Функционални области на тРНК. Структура и функции на тРНК, особености на активиране на аминокиселини
70-90N | вторична страница - детелина | CCA 3" const за всички tRNA |
наличието на тимин, псевдоуридин-пси, дигируридин DGU в D-примката - защита срещу рибонуклеази? дълголетни | Разнообразие от първични структури на tRNA - 61 + 1 - по броя на кодоните + формилметионин tRNA, антикодонът на котката е същият като този на метиониновата tRNA. Разнообразие от третични структури - 20 (според броя на аминокиселините) | разпознаване - образуването на ковалентна връзка m-y tRNA и действат | аминоацил-тРНК синтетазите прикрепят действия към тРНК
Функцията на tRNA е да пренася аминокиселини от цитоплазмата към рибозомите, в които се осъществява синтеза на протеин.
тРНК, които свързват една аминокиселина, се наричат изоакцепторни.
Общо 64 различни tRNAs едновременно съществуват в клетка.
Всяка tRNA се сдвоява само със собствен кодон.
Всяка tRNA разпознава свой собствен кодон без участието на аминокиселина. Свързаните с tRNA аминокиселини бяха химически модифицирани, след което полученият полипептид, който съдържаше модифицираната аминокиселина, беше анализиран. Цистеинил-tRNACys (R=CH2-SH) се редуцира до аланил-tRNACys (R=CH3).
Повечето тРНК, независимо от тяхната нуклеотидна последователност, имат вторична структура с форма на детелина, поради наличието на три фиби в нея.
Структурни особености на тРНК
Винаги има четири несдвоени нуклеотида в 3 "края на молекулата и три от тях са задължително CCA. 5" и 3" краищата на РНК веригата образуват акцепторно стъбло. Веригите се държат заедно поради комплементарното сдвояване на седем нуклеотида 5 "- завършват със седем нуклеотида, разположени близо до края 3". 2. Всички молекули имат фиби T? C, обозначени така, защото съдържат два необичайни остатъка: риботимидин (T) и псевдоуридин (? Фибичката се състои от двоен -верижно стебло от пет сдвоени бази, включително двойката G-C, и бримка с дължина от седем нуклеотида.
в същата точка в цикъла. 3. В антикодон фиби, стъблото винаги е представено от семейство сдвоени
основания. Триплетът, комплементарен на свързания кодон, антикодона, се намира в бримката.
le, състоящ се от седем нуклеотида. Инвариантна ура-
цил и модифициран цитозин, и модифициран пурин прилепва към неговия 3" край, като правило
аденин. 4. Друга фиби се състои от дръжка с дължина три до четири двойки нуклеотиди и променлива бримка
размер, често съдържащ урацил в намалена форма - дихидроурацил (DU). Най-силно варират нуклеотидните последователности на стъблата, броят на нуклеотидите между стъблото на антикодон и T?C ствола (променлива бримка), както и размерът на бримката и локализацията на дихидроурациловите остатъци в DU бримката.
[Певец, 1998].
Третична структура на тРНК
L-образна структура.
Прикрепване на аминокиселини към tRNA
За да може една аминокиселина да образува полипептидна верига, тя трябва да бъде прикрепена към tRNA чрез ензима аминоацил-tRNA синтетаза. Този ензим образува ковалентна връзка между аминокиселинната карбоксилна група и рибозната хидроксилна група в 3' края на tRNA с участието на АТФ. Аминоацил-тРНК синтетазата разпознава специфичен кодон не поради наличието на антикодон върху тРНК, а чрез наличието на специфично място за разпознаване върху тРНК.
Общо в клетката има 21 различни аминоацил-тРНК синтетази.
Присъединяването става на два етапа:
1. Карбоксилната група на аминокиселина е прикрепена към ATP a-фосфат. Полученият нестабилен аминоацил аденилат се стабилизира чрез свързване с ензима.
2. Прехвърляне на аминоацилната група на аминоацил аденилат към 2’ или 3’-OH групата на крайната рибоза на tRNA
Някои аминоацил-тРНК синтетази се състоят от една полипептидна верига, докато други се състоят от две или четири идентични вериги, всяка с молекулно тегло от 35 до 115 kDa. Някои димерни и тетрамерни ензими са съставени от два вида субединици. Няма ясна корелация между размера на ензимната молекула или естеството на нейната структура на субединицата и специфичността.
Специфичността на ензима се определя от неговото силно свързване с акцепторния край на tRNA, DU региона и променливата бримка. Някои ензими изглежда не разпознават антикодоновия триплет и катализират реакцията на аминоацетилиране, дори когато антикодонът е променен. Въпреки това, някои ензими показват намалена активност по отношение на такива модифицирани tRNAs и добавят грешна аминокиселина при заместване на антикодона.
70-90n | вторична страница - детелина | CCA 3" const за всички tRNA |
наличието на тимин, псевдоуридин-пси, дигируридин DGU в D-примката - защита срещу рибонуклеази? дълголетни | Разнообразие от първични структури на tRNA - 61 + 1 - по броя на кодоните + формилметионин tRNA, антикодонът на котката е същият като този на метиониновата tRNA. Разнообразие от третични структури - 20 (според броя на аминокиселините)
Има два вида tRNA свързващи метионин tRNAFMet и tRNAMMet при прокариотите и tRNAIMet и tRNAMMet при еукариоти. Метионинът се добавя към всяка tRNA, като се използва подходящ синтез на аминоацил-tRNA. метионинът, прикрепен към tRNAFMet и tRNAIMet, се образува от ензима метионил-tRNA-трансформилаза до Fmet-tRNAFMet. тРНК, натоварени с формилметионин, разпознават иницииращия кодон AUG.
литература:
За съжаление няма библиография.
Учебник. Въпреки факта, че tRNA е много по-малка, една история за нейната структура, характеристики и функциониране заслужава отделна глава.
И така, tRNA е „адаптер“, който разпознава трибуквената последователност на генетичния код в единия край, съпоставяйки го с единствената съответна аминокиселина, фиксирана в другия край на tRNA. В края на трансферната РНК, която докосва информационната РНК, има 3 нуклеотида, които образуват антикодон. Само ако антикодонът е комплементарен на иРНК региона, трансферната РНК може да се присъедини към него. Но дори и в този случай тРНК не може да се присъедини сама към иРНК, тя се нуждае от помощта на рибозомата, която е мястото на тяхното взаимодействие, както и от активен участник в транслацията. Например рибозомата създава връзки между аминокиселините, донесени от tRNA, образувайки протеинова верига.
Структурните особености на tRNA се определят от генетичния код, тоест правилата за конструиране на протеин според ген, който чете трансферната РНК. Този код работи във всяко живо същество на Земята: създаването на вирус е написано със същите трибуквени кодони, които се използват за написване на "инструкциите за сглобяване" на делфин. Експериментално е потвърдено, че гените на едно живо същество, поставени в клетката на друго, са перфектно копирани и преведени в протеини, които са неразличими от гените, възпроизвеждащи се в клетките на гостоприемника. Еднородността на генетичния код е в основата на производството на модифицирана E. coli от колонии от инсулин и много други човешки ензими, които се използват като лекарства за хора, чиито тела не са в състояние да ги произвеждат или произвеждат недостатъчно. Въпреки очевидната разлика между хората и E. coli, човешките протеини лесно се създават от човешки чертежи с помощта на копирна машина на E. coli. Не е изненадващо, че трансферните РНК на различни същества се различават много малко.

Всеки кодон от този списък, с изключение на три стоп кодони, сигнализиращ за завършване на транслацията, трябва да бъде разпознат от трансферната РНК. Разпознаването се извършва чрез прикрепване на антикодон към информационна РНК, която може да се свърже само с един кодон от списъка, така че tRNA може да разпознае само един кодон. Това означава, че има поне 61 вида от тези молекули в клетката. Всъщност има дори повече от тях, тъй като в някои ситуации за разчитане на информационната РНК не е достатъчно само да имате правилния антикодон: необходими са други условия, в съответствие с които се създава специална, модифицирана tRNA.
На пръв поглед такова разнообразие от тРНК би трябвало значително да усложни процеса на транслация: в края на краищата всяка от тези молекули ще проверява матричния РНК кодон, заместен от рибозомата, за съответствие с неговия антикодон - изглежда, че толкова много безсмислена механична работа , толкова много загубено време и енергия. Но в резултат на еволюцията са се формирали и клетъчни механизми, които предотвратяват този проблем. Например, количеството tRNA на всеки вид в клетката съответства на това колко често аминокиселината, пренасяна от този вид, се намира в протеините, които се изграждат. Има аминокиселини, които рядко се използват от клетката, а има и такива, които се използват често, и ако броят на тРНК, които ги носят, беше същият, това би усложнило значително сглобяването на протеините. Следователно, има малко "редки" аминокиселини и съответните им tRNAs в клетката, докато често срещаните аминокиселини се произвеждат в големи количества.

С такова разнообразие от tRNA молекули всички те са много сходни, следователно, като се има предвид тяхната структура и функции, ще изучаваме основно характеристиките, общи за всички видове. Когато погледнете 3D оформлението на tRNA, то изглежда като плътна купчина атоми. Изглежда невероятно, че тази сложно навита молекула е резултат от сгъването на дълга верига от нуклеотиди, но така се образува.
Възможно е да се проследят етапите на неговото формиране, като се започне от първия: съставянето на нуклеотидна последователност от РНК полимераза в съответствие с гена, съдържащ информация за тази трансферна РНК. Нарича се редът, в който тези нуклеотиди следват един друг и техният брой първична структура на тРНК. Оказва се, че това е първичната структура на tRNA, която е кодирана в гена, прочетен от РНК полимераза. Като цяло, първичната структура е последователност от относително прости молекули от същия тип, от които е съставена по-сложна, нагъната полимерна молекула. Например, първичната структура на протеиновата молекула е простата последователност от съставните й аминокиселини.

Никаква верига от нуклеотиди не може да бъде в разгънато състояние в клетка, просто разпъната в линия. В краищата на нуклеотидите има твърде много положително и отрицателно заредени части, които лесно образуват водородни връзки помежду си. Как се образуват едни и същи връзки между нуклеотидите на две ДНК молекули, свързвайки ги в двойна спирала, е описано в и можете да навлезете в подробности за водородните връзки. Водородните връзки са по-малко силни от връзките между атомите в молекулите, но са достатъчни, за да усучат сложно нишката на tRNA и да я задържат в това положение. Отначало тези връзки се образуват само между някои нуклеотиди, сгъвайки тРНК във форма на листа от детелина. Резултатът от това първоначално сгъване се нарича вторична структура тРНК. Диаграмата вляво показва, че само някои нуклеотиди са свързани с водородни връзки, докато други остават несдвоени, образувайки пръстени и бримки. Разликите между вторичната структура на различните видове tRNA се дължат на различията в тяхната първична структура. Това се проявява в различни дължини на "листата на детелина" или "стъблото" поради различната дължина на първоначалната верига от нуклеотиди.
Друга разлика в първичната структура на различните tRNAs е, че само в някои позиции те имат едни и същи нуклеотиди (в диаграмата по-горе те са маркирани с първите букви на техните имена), докато повечето от нуклеотидите в различните tRNAs се различават един от друг. Схемата по-горе е обща за всички tRNA, така че различните нуклеотиди са маркирани с числа.
Основните функционални части на tRNA са:
=) антикодон, тоест нуклеотидната последователност, която е комплементарна на единичен кодон на информационна РНК, разположена на антикодон фиби
=) край на акцепторакъм които може да се прикрепи аминокиселина. Намира се от противоположната страна на антикодонната фиби.
В действителност нито една тРНК не изглежда така, както изглежда в диаграмата на вторичната структура, защото само някои нуклеотиди се съединяват, за да я образуват, докато останалите остават несдвоени. Поради образуването на водородни връзки между нуклеотиди от различни части на листа от детелина, той се сгъва допълнително в много по-сложна третична структуравъв формата на L. Можете да разберете как точно различните части на вторичната структура са извити, за да образуват третичната структура, като съпоставите цветовете в техните диаграми по-долу. Антикодонната щипка за коса, маркирана в синьо и сиво, остава на дъното (заслужава си да се помни, че това „дъно“ е условно: удобно е да се изобразява tRNA в тази пространствена ориентация в схемите за транслация на протеини), а акцепторният край (жълт) е огънат настрани.

Ето как изглежда tRNA, когато е готова да прикачи аминокиселина. tRNA не може да се комбинира с аминокиселината самостоятелно, това изисква участието на специален ензим: аминоацил-тРНК синтетази. Броят на видовете синтетаза в клетката съвпада с броя на типовете tRNA.
Еднаквостта на формата на всички видове тРНК е необходима, за да може рибозомата да разпознае всеки от тях, да улесни свързването им с иРНК и да се премести в себе си от едно място на друго. Ако различните типове tRNA се различават значително един от друг, това би направило работата на рибозомата изключително трудна, намалявайки критично скоростта на протеиновия синтез. Така естественият подбор има за цел да направи тРНК подобни една на друга. Но в същото време има и друг фактор, който изисква наличието на забележими разлики между различните видове tRNA: в края на краищата е необходимо да се разпознае всеки тип и да се прикрепи към него единствената съответна аминокиселина. Очевидно тези разлики трябва да бъдат забележими, но не твърде значителни, така че работата по разпознаването на видовете tRNA да се превърне в процес на бижута. И точно това се осъществява от аминоацил-тРНК синтетазите: всяка от тях може да се свърже само с една от 20-те аминокиселини и да я прикрепи точно към онези видове тРНК, които съответстват на тази аминокиселина. От таблицата с генетичния код може да се види, че всяка аминокиселина е кодирана от няколко нуклеотидни последователности, следователно, например, всичките четири tRNA с антикодони CGA, CGG, CGU и CGC ще бъдат разпознати от една и съща синтетаза, която свързва аланин на тях. Такива tRNA, обработени от една синтетаза, се наричат свързани.
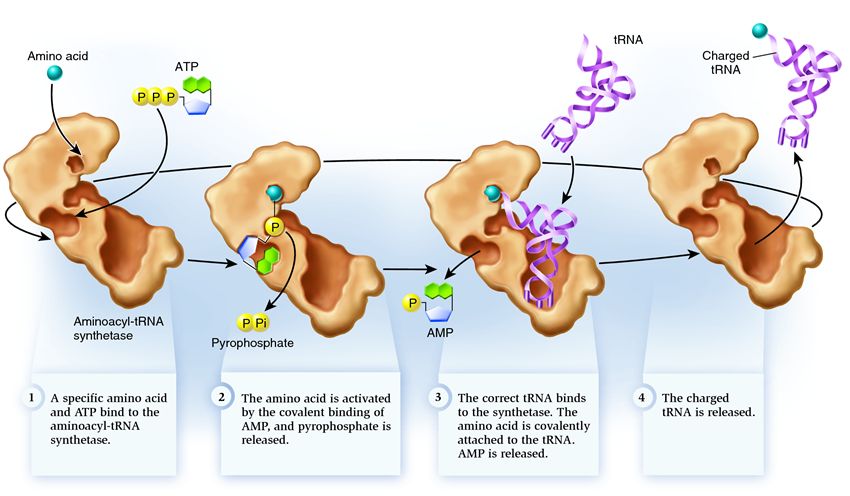
Синтетазата принадлежи към група ензими, чиято функция е да се свързват с отделно съществуващи молекули и да ги комбинират в едно:
1 . синтетазата свързва аминокиселина и АТФ молекула. Две фосфатни групи се отделят от АТФ, освобождавайки енергията, необходима за следните дейности. Аденозин монофосфатът (АМФ), останал от разрушената молекула, се прикрепя към аминокиселината, подготвяйки я за свързване с акцепторната фиба.
2 . синтетазата прикрепя към себе си една от свързаните tRNA, съответстваща на тази аминокиселина.

На този етап се проверява съответствието на трансферната РНК със синтетазата. Има няколко начина за разпознаване и всяка синтетаза има уникална комбинация от тях. Най-малко един антикодон нуклеотид участва във взаимодействието между синтетазата и tRNA. Акцепторната шпилка също трябва да бъде проверена: определя се наличието на специфични нуклеотиди върху нея, които са общи за свързани тРНК, съответстващи на желаната аминокиселина. Нуклеотидите от други части на tRNA също могат да участват в съвпадението чрез свързване към определени места на синтетаза. Грешната tRNA може да съвпадне с желаната по някакъв начин, но поради непълно съвпадение тя ще се присъедини към синтетазата бавно и свободно, като лесно ще падне. И правилната tRNA ще се придържа към синтетазата бързо и здраво, в резултат на което структурата на синтетазата се променя, започвайки процеса аминоацилиране , тоест свързването на аминокиселина към tRNA.

3 . аминоацилирането се състои в заместване на молекулата AMP, прикрепена към аминокиселината, с tRNA молекула. След това заместване AMP напуска синтетазата и tRNA се задържа за последна проверка на аминокиселината. Ако прикрепената аминокиселина бъде разпозната като неправилна, тя ще се отдели от тРНК, мястото на аминокиселината в синтетазата ще бъде празно и друга молекула може да се присъедини там. Новата аминокиселина ще премине през етапите на свързване с АТФ и tRNA и също ще бъде тествана. Ако не бяха допуснати грешки, заредената с аминокиселина tRNA се освобождава: тя е готова да играе ролята си в транслацията на протеин. И синтетазата е готова да прикрепи нови аминокиселини и тРНК и цикълът ще започне отначало.
Много зависи от правилната работа на аминоацил-тРНК синтетазата: ако на този етап възникне неуспех, тогава грешната аминокиселина ще бъде прикрепена към тРНК. И ще бъде вграден в протеина, растящ върху рибозомата, тъй като тРНК и рибозомата нямат функцията да проверяват съответствието на кодона и аминокиселината. Последиците от грешката могат да бъдат незначителни или катастрофални и чрез естествен подбор същества с ензими, които нямат функцията на такива проверки, са били изместени от по-адаптивни такива с различни опции за съвпадение между аминокиселина и тРНК. Следователно в съвременните клетки синтетазата се комбинира с грешната аминокиселина средно в един случай от 50 хиляди, а с грешната tRNA само веднъж на 100 хиляди прикрепвания.

Някои аминокиселини се различават една от друга само с няколко атома. Ако погледнете техните схеми, става очевидно, че вероятността да объркате аргинин с аланин е много по-малка, отколкото да объркате изолевцин с левцин или валин. Следователно всяка синтетаза, която се свързва с една от подобните една на друга аминокиселини, има допълнителни механизми за проверка. Ето пример за такава адаптация в изолевцин-тРНК синтетаза:

Всяка синтаза има синтетичен центърв който аминокиселина е прикрепена към tRNA. Акцепторната шпилка на tRNA, уловена от синтетазата, отива там, както и аминокиселината, която е готова да реагира с нея. Работата на някои синтетази завършва веднага след свързването на аминокиселината и tRNA. Но Ile-tRNA синтетазата има повишен шанс да направи грешки поради съществуването на други изолевциноподобни аминокиселини. Следователно тя също има поправителен център: от името става ясно каква роля играе в процеса на свързване на тРНК и аминокиселини. Фигурата вдясно показва, че позицията на края на фиби за акцептор на tRNA в синтетичния център на Ile-tRNA синтетазата придава на тази фиба неестествено извиване. Синтетазата обаче задържа tRNA в това положение, докато аминокиселината не се прикрепи към нея. След като тази връзка е настъпила, необходимостта от намиране на акцепторната фиба в синтетичния център е изчерпана и tRNA се изправя, като краят си с прикрепената към нея аминокиселина се намира в центъра за корекция.

Разбира се, синтетичният център също играе роля при скрининг на аминокиселини, които не са подходящи за синтетаза. За да влезе в него, молекулата трябва да отговаря на редица условия, включително да има правилния размер. Въпреки факта, че левцинът и изолевцинът съдържат еднакъв брой атоми, поради разликите в пространствената структура, левцинът е по-голям. Следователно, той не може да проникне в синтетичния център, чийто размер съответства на по-компактния изолевцин, и просто отскача от Ile-tRNA синтетазата.
Но валинът, който е най-малката от тези три молекули с подобна атомна структура, лесно заема мястото на изолевцин в синтетичния център и синтетазата го прикрепя към тРНК. В този случай се задейства корекционният център на синтетазата. Ако фиби за изправяне акцептор е правилно зареден и носи изолевцин, тогава той не може да се притисне вътре в центъра за корекция: просто е твърде малък за тази молекула. Така изправената tRNA вече не се държи от нищо и тя се отделя от синтетазата. Но ако валинът е прикрепен към tRNA, той се плъзга в центъра за корекция, като по този начин поддържа tRNA свързана с него в синтетазата. Такъв прекалено дълъг престой на tRNA вътре е сигнал за грешка за синтетазата, променящ нейната пространствена конфигурация. Като резултат:
=) валинът се отделя от tRNA и се отстранява от синтетазата
=) акцепторната фиби се връща към синтетичното място, чакайки прикрепване към аминокиселината
=) синтетазата се свързва с нова аминокиселина, „зарежда“ tRNA с нея и отново проверява дали за това е използван изолевцин.
Подобен механизъм за двойно разпознаване се използва от други синтетази.
Физични и химични свойства на ДНК
Различни фактори, които разрушават водородните връзки (повишаване на температурата над 80 С, промени в рН и йонната сила, действието на уреята и др.) предизвикват денатурация на ДНК, т.е. промяна в пространственото подреждане на ДНК веригите без прекъсване на ковалентни връзки. Двойната спирала на ДНК по време на денатурация е напълно или частично разделена на съставните си вериги. Денатурацията на ДНК е придружена от увеличаване на оптичната абсорбция в UV областта на пуринови и пиримидинови бази. Това явление се нарича хиперхромен ефект . Денатурацията също така намалява високия вискозитет, присъщ на естествените ДНК разтвори. Когато оригиналната двуверижна ДНК структура се възстанови, в резултат на ренатурация, абсорбцията при 260 nm от азотните бази намалява поради тяхното "екраниране". Това явление се нарича хипохромен ефект .
"Развиването" на всяка ДНК в нейните съставни вериги се извършва в рамките на определен температурен диапазон. Средната точка на този интервал се нарича точка на топене. Температурата на топене на ДНК зависи при стандартни условия (определено рН и йонна сила) от съотношението на азотните основи. G-C двойките, съдържащи три водородни връзки, са по-силни, следователно, колкото по-високо е съдържанието на G-C двойки в ДНК, толкова по-висока е точката на топене.
Функции на ДНК. В последователността от нуклеотиди в молекулите на ДНК е кодирана генетична информация. Основните функции на ДНК са, първо, да осигури възпроизвеждането на себе си в поредица от клетъчни поколения и поколения организми, и второ, да осигури синтеза на протеини. Тези функции се дължат на факта, че ДНК молекулите служат като матрица в първия случай за репликация, т.е. копиране на информация в дъщерни ДНК молекули, във втората - за транскрипция, т.е. за прекодиране на информация в структурата на РНК.
Ориз. 5 Крива на топене (денатурация на ДНК)
Комплементарните нишки на ДНК, разделени по време на денатурацията, могат при определени условия да се свържат отново в двойна спирала. Този процес се нарича РЕНАТУРАЦИЯ. Ако денатурацията не е настъпила напълно и поне няколко бази не са загубили взаимодействие чрез водородни връзки, ренатурацията протича много бързо.
Цитоплазмата на клетките съдържа три основни функционални типа РНК. Това са информационни РНК – иРНК, които действат като шаблони за протеинов синтез, рибозомни РНК – рРНК, които действат като структурни компоненти на рибозомите, и трансферни РНК – тРНК, участващи в транслацията (транслацията) на информацията за иРНК в аминокиселинната последователност в протеина.
Таблица 2 показва разликите между ДНК и РНК по отношение на структурата, локализацията в клетката и функциите.
Таблица 2 Разлики между ДНК и РНК
Трансферна РНК, тРНК-рибонуклеинова киселина, чиято функция е да транспортира АА до мястото на протеиновия синтез. Той има типична дължина от 73 до 93 нуклеотида и размер около 5 nm. тРНК също участват пряко в растежа на полипептидната верига, като се присъединяват – като са в комплекс с аминокиселина – към кодона на иРНК и осигуряват конформацията на комплекса, необходима за образуването на нова пептидна връзка. Всяка аминокиселина има своя собствена tRNA. tRNA е едноверижна РНК, но във функционалната си форма има конформация на детелина. АА ковалентно се прикрепя към 3" края на молекулата, използвайки ензима аминоацил-тРНК синтетаза, специфичен за всеки тип тРНК. На място С има антикодон, съответстващ на AA-te. tRNAs се синтезират от обикновена РНК полимераза в случая на прокариоти и от РНК полимераза III в случая на еукариоти Преписите на tRNA гени се подлагат на многоетапна обработка, което води до образуването на пространствена структура, типична за tRNA.
Обработката на tRNA включва 5 ключови стъпки:
отстраняване на 5" лидерната нуклеотидна последователност;
отстраняване на 3'-терминалната последователност;
добавяне на CCA последователност в 3" края;
изрязване на интрони (при еукариоти и археи);
модификации на отделни нуклеотиди.
Транспортирането на tRNA се извършва по Ran-зависим път с участието на транспортния фактор exportin t, който разпознава характерния вторичен и третичен str-ru на зрялата tRNA: къси двуверижни участъци и правилно обработени 5 "- и 3" завършва. Този механизъм гарантира, че само зрели tRNAs се изнасят от ядрото.
62. Транслация - разпознаване на кодон на иРНК
Транслацията е протеинов синтез, осъществяван от рибозоми от аминокиселини върху иРНК (или и РНК) шаблон. Съставните елементи на процеса на транслация: аминокиселини, тРНК, рибозоми, иРНК, ензими за аминоацилиране на тРНК, протеинови транслационни фактори (протеинови фактори на иницииране, удължаване, терминиране - специфични екстрарибозомни протеини, необходими за процесите на транслация), ATP и GTP енергийни източници , магнезиеви йони (стабилизират рибозомната структура). 20 аминокиселини участват в протеиновия синтез. За да може една аминокиселина да „разпознае“ своето място в бъдещата полипептидна верига, тя трябва да се свърже с трансферна РНК (tRNA), която изпълнява функция на адаптер. След това тРНК, която се свързва с аминокиселината, разпознава съответния кодон на иРНК. разпознаване на иРНК кодон:
Взаимодействието кодон-антикодон се основава на принципите на комплементарност и антипаралелизъм:
3'----C - G-A*------5' tRNA антикодон
5'-----G-C-Y*------3' иРНК кодон
Хипотезата за колебание е предложена от Ф. Крик:
3'-базата на тРНК кодона има нестриктно сдвояване с 5'-базата на тРНК антикодона: например Y (mRNA) може да взаимодейства с A и G (tRNA)
Някои tRNAs могат да се сдвоят с повече от един кодон.
63. Характеристика на съставните елементи на процеса на превод.Транслацията (translatio-translation) е процесът на синтез на протеин от аминокиселини върху матрицата на информационната (матрична) РНК (mRNA, mRNA), осъществяван от рибозомата.
Синтезът на протеини е в основата на клетъчния живот. За извършване на този процес в клетките на всички организми има специални органели - рибозоми- рибонуклеопротеинови комплекси, изградени от 2 субединици: голяма и малка. Функцията на рибозомите е да разпознават три букви (три нуклеотида) кодонитРНК, сравнявайки ги със съответните тРНК антикодони, носещи аминокиселинии добавянето на тези аминокиселини към растящата протеинова верига. Движейки се по протежение на молекулата на иРНК, рибозомата синтезира протеин в съответствие с информацията, съдържаща се в молекулата на иРНК.
За разпознаване на AK-t в клетката има специални "адаптери", трансфер на РНК молекули(tRNA). Тези молекули с форма на детелина имат място (антикодон), комплементарно на тРНК кодон, както и друго място, към което е прикрепена аминокиселината, съответстваща на този кодон. Прикрепването на аминокиселини към tRNA се осъществява в енергийно-зависима реакция от ензимите аминоацил-тРНК синтетази и получената молекула се нарича аминоацил-тРНК. По този начин специфичността на транслацията се определя от взаимодействието между тРНК кодона и тРНК антикодона, както и специфичността на аминоацил-тРНК синтетазите, които прикрепват аминокиселини стриктно към съответните им tRNAs (например, GGU кодонът ще съответства на tRNA, съдържаща CCA антикодона и само AK глицин).
прокариотна рибозома
5S и 23S рРНК 16S рРНК
34 катерици 21 катерици
Прокариотните рибозоми имат константа на утаяване 70S, поради което се наричат 70S частици. Те са изградени от две различни субединици: 30S и 50S субединици. Всяка субединица е комплекс от рРНК и рибозомни протеини.
30S частицата съдържа една 16S rRNA молекула и в повечето случаи една протеинова молекула от повече от 20 вида (21) . 50S субединицата се състои от две rRNA молекули (23S и 5S). Състои се от повече от 30 различни протеина (34), също представени, като правило, от едно копие. Повечето от рибозомните протеини изпълняват структурна функция.
еукариотна рибозома
5S; 5,8S и 28S рРНК 18S рРНК
най-малко 50 протеина най-малко 33 протеина
Рибозомата се състои от големи и малки субединици. Основата на структурата на всяка субединица е сложно нагъната рРНК. Рибозомните протеини бяха прикрепени към скелета на рРНК.
Коефициентът на утаяване на пълната еукариотна рибозома е около 80 единици на Сведберг (80S), а коефициентът на утаяване на нейните субчастици е 40S и 60S.
По-малката 40S субединица се състои от една 18S rRNA молекула и 30-40 протеинови молекули. Голямата субединица 60S съдържа три вида рРНК с коефициенти на утаяване от 5S, 5.8S и 28S и 40-50 протеини (например рибозомите на хепатоцитите на плъх включват 49 протеина).
Функционални области на рибозомите
P - пептидилов сайт за пептидил tRNA
A - аминоацилно място за аминоацил tRNA
E - място за освобождаване на tRNA от рибозомата
Рибозомата съдържа 2 функционални места за взаимодействие с tRNA: аминоацил (акцептор) и пептидил (донор). Аминоацил-тРНК влиза в акцепторното място на рибозомата и взаимодейства, за да образува водородни връзки между кодон и антикодон триплети. След образуването на водородни връзки, системата придвижва 1 кодон и се озовава в донорното място. В същото време в освободеното акцепторно място се появява нов кодон и към него се прикрепя съответната аминоацил-t-РНК.
Рибозоми: структура, функция
Рибозомите са цитоплазмените центрове на биосинтеза на протеини. Те се състоят от големи и малки субединици, които се различават по коефициенти на утаяване (скорост на утаяване по време на центрофугиране), изразени в единици на Svedberg - S.
Рибозомите присъстват както в еукариотните, така и в прокариотните клетки, тъй като те играят важна роля в протеинова биосинтеза.Всяка клетка съдържа десетки, стотици хиляди (до няколко милиона) от тези малки заоблени органели. Това е закръглена рибонуклеопротеинова частица. Диаметърът му е 20-30 nm. Рибозомата се състои от големи и малки субединици, различаващи се по коефициенти на утаяване (скорост на утаяване по време на центрофугиране), изразени в единици на Сведберг - S. Тези субединици се комбинират в присъствието на верига от m-RNA (матрична, или информационна, РНК). Комплекс от група рибозоми, обединени от една молекула иРНК като низ от мъниста, се нарича полизома. Тези структури са или свободно разположени в цитоплазмата, или прикрепени към мембраните на гранулирания ER (и в двата случая протеиновият синтез протича активно върху тях).
Полизомите на гранулирания ЕР образуват протеини, които се отделят от клетката и се използват за нуждите на целия организъм (например храносмилателни ензими, протеини на човешката кърма). Освен това рибозомите присъстват на вътрешната повърхност на митохондриалните мембрани, където също участват активно в синтеза на протеинови молекули.
Трансферната РНК (tRNA) играе важна роля в процеса на използване на наследствена информация от клетката. Доставяйки необходимите аминокиселини до мястото на сглобяване на пептидните вериги, tRNA действа като транслационен медиатор.
tRNA молекулите са полинуклеотидни вериги, синтезирани върху специфични ДНК последователности. Те се състоят от относително малък брой нуклеотиди -75-95. В резултат на комплементарното свързване на бази, които се намират в различни части на полинуклеотидната верига на tRNA, тя придобива структура, наподобяваща по форма лист на детелина (фиг. 3.26).
Ориз. 3.26. Структурата на типична tRNA молекула.
Той има четири основни части, които изпълняват различни функции. акцептор"Стъбла" се образува от две допълващи се свързани крайни части на tRNA. Състои се от седем базови двойки. 3'-краят на това стебло е малко по-дълъг и образува едноверижен регион, който завършва с CCA последователност със свободна ОН група. Към тази цел е прикрепена транспортируема аминокиселина. Останалите три клона са комплементарни сдвоени нуклеотидни последователности, които завършват в несдвоени участъци, които образуват бримки. Средата на тези разклонения - антикодон - се състои от пет двойки нуклеотиди и съдържа антикодон в центъра на своята бримка. Антикодонът е три нуклеотида, комплементарни на кодона на иРНК, който кодира аминокиселината, транспортирана от тази tRNA до мястото на пептидния синтез.
Между акцепторния и антикодонния клон има два странични клона. В своите бримки те съдържат модифицирани бази - дихидроуридин (D-контур) и TψC триплет, където \y е псевдоуриаин (T^C-контур).
Между разклоненията на атикодон и T^C има допълнителен контур, който включва от 3-5 до 13-21 нуклеотида.
Като цяло различните видове tRNA се характеризират с определено постоянство на нуклеотидната последователност, която най-често се състои от 76 нуклеотида. Вариацията в техния брой се дължи главно на промяната в броя на нуклеотидите в допълнителната бримка. Комплементарните региони, които поддържат структурата на tRNA, обикновено са запазени. Първичната структура на tRNA, определена от последователността на нуклеотидите, образува вторичната структура на tRNA, която има формата на лист от детелина. От своя страна вторичната структура причинява триизмерна третична структура, която се характеризира с образуването на две перпендикулярни двойни спирали (фиг. 3.27). Единият от тях се образува от акцепторните и TψC разклонения, другият от антикодона и D разклонения.
В края на едната от двойните спирали е транспортираната аминокиселина, в края на другата е антикодонът. Тези зони са най-отдалечени една от друга. Стабилността на третичната структура на tRNA се поддържа поради появата на допълнителни водородни връзки между основите на полинуклеотидната верига, разположени в различни части от нея, но пространствено близки в третичната структура.
Различните видове tRNAs имат сходна третична структура, макар и с някои вариации.
Ориз. 3.27. Пространствена организация на tRNA:
I - вторичната структура на tRNA под формата на "лист от детелина", определена от нейната първична структура (последователността на нуклеотидите във веригата);
II - двуизмерна проекция на третичната структура на tRNA;
III - разположение на молекулата tRNA в пространството
ПРИЛОЖЕНИЕ (ако някой не разбира това)
Светкавични зъби - нуклеотиди (Аденин-Тимин/Урацил/, Гуанин-Цитазин). Всички мълнии са ДНК.

За да прехвърлите информация от ДНК, трябва да разкъсате 2 нишки. Връзката между A-T и G-C е водород, поради което лесно се разрушава от ензима Helicase:

За да предотвратите образуването на възли (Като пример, усуках кърпа):



Топоизомераза разрязва една верига от ДНК в началото на репликацията, така че веригата да не се усуква.

Когато една нишка е свободна, втората може лесно да се върти около оста си, като по този начин облекчава напрежението по време на "развиване". Възлите не се появяват, енергията се пести.
След това е необходим РНК праймер, за да започне събирането на РНК. Протеин, който сглобява иРНК, не може просто да сглоби първия нуклеотид, той се нуждае от парче РНК, за да започне (това е написано подробно там, ще го напиша по-късно). Това парче се нарича РНК праймер. И този протеин вече прикрепя първия нуклеотид към него.


При описването на структурата на нуклеиновите киселини се вземат предвид различни нива на организация на макромолекулите: първична и вторична структура.
Първичната структура на нуклеиновите киселини е нуклеотидният състав и определена последователност от нуклеотидни единици в полимерната верига.
Вторична структура на РНК. Молекулата на рибонуклеинова киселина е изградена от единична полинуклеотидна верига.
Вторична структура на РНК
Отделни участъци от веригата на РНК образуват спираловидни бримки - "шпильки", дължащи се на водородни връзки между комплементарните азотни бази A-U и G-C. Секциите на РНК веригата в такива спираловидни структури са антипаралелни, но не винаги напълно комплементарни; те съдържат несдвоени нуклеотидни остатъци или дори едноверижни бримки, които не се вписват в двойната спирала. Наличието на спираловидни участъци е характерно за всички видове РНК.
Основната роля на РНК е прякото участие в биосинтеза на протеини.
Известни са три типа клетъчна РНК, които се различават по местоположението си в клетката, състава, размера и свойствата, които определят тяхната специфична роля в образуването на протеинови макромолекули:
- информационните (матрични) РНК предават кодирана в ДНК информация за структурата на протеина от клетъчното ядро до рибозомите, където се осъществява протеиновия синтез; първичната структура на всички иРНК, независимо от уникалността на тяхната кодираща последователност, има една и съща структура на 5'- и 3'-края.
И така, в 5'-края има модифициран нуклеотид 7-метилгуанозин-5'-трифосфат (cap). Няколко десетки нуклеотида отделят капачката от иницииращия кодон, обикновено триплета -AUG-. Кодиращата област е последвана от един от крайните кодони -UGA-, -UUA-, -UAG-. В 3' края на повечето иРНК има нуклеотидна последователност от 100-200 аденозин монофосфатни остатъци.
- трансферни РНК събират аминокиселини в цитоплазмата на клетката и ги пренасят в рибозомата; РНК молекулите от този тип "научават" от съответните участъци на веригата на информационната РНК кои аминокиселини трябва да участват в протеиновия синтез.
Пространствената структура на всяка tRNA, независимо от разликите в нуклеотидната последователност, се описва от универсалния модел на листата на детелина. Всяка tRNA молекула има верижни участъци, които не участват в образуването на водородни връзки между нуклеотидни остатъци.
Те включват по-специално мястото, отговорно за свързването с аминокиселината в 3'-края на молекулата и антикодона, специфичен триплет от нуклеотиди, който взаимодейства комплементарно с кодона на иРНК.
- рибозомната РНК осигурява протеинов синтез на определена структура, четейки информация от информационната (матрична) РНК. рРНК образуват комплекси с протеини, наречени рибозоми.
Всяка рибозома се състои от две субединици - малка (40S) и голяма (60S). Рибозомните субединици се различават не само по набора от рРНК, но и по броя и структурата на протеините.
Дата на публикуване: 2015-02-03; Прочетено: 2729 | Нарушаване на авторски права на страницата

РНК е полимер, чиито мономери са рибонуклеотиди.
За разлика от ДНК, РНК се образува не от две, а от една полинуклеотидна верига (изключение - някои РНК-съдържащи вируси имат двуверижна РНК). РНК нуклеотидите са способни да образуват водородни връзки помежду си. РНК веригите са много по-къси от ДНК веригите.
РНК мономерът - нуклеотид (рибонуклеотид) - се състои от остатъци от три вещества: 1) азотна основа, 2) петвъглероден монозахарид (пентоза) и 3) фосфорна киселина. Азотните бази на РНК също принадлежат към класовете пиримидини и пурини.
Пиримидинови бази на РНК - урацил, цитозин, пуринови основи - аденин и гуанин.
31. Видове РНК и особености на нейната структура
РНК нуклеотидният монозахарид е представен от рибоза.
Има три вида РНК: 1) информационна (матрична) РНК - иРНК (тРНК), 2) трансферна РНК - тРНК, 3) рибозомна РНК - рРНК.
Всички видове РНК са неразклонени полинуклеотиди, имат специфична пространствена конформация и участват в процесите на протеинов синтез.
Информацията за структурата на всички видове РНК се съхранява в ДНК. Процесът на синтез на РНК върху ДНК шаблон обикновено се нарича транскрипция.

Трансферните РНК обикновено съдържат 76 (от 75 до 95) нуклеотида; молекулно тегло - 25 000–30 000.
tRNA представлява около 10% от общото съдържание на РНК в клетката. Функции на tRNA: 1) транспорт на аминокиселини до мястото на синтеза на протеини, до рибозомите, 2) транслационен медиатор. В клетката се намират около 40 вида tRNA, всеки от тях има характерна само за нея нуклеотидна последователност. В същото време всички tRNAs имат няколко вътрешномолекулни комплементарни области, поради което tRNAs придобиват конформация, наподобяваща по форма лист на детелина.
Всяка тРНК има бримка за контакт с рибозомата (1), антикодонна бримка (2), бримка за контакт с ензима (3), акцепторно стъбло (4) и антикодон (5). Аминокиселината е прикрепена към 3' края на акцепторното стъбло. Антикодон - три нуклеотида, които "разпознават" тРНК кодон.
Трябва да се подчертае, че определена tRNA може да транспортира строго определена аминокиселина, съответстваща на нейния антикодон. Специфичността на връзката на аминокиселините и tRNA се постига благодарение на свойствата на ензима аминоацил-тРНК синтетаза.
Рибозомните РНК съдържат 3000–5000 нуклеотида; молекулно тегло - 1 000 000–1 500 000.
rRNA представлява 80-85% от общото съдържание на РНК в клетката. В комплекс с рибозомните протеини, рРНК образува рибозоми - органели, които осъществяват протеинов синтез. В еукариотните клетки синтезът на рРНК се извършва в ядрото. Функции на рРНК: 1) необходим структурен компонент на рибозомите и по този начин осигуряващ функционирането на рибозомите; 2) осигуряване на взаимодействието на рибозомата и tRNA; 3) първоначалното свързване на рибозомата и инициаторния кодон на иРНК и определяне на рамката за четене, 4) образуване на активния център на рибозомата.
Информационните РНК са разнообразни по нуклеотидно съдържание и молекулно тегло (от 50 000 до 4 000 000).
Делът на иРНК представлява до 5% от общото съдържание на РНК в клетката. Функции на иРНК: 1) пренос на генетична информация от ДНК към рибозоми, 2) матрица за синтез на протеинова молекула, 3) определяне на аминокиселинната последователност на първичната структура на протеиновата молекула.
Прочетете също
РНК е полимер, чиито мономери са нуклеотиди.
Трите азотни бази са същите като в ДНК (аденин, гуанин, цитозин); четвъртият - урацил - присъства в молекулата на РНК вместо тимин. РНК нуклеотидите съдържат рибоза вместо дезоксирибоза. Във веригата на РНК...
три основни типа РНК: информационни(тРНК), или матрица(иРНК), рибозомни(рРНК) и транспорт(tRNA). Те се различават по молекулен размер и функция. Всички видове РНК се синтезират върху ДНК с участието на ензими - РНК полимерази. Информационната РНК съставлява 2-3% от цялата клетъчна РНК, рибозомната - 80-85, транспортната - около 15%.
иРНК.
той чете наследствена информация от ДНК сегмент и под формата на копирана последователност от азотни бази я прехвърля към рибозомите, където се синтезира определен протеин. Всяка от молекулите на иРНК в реда на нуклеотиди и по размер съответства на гена в ДНК, от който е транскрибирана. Средно иРНК съдържа 1500 нуклеотида (75-3000). Всеки триплет (три нуклеотида) върху иРНК се нарича кодон.От кодона зависи коя аминокиселина ще се появи на дадено място по време на протеиновия синтез.
(тРНК)има относително ниско молекулно тегло от около 24-29 хиляди.
D и съдържа от 75 до 90 нуклеотида в молекулата. До 10% от всички тРНК нуклеотиди са второстепенни бази, което, очевидно, го предпазва от действието на хидролитичните ензими.Ролята на тРНК е, че те пренасят аминокиселини в рибозомите и участват в процеса на синтеза на протеини. Всяка аминокиселина се прикрепя към специфична tRNA. Редица аминокиселини имат повече от една tRNA. Към днешна дата са открити повече от 60 tRNA, които се различават по своята първична структура (базова последователност).
Вторичната структура на всички tRNAs е представена под формата на лист от детелина с двуверижно стъбло и три едноверижни). В края на една от веригите има акцепторно място - триплета CCA, към чийто аденин е прикрепена специфична аминокиселина.
(рРНК). Те съдържат 120-3100 нуклеотида. Рибозомната РНК се натрупва в ядрото, в нуклеолите.
Рибозомните протеини се транспортират до нуклеолите от цитоплазмата и там става спонтанно образуване на рибозомни субчастици чрез комбиниране на протеини със съответната рРНК. Субчастиците на рибозомата се транспортират заедно или поотделно през порите на ядрената мембрана в цитоплазмата. Рибозомиса органели с размер 20-30 nm.
Те са изградени от две субчастици с различни размери и форми. На определени етапи от протеиновия синтез в клетката рибозомите се разделят на субчастици.
Рибозомната РНК служи като рамка за рибозомите и улеснява първоначалното свързване на иРНК с рибозомата по време на биосинтеза на протеин.
Въпрос 6 Връзките, които образуват първичните и вторичните структури на ДНК и РНК. Видове РНК
Генетичният код е начин за кодиране на аминокиселинната последователност на протеините с помощта на последователност от нуклеотиди, характерна за всички живи организми.
Свойства: 1) генетичен код тройка(всяка аминокиселина е кодирана от три нуклеотида); 2) не препокриващи се(съседните триплети нямат общи нуклеотиди); 3) изродени(с изключение на метионин и триптофан, всички аминокиселини имат повече от един кодон); 4) универсален(предимно еднакви за всички живи организми); 5) в кодони за една аминокиселина първите два нуклеотида обикновено са еднакви, а третият варира; 6) има линеен ред на четене и се характеризира с колинеарност,Т.
д. съвпадението на реда на кодоните в иРНК с реда на аминокиселините в синтезираната полипептидна верига.
Дата на публикуване: 2014-12-08; Прочетено: 11268 | Нарушаване на авторски права на страницата
studopedia.org - Studopedia.Org - 2014-2018. (0,001 s) ...
Цитоплазмата на клетките съдържа три основни функционални типа РНК:
- информационна РНК (мРНК), която действа като шаблони за протеинов синтез;
- рибозомна РНК (рРНК), действаща като структурни компоненти на рибозомите;
- трансферни РНК (tRNAs), участващи в транслацията (транслацията) на информацията за иРНК в аминокиселинната последователност на протеинова молекула.
В ядрото на клетките се намира ядрена РНК, която съставлява от 4 до 10% от общата клетъчна РНК.
По-голямата част от ядрената РНК е представена от високомолекулни прекурсори на рибозомна и трансферна РНК. Прекурсорите на рРНК с високо молекулно тегло (28 S, 18 S и 5 S РНК) са локализирани главно в ядрото.
РНК е основният генетичен материал в някои животински и растителни вируси (геномна РНК). Повечето РНК вируси се характеризират с обратна транскрипция на техния РНК геном, насочена от обратна транскриптаза.
Всички рибонуклеинови киселини са полимери на рибонуклеотиди, свързани, както в молекула на ДНК, чрез 3′,5′-фосфородиестерни връзки.
За разлика от ДНК, която има двуверижна структура, РНК е едноверижна линейна полимерна молекула.
структура на иРНК. иРНК е най-хетерогенният клас РНК по отношение на размера и стабилността.
структура на тРНК.
Трансферните РНК действат като медиатори (адаптери) по време на транслацията на иРНК. Те представляват приблизително 15% от общата клетъчна РНК. Всяка от 20-те протеиногенни аминокиселини има своя собствена tRNA. За някои аминокиселини, кодирани от два или повече кодона, има няколко tRNA.
tRNAs са относително малки едноверижни молекули, състоящи се от 70-93 нуклеотида. Молекулното им тегло е (2,4-3,1) .104 kDa.
Вторичната структура на tRNA се образува поради образуването на максимален брой водородни връзки между вътрешномолекулни комплементарни двойки азотни основи.
В резултат на образуването на тези връзки, полинуклеотидната верига на tRNA се усуква с образуването на спираловидни разклонения, завършващи с бримки от несдвоени нуклеотиди. Пространственото изображение на вторичните структури на всички tRNAs има формата на детелина.
Четири задължителни клона се разграничават в "детелината", по-дълги tRNAs, освен това съдържат кратък пети (допълнителен) клон.
Адапторната функция на tRNA се осигурява от акцепторно разклонение, към чийто 3'-край е прикрепен аминокиселинен остатък чрез етерна връзка и антикодонов клон срещу акцепторния клон, в горната част на който има бримка, съдържаща антикодон.
Антикодонът е специфичен триплет от нуклеотиди, който е комплементарен в антипаралелна посока на кодона на иРНК, кодиращ съответната аминокиселина.
Т-клонът, носещ псевдоуридиновата бримка (TyC-loop), осигурява взаимодействието на tRNA с рибозомите.
D-клонът, носещ дехидроуридиновата бримка, осигурява взаимодействието на tRNA със съответната аминоацил-tRNA синтетаза.

Вторична структура на тРНК
Функциите на петия допълнителен клон все още са слабо разбрани; най-вероятно той изравнява дължината на различните tRNA молекули.
Третичната структура на тРНК е много компактна и се образува чрез обединяване на отделни клони на листата на детелина поради допълнителни водородни връзки, за да се образува L-образна структура на "лакътя".
Транспортна РНК, структура и функционален механизъм.
В този случай акцепторното рамо, което свързва аминокиселината, се намира в единия край на молекулата, а антикодонът е в другия.

Третична структура на tRNA (според A.S. Spirin)
Структурата на рРНК и рибозомите. Рибозомните РНК образуват гръбнака, към който се свързват специфични протеини, за да образуват рибозоми. Рибозомите са нуклеопротеинови органели, които осигуряват синтеза на протеин от иРНК.
Броят на рибозомите в клетката е много голям: от 104 при прокариотите до 106 при еукариотите. Рибозомите са локализирани главно в цитоплазмата, в еукариотите, освен това в ядрото, в митохондриалния матрикс и в стромата на хлоропластите. Рибозомите се състоят от две субчастици: голяма и малка. По размер и молекулно тегло всички изследвани рибозоми са разделени на 3 групи - 70S рибозоми на прокариоти (S-коефициент на утаяване), състоящи се от малки 30S и големи 50S субчастици; 80S еукариотни рибозоми, състоящи се от 40S малки и 60S големи субединици.
Малката субединица от 80S рибозоми се образува от една rRNA молекула (18S) и 33 молекули от различни протеини.
Голямата субединица се образува от три rRNA молекули (5S, 5.8S и 28S) и приблизително 50 протеина.
Вторичната структура на рРНК се образува поради къси двуверижни участъци от молекулата - фиби (около 2/3 от рРНК), 1/3 - е представена от едноверижни участъци, богати на пуринови нуклеотиди.
Социални бутони за Joomla
Протеините са в основата на живота. Техните функции в клетката са много разнообразни. Протеините обаче "не могат" да се възпроизвеждат. И цялата информация за структурата на протеините се съдържа в гените (ДНК).
При висшите организми протеините се синтезират в цитоплазмата на клетката, а ДНК е скрита зад обвивката на ядрото. Следователно ДНК не може директно да служи като шаблон за протеинов синтез. Тази роля изпълнява друга нуклеинова киселина - РНК.
РНК молекулата е неразклонен полинуклеотид с третична структура.
Образува се от една полинуклеотидна верига и въпреки че комплементарните нуклеотиди, включени в нея, също са способни да образуват водородни връзки помежду си, тези връзки възникват между нуклеотидите на една верига. РНК веригите са много по-къси от ДНК веригите. Ако съдържанието на ДНК в клетката е относително постоянно, тогава съдържанието на РНК варира значително. Най-голямо количество РНК в клетките се наблюдава по време на протеиновия синтез.
РНК играе основна роля в предаването и прилагането на наследствена информация.
В съответствие с функцията и структурните особености се разграничават няколко класа клетъчна РНК.
Има три основни класа клетъчна РНК.
- Информационна (mRNA) или матрична (mRNA). Неговите молекули са най-разнообразни по отношение на размери, молекулно тегло (от 0,05x106 до 4x106) и стабилност.
Те съставляват около 2% от общото количество РНК в клетката. Всички иРНК са носители на генетична информация от ядрото до цитоплазмата, до мястото на протеиновия синтез. Те служат като матрица (работен чертеж) за синтеза на протеинова молекула, тъй като определят аминокиселинната последователност (първична структура) на протеиновата молекула.

- Рибозомна РНК (рРНК).
Те съставляват 80-85% от общото съдържание на РНК в клетката.
31. Структурата на РНК. Типове РНК, структурни особености и функции. Вторична структура на тРНК
Рибозомната РНК се състои от 3-5 хиляди нуклеотида. Синтезира се в нуклеолите на ядрото. В комплекс с рибозомните протеини рРНК образува рибозоми - органели, върху които се сглобяват протеинови молекули. Основното значение на рРНК е, че осигурява първоначалното свързване на иРНК и рибозома и образува активния център на рибозомата, в който се образуват пептидни връзки между аминокиселини по време на синтеза на полипептидната верига.
- Трансферни РНК (tRNAs).
tRNA молекулите обикновено съдържат 75-86 нуклеотида. Молекулното тегло на молекулите на тРНК е около 25 хил. Молекулите на тРНК играят ролята на посредници в протеиновата биосинтеза – доставят аминокиселини до мястото на протеиновия синтез, тоест до рибозомите. Клетката съдържа повече от 30 вида tRNA. Всеки тип tRNA има своя уникална нуклеотидна последователност.
Въпреки това, всички молекули имат няколко вътрешномолекулни комплементарни области, поради наличието на които всички tRNAs имат третична структура, наподобяваща по форма лист на детелина.
Вторична структура на РНК- характерна за tRNA, едноверижна, оформена като "лист от детелина".
Включва:
- относително къси двойни спирали - стъбла,
- едноверижни секции - бримки.
Има 4 стъбла (акцептор, антикодон, дихидроуридил, псевдоуридил) и 3 бримки.

"Stem-loop" - елемент от вторичната структура на РНК, схематично

"Псевдовъзел" - елемент от вторичната структура на РНК, схематично

Акцепторното стъбло съдържа 3'- и 5'-края на полинуклеотидната верига, 5'-краят завършва с остатък от гуанилова киселина, 3'-краят е CCA триплет и служи за образуване на естерна връзка с АА.
Стъблото на антикодон разпознава своя кодон върху иРНК в рибозомите по принципа на комплементарност.
Псевдоуридилното стъбло служи за прикрепване към рибозомата.
Дихидроуридиловата основа служи за свързване с аминоацил-тРНК синтетазата.
Социални бутони за Joomla
Ние също препоръчваме
 Импулсно захранване: ремонт и усъвършенстване
Импулсно захранване: ремонт и усъвършенстване
 Дистанционно управление на светлината
Дистанционно управление на светлината
 Уроци по плуване за деца в предучилищна възраст
Уроци по плуване за деца в предучилищна възраст
 Бележки за майстора - домашни битови аларми
Бележки за майстора - домашни битови аларми
 Часовниково витло на Atmega8
Часовниково витло на Atmega8
 Примери за приложение на устройства и реле, как да изберете и свържете правилно реле Микроконтролер и релейни прости превключващи вериги
Примери за приложение на устройства и реле, как да изберете и свържете правилно реле Микроконтролер и релейни прости превключващи вериги