում
Կենդանի օրգանիզմների բջիջներում գենետիկական տեղեկատվությունը տեղայնացված է միջուկում, միտոքոնդրիումներում, քլորոպլաստներում և պլազմիդներում։ Միտոքոնդրիումներն ու քլորոպլաստները պարունակում են փոքր քանակությամբ կենդանիների և բույսերի ԴՆԹ, մինչդեռ բակտերիալ պլազմիդները հանդիսանում են շրջակա միջավայրի պայմաններին արագ հարմարվելու համար պատասխանատու գեների պահեստավորման վայրը:
Վիրուսային մարմիններում ժառանգական տեղեկատվությունը պահվում է նաև ՌՆԹ-ի կամ ԴՆԹ-ի պոլիմերների տեսքով։ Բայց դրա իրականացման գործընթացը կապված է նաև արտագրման անհրաժեշտության հետ։ Կենսաբանության մեջ այս գործընթացը բացառիկ նշանակություն ունի, քանի որ հենց դա է հանգեցնում ժառանգական տեղեկատվության ներդրմանը, որը հրահրում է սպիտակուցի կենսասինթեզը:

Կենդանական բջիջներում ժառանգական տեղեկատվությունը ներկայացված է ԴՆԹ-ի պոլիմերով, որը կոմպակտ փաթեթավորված է միջուկի ներսում: Հետևաբար, նախքան սպիտակուցի սինթեզը կամ որևէ գենի ընթերցումը, պետք է անցնեն որոշակի փուլեր՝ խտացրած քրոմատինի լուծարում և ցանկալի գենի «ազատում», ֆերմենտային մոլեկուլների կողմից դրա ճանաչում, տրանսկրիպցիա։
Կենսաբանության և կենսաբանական քիմիայի մեջ այս փուլերն արդեն ուսումնասիրված են։ Դրանք հանգեցնում են սպիտակուցի սինթեզի, որի առաջնային կառուցվածքը կոդավորված էր մեկ գենում։
Էուկարիոտիկ բջիջներում տրանսկրիպցիոն օրինաչափություն
Թեև կենսաբանության մեջ տրանսկրիպցիան բավականաչափ ուսումնասիրված չէ, դրա հաջորդականությունը ավանդաբար ներկայացվում է դիագրամի տեսքով։ Այն բաղկացած է մեկնարկից, երկարացումից և ավարտից: Սա նշանակում է, որ ամբողջ գործընթացը բաժանված է երեք բաղադրիչ երևույթների.

Նախաձեռնումը կենսաբանական և կենսաքիմիական գործընթացների մի շարք է, որոնք հանգեցնում են տրանսկրիպցիայի սկզբին: Երկարացման էությունը մոլեկուլային շղթայի շարունակական աճն է։ Դադարեցումը գործընթացների մի շարք է, որոնք հանգեցնում են ՌՆԹ-ի սինթեզի դադարեցմանը: Ի դեպ, սպիտակուցների կենսասինթեզի համատեքստում կենսաբանության մեջ տրանսկրիպցիայի գործընթացը սովորաբար նույնացվում է սուրհանդակային ՌՆԹ-ի սինթեզի հետ։ Դրա հիման վրա հետագայում կսինթեզվի պոլիպեպտիդային շղթա։

Ընդունելը
Նախաձեռնումը կենսաբանության մեջ ամենաքիչ հասկացված տրանսկրիպցիոն մեխանիզմն է: Ինչ է դա կենսաքիմիական տեսանկյունից, անհայտ է: Այսինքն՝ տրանսկրիպցիան հրահրելու համար պատասխանատու հատուկ ֆերմենտներն ընդհանրապես չեն ճանաչվում։ Անհայտ են նաև ներբջջային ազդանշաններն ու դրանց փոխանցման եղանակները, որոնք վկայում են նոր սպիտակուցի սինթեզի անհրաժեշտության մասին։ Սա բջջաբանության և կենսաքիմիայի հիմնարար խնդիր է:
Երկարացում
Դեռևս հնարավոր չէ ժամանակին տարանջատել մեկնարկի և երկարացման գործընթացը՝ հատուկ ֆերմենտների և հրահրող գործոնների առկայությունը հաստատելու համար նախատեսված լաբորատոր հետազոտությունների անցկացման անհնարինության պատճառով: Հետեւաբար, այս սահմանը շատ պայմանական է։ Երկարացման գործընթացի էությունը հանգում է ԴՆԹ-ի կաղապարի հատվածի հիման վրա սինթեզված աճող շղթայի երկարացմանը:
Ենթադրվում է, որ երկարացումը սկսվում է ՌՆԹ պոլիմերազի առաջին տեղափոխումից և ՌՆԹ-ի սկզբնական տեղամասին առաջին կադոնի կցման սկզբից հետո։ Երկարացման ժամանակ կադոնները կարդում են 3"-5" շղթայի ուղղությամբ երկու շղթայի բաժանված ԴՆԹ-ի հյուծված հատվածի վրա: Միևնույն ժամանակ, աճող ՌՆԹ-ի շղթան ավելացվում է նոր նուկլեոտիդներով, որոնք լրացնում են կաղապարային ԴՆԹ-ի շրջանը: Այս դեպքում ԴՆԹ-ն «ընդլայնվում է» մինչև 12 նուկլեոտիդ, այսինքն՝ 4 կադոն:

ՌՆԹ պոլիմերազ ֆերմենտը շարժվում է աճող շղթայի երկայնքով, և դրա «ետևում» ԴՆԹ-ն հակառակ «խաչ կապված» է երկշղթա կառուցվածքի մեջ՝ նուկլեոտիդների միջև ջրածնային կապերի վերականգնմամբ: Սա մասամբ պատասխանում է այն հարցին, թե ինչ գործընթաց է կոչվում տրանսկրիպցիա կենսաբանության մեջ: Հենց երկարացումն է տրանսկրիպցիայի հիմնական փուլը, քանի որ դրա ընթացքում հավաքվում է այսպես կոչված միջնորդը գենի և սպիտակուցի սինթեզի միջև։
Ավարտ
Էուկարիոտիկ բջիջներում տրանսկրիպցիայի ավարտի գործընթացը վատ է հասկացված: Մինչ այժմ գիտնականները կրճատել են դրա էությունը՝ դադարեցնելով ԴՆԹ-ի ընթերցումը 5 դյույմանոց ծայրում և մի խումբ ադենինային հիմքեր կցելով ՌՆԹ-ի 3 դյույմ ծայրին: Վերջին գործընթացը թույլ է տալիս կայունացնել ստացված ՌՆԹ-ի քիմիական կառուցվածքը։ Բակտերիալ բջիջներում գոյություն ունի դադարեցման երկու տեսակ. Դա Rho-կախյալ և Rho-անկախ գործընթաց է:

Առաջինը տեղի է ունենում Rho սպիտակուցի առկայության դեպքում և վերածվում է ջրածնային կապերի պարզ խզման ԴՆԹ-ի կաղապարային շրջանի և սինթեզված ՌՆԹ-ի միջև: Երկրորդը, Rho-անկախ, տեղի է ունենում ցողունային հանգույցի հայտնվելուց հետո, եթե դրա հետևում կա ուրացիլային հիմքերի մի շարք: Այս համակցությունը հանգեցնում է նրան, որ ՌՆԹ-ն անջատվում է ԴՆԹ-ի կաղապարից: Ակնհայտ է, որ տրանսկրիպցիայի ավարտը ֆերմենտային գործընթաց է, սակայն դրա համար հատուկ կենսակատալիզատորներ դեռ չեն հայտնաբերվել։
Վիրուսային տառադարձում
Վիրուսային մարմինները չունեն իրենց սեփական սպիտակուցի կենսասինթեզի համակարգը և, հետևաբար, չեն կարող վերարտադրվել առանց բջիջների շահագործման: Բայց վիրուսներն ունեն իրենց գենետիկական նյութը, որը պետք է գիտակցել և նաև ինտեգրել վարակված բջիջների գեներին։ Դա անելու համար նրանք ունեն մի շարք ֆերմենտներ (կամ շահագործում են բջջային ֆերմենտային համակարգերը), որոնք արտագրում են իրենց նուկլեինաթթուն։ Այսինքն՝ այս ֆերմենտը, հիմնվելով վիրուսի գենետիկ տեղեկատվության վրա, սինթեզում է մեսենջեր ՌՆԹ-ի անալոգը։ Բայց դա ամենևին էլ ՌՆԹ չէ, այլ ԴՆԹ պոլիմեր, որը լրացնում է, օրինակ, մարդու գեները։
Սա լիովին խախտում է կենսաբանության մեջ տրանսկրիպցիայի ավանդական սկզբունքները, ինչպես երևում է ՄԻԱՎ վիրուսի օրինակից։ Դրա հակադարձ ֆերմենտային ֆերմենտը ունակ է վիրուսային ՌՆԹ-ից մարդու նուկլեինաթթվին լրացնող ԴՆԹ սինթեզելու: ՌՆԹ-ից կոմպլեմենտար ԴՆԹ-ի սինթեզման գործընթացը կոչվում է հակադարձ տրանսկրիպցիա: Սա կենսաբանության մեջ բնորոշվում է այն գործընթացի, որը պատասխանատու է վիրուսի ժառանգական տեղեկատվության մարդու գենոմում ինտեգրվելու համար:
Կոլպինոյում լոգարանների վերականգնում vk.com/restavraciya_vann_kolpino.
Տառադարձում. Սկիզբ - տառադարձման սկիզբ, Ավարտ - տառադարձման ավարտ, ԴՆԹ - ԴՆԹ:
Տրանսկրիպցիան ՌՆԹ-ի սինթեզի գործընթաց է՝ օգտագործելով ԴՆԹ-ն որպես ձևանմուշ և տեղի է ունենում բոլոր կենդանի բջիջներում: Այլ կերպ ասած, դա գենետիկ տեղեկատվության փոխանցումն է ԴՆԹ-ից ՌՆԹ:
Տրանսկրիպցիան կատալիզացվում է ԴՆԹ-կախյալ ՌՆԹ պոլիմերազ ֆերմենտի միջոցով: ՌՆԹ-ի սինթեզի գործընթացն ընթանում է 5"-ից մինչև 3" ծայր ուղղությամբ, այսինքն՝ ԴՆԹ-ի կաղապարի շղթայի երկայնքով ՌՆԹ պոլիմերազը շարժվում է 3"->5" ուղղությամբ:
Տրանսկրիպցիան բաղկացած է մեկնարկի, երկարացման և ավարտի փուլերից:
Տառադարձման նախաձեռնում
Տրանսկրիպցիան սկսելը բարդ գործընթաց է, որը կախված է տառադարձված հաջորդականության մոտակայքում գտնվող ԴՆԹ-ի հաջորդականությունից և տարբեր սպիտակուցային գործոնների առկայությունից կամ բացակայությունից:
Տառադարձման երկարացում
Այն պահը, երբ ՌՆԹ պոլիմերազն անցնում է տրանսկրիպցիայի մեկնարկից դեպի երկարացում, ճշգրիտ որոշված չէ: Երեք հիմնական կենսաքիմիական իրադարձությունները բնութագրում են այս անցումը Escherichia coli ՌՆԹ պոլիմերազի դեպքում. պոլիմերազ, ներառում է աճող ՌՆԹ շղթան և տառադարձված ԴՆԹ-ն։ Նույն երեւույթները բնորոշ են նաեւ էուկարիոտ ՌՆԹ պոլիմերազներին։ Անցումը մեկնարկից դեպի երկարացում ուղեկցվում է ֆերմենտի, պրոմոտորի, տրանսկրիպցիան մեկնարկող գործոնների միջև կապերի խզմամբ, իսկ որոշ դեպքերում՝ ՌՆԹ պոլիմերազի անցումով երկարացման կոմպետենտության վիճակի։ Երկարացման փուլն ավարտվում է աճող տառադարձման թողարկումից հետո, և ֆերմենտը կաղապարից անջատվում է:
Երկարացման փուլում մոտավորապես 18 նուկլեոտիդային զույգ չի ոլորվում ԴՆԹ-ում: ԴՆԹ-ի կաղապարի մոտ 12 նուկլեոտիդները կազմում են հիբրիդային պարույր ՌՆԹ շղթայի աճող ծայրով: Երբ ՌՆԹ պոլիմերազը շարժվում է կաղապարի միջով, ԴՆԹ-ի կրկնակի պարույրի լուծարումը տեղի է ունենում դրանից առաջ, իսկ ԴՆԹ-ի կրկնակի պարույրի վերականգնումը տեղի է ունենում դրա հետևում: Միևնույն ժամանակ, կաղապարով և ՌՆԹ պոլիմերազով համալիրից ազատվում է աճող ՌՆԹ շղթայի հաջորդ օղակը։ Այս շարժումները պետք է ուղեկցվեն ՌՆԹ պոլիմերազի և ԴՆԹ-ի հարաբերական պտույտով։ Դժվար է պատկերացնել, թե ինչպես դա կարող է տեղի ունենալ բջջում, հատկապես քրոմատինի արտագրման ժամանակ: Ուստի հնարավոր է, որ նման պտույտը կանխելու համար ԴՆԹ-ի երկայնքով շարժվող ՌՆԹ պոլիմերազն ուղեկցվի տոպոիզոմերազներով։
Երկարացումն իրականացվում է հիմնական երկարացման գործոնների օգնությամբ, որոնք անհրաժեշտ են, որպեսզի գործընթացը ժամանակից շուտ չդադարի։
Վերջերս ի հայտ եկան ապացույցներ, որոնք ցույց են տալիս, որ կարգավորիչ գործոնները կարող են նաև կարգավորել երկարացումը: Երկարացման գործընթացում ՌՆԹ պոլիմերազը կանգ է առնում գենի որոշ հատվածներում։ Սա հատկապես հստակ երևում է սուբստրատների ցածր կոնցենտրացիաների դեպքում: Մատրիցայի որոշ հատվածներում նկատվում են ՌՆԹ պոլիմերազի առաջխաղացման երկար ձգձգումներ, այսպես կոչված: դադարներ են նկատվում նույնիսկ սուբստրատի օպտիմալ կոնցենտրացիաների դեպքում: Այս դադարների տեւողությունը կարելի է վերահսկել երկարացման գործոններով։
| Տրիպտոֆան օպերոն |
Տրանսկրիպցիան սինթեզի գործընթացն էմոլեկուլներըՌՆԹ-ն միացված էտարածքԴՆԹ, օգտագործվում է որպես մատրիցա։ Տառադարձության իմաստն է գենետիկական տեղեկատվության փոխանցում ԴՆԹ-ից ՌՆԹ.
ԴՆԹ-ի մոլեկուլը բաղկացած է երկու լրացնող շղթաներից, մինչդեռ ՌՆԹ-ն բաղկացած է միայն մեկից: Տրանսկրիպցիայի ընթացքում ԴՆԹ-ի շղթաներից միայն մեկն է ծառայում որպես ՌՆԹ սինթեզի ձևանմուշ։ Նրան կանչում են իմաստային շղթա. Բացառություն է կազմում միտոքոնդրիալ ԴՆԹ-ն, որտեղ երկու շղթաներն էլ զգայական են և պարունակում են տարբեր գեներ։ Բացի միջուկային ԴՆԹ-ում բացառություն լինելուց, որոշ գեներ կարող են տեղայնացվել անհեթեթ շղթայի վրա:
Տրանսկրիպցիայի ընթացքում ՌՆԹ-ի մոլեկուլը սինթեզվում է 5"-ից մինչև 3" ծայրը (ինչը բնական է բոլոր նուկլեինաթթուների սինթեզի համար), մինչդեռ ԴՆԹ-ի շղթայի երկայնքով սինթեզն ընթանում է հակառակ ուղղությամբ՝ 3"→5: «.
Էուկարիոտներում յուրաքանչյուր գեն արտագրվում է առանձին։ Բացառություն, դարձյալ, միտոքոնդրիալ ԴՆԹ-ն է, որը տրանսկրիպտացվում է ընդհանուր բազմածին տառադարձման, որն այնուհետև կտրվում է: Քանի որ պրոկարիոտներում գեները կազմում են խմբեր՝ ձևավորելով մեկ օպերոն, այդպիսի գեները տառադարձվում են միասին։ Ինչեւէ տրանսկրիպտոնկոչվում է ԴՆԹ-ի մի հատված, որը բաղկացած է խթանողից, տառադարձված շրջանից և տերմինատորից:
Տառադարձման 3 փուլ կա. սկիզբ, երկարացում, վերջացում.
Ընդունելըտրանսկրիպցիան թույլ է տալիս սկսել ՌՆԹ մոլեկուլի սինթեզը: Նախաձեռնումը ներառում է ֆերմենտների համալիրի կցումը պրոմոտորին: Հիմնականը ՌՆԹ պոլիմերազն է (տվյալ դեպքում՝ ԴՆԹ-ից կախված), որն իր հերթին բաղկացած է մի քանի ենթամիավոր սպիտակուցներից և կատարում է գործընթացի կատալիզատորի դեր։ Էուկարիոտներում տրանսկրիպցիան սկսելու վրա ազդում են ԴՆԹ-ի հատուկ բաժինները՝ ուժեղացուցիչներ (ուժեղացնում են) և խլացուցիչները (ճնշված), որոնք սովորաբար գտնվում են գենից որոշակի հեռավորության վրա: Կան տարբեր սպիտակուցային գործոններ, որոնք ազդում են տրանսկրիպցիան սկսելու հնարավորության վրա:
Պրոկարիոտներն ունեն միայն մեկ տեսակի ՌՆԹ պոլիմերազ, իսկ էուկարիոտները՝ երեք։ ՌՆԹ պոլիմերազ 1-ն օգտագործվում է ռիբոսոմային ՌՆԹ-ի երեք տեսակների սինթեզման համար (ընդհանուր առմամբ կա 4 տեսակի rRNA): ՌՆԹ պոլիմերազ 2-ն օգտագործվում է նախամՌՆԹ (նախորդ սուրհանդակ ՌՆԹ) սինթեզելու համար։ ՌՆԹ պոլիմերազ-3-ը սինթեզում է ռիբոսոմային ՌՆԹ-ի, տրանսպորտային և փոքր միջուկային տեսակներից մեկը։
ՌՆԹ պոլիմերազն ի վիճակի է ճանաչել նուկլեոտիդների հատուկ հաջորդականությունը և կցել դրանց։ Այս հաջորդականությունները կարճ են և համընդհանուր բոլոր կենդանի էակների համար:
Այն բանից հետո, երբ ՌՆԹ պոլիմերազը կապվում է պրոմոտորին, ԴՆԹ-ի կրկնակի պարույրի մի հատվածը արձակվում է, և այս հատվածի շղթաների միջև նուկլեոտիդային կապերը կոտրվում են: Մոտավորապես 18 զույգ նուկլեոտիդ է բացահայտված։
Բեմի վրա երկարացումհաջորդական ավելացումը տեղի է ունենում ազատ նուկլեոտիդների փոխլրացման սկզբունքի համաձայն ազատված ԴՆԹ-ի հատվածին: ՌՆԹ պոլիմերազը միավորում է նուկլեոտիդները պոլիրիբոնուկլեոտիդային շղթայի մեջ։
ՌՆԹ-ի սինթեզի ընթացքում նրա մոտ 12 նուկլեոտիդները փոխլրացնող են և ժամանակավորապես կապված են ԴՆԹ նուկլեոտիդների հետ։ Երբ ՌՆԹ պոլիմերազը շարժվում է դրա դիմաց, ԴՆԹ շղթաները շեղվում են, իսկ հետևում դրանք «կարվում» են ֆերմենտների օգնությամբ։ ՌՆԹ շղթան աստիճանաբար աճում է և դուրս է գալիս ՌՆԹ պոլիմերազային համալիրից։
Կան երկարացման գործոններ, որոնք կանխում են տրանսկրիպցիայի վաղաժամ դադարեցումը։
ԱվարտՏրանսկրիպցիայի գործընթացը տեղի է ունենում տերմինատորի շրջանում, որը ճանաչվում է ՌՆԹ պոլիմերազի կողմից՝ շնորհիվ հատուկ սպիտակուցի վերջավորության գործոնների:
Շատ ադենինային նուկլեոտիդներ (պոլի-Ա) կցվում են սինթեզված ՌՆԹ-ի մոլեկուլի 3" ծայրին, որպեսզի կանխեն դրա ֆերմենտային քայքայումը: Նույնիսկ ավելի վաղ, երբ սինթեզվում էր 5" ծայրը, այսպես կոչված. գլխարկ.
Շատ դեպքերում, տրանսկրիպցիան չի արտադրում պատրաստի ՌՆԹ: Հում ՌՆԹ-ն դեռ պետք է գործընթաց անցնի վերամշակում, որի ժամանակ տեղի են ունենում դրա մոդիֆիկացիայի փոփոխություններ և այն դառնում է ֆունկցիոնալ ակտիվ: Էուկարիոտների ՌՆԹ-ի յուրաքանչյուր տեսակ ենթարկվում է իր փոփոխությունների: Պոլի-Ա-ի և գլխարկի ձևավորումը հաճախ կոչվում է նաև վերամշակում:
Մենք նաև խորհուրդ ենք տալիս
 Թվային սիմվոլիկան Հովհաննեսի Հայտնության գրքում
Թվային սիմվոլիկան Հովհաննեսի Հայտնության գրքում
 Ուսուցման դաս «Կոնֆլիկտոլոգիա»՝ արաբ-իսրայելական հակամարտություն
Ուսուցման դաս «Կոնֆլիկտոլոգիա»՝ արաբ-իսրայելական հակամարտություն
 Հրաշք օձի հետ, որ պատահեց Սուրբ Մեծ նահատակ Գեորգի հետ
Հրաշք օձի հետ, որ պատահեց Սուրբ Մեծ նահատակ Գեորգի հետ
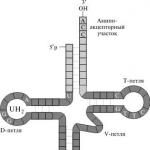 ՌՆԹ-ն ստանում է ժառանգական տեղեկատվություն
ՌՆԹ-ն ստանում է ժառանգական տեղեկատվություն
 Սկաևոլա, էտրուսկների հետ պատերազմի հերոս Գայուս Մուսիուսի պատվավոր անունը
Սկաևոլա, էտրուսկների հետ պատերազմի հերոս Գայուս Մուսիուսի պատվավոր անունը
 Որտե՞ղ է նախադրյալը: Ի՞նչ է «պրեդիկատը»: Պրեդիկատների տեսակները. Ճի՞շտ է, որ պարզ բառային նախադեպը այն նախադեպն է, որի բառային և քերականական իմաստը արտահայտվում է մեկ բայով:
Որտե՞ղ է նախադրյալը: Ի՞նչ է «պրեդիկատը»: Պրեդիկատների տեսակները. Ճի՞շտ է, որ պարզ բառային նախադեպը այն նախադեպն է, որի բառային և քերականական իմաստը արտահայտվում է մեկ բայով: