Funktionella regioner av tRNA. Struktur och funktioner hos tRNA, egenskaper hos aminosyraaktivering
70-90N | sekundär sida - klöverblad | CCA 3"-konst för alla tRNA |
förekomsten av tymin, pseudouridin-psi, digirouridin DGU i D-loopen - skydd mot ribonukleaser? långlivade | En mängd olika primära strukturer av tRNA - 61 + 1 - med antalet kodoner + formylmetionin tRNA, kattens antikodon är detsamma som för metionin tRNA. Olika tertiära strukturer - 20 (beroende på antalet aminosyror) | igenkänning - bildandet av en kovalent bindning m-y tRNA och agera | aminoacyl-tRNA-syntetaser fäster handlingar till tRNA
tRNA:s funktion är att överföra aminosyror från cytoplasman till ribosomerna, där proteinsyntes sker.
tRNA som binder en aminosyra kallas isoacceptor.
Totalt existerar 64 olika tRNA samtidigt i en cell.
Varje tRNA parar endast med sitt eget kodon.
Varje tRNA känner igen sitt eget kodon utan inblandning av en aminosyra. Aminosyrorna bundna till tRNA:t modifierades kemiskt, varefter den resulterande polypeptiden, som innehöll den modifierade aminosyran, analyserades. Cysteinyl-tRNACys (R=CH2-SH) reducerades till alanyl-tRNACys (R=CH3).
De flesta tRNA, oavsett deras nukleotidsekvens, har en klöverbladsformad sekundär struktur på grund av närvaron av tre hårnålar i den.
Strukturella egenskaper hos tRNA
Det finns alltid fyra oparade nukleotider vid 3 "änden av molekylen, och tre av dem är nödvändigtvis CCAs. 5" och 3" ändarna av RNA-kedjan bildar en acceptorstam. Kedjorna hålls samman på grund av den komplementära parningen av sju nukleotider 5" - slutar med sju nukleotider belägna nära 3"-änden. 2. Alla molekyler har en T? C-hårnål, så betecknad eftersom den innehåller två ovanliga rester: ribotymidin (T) och pseudouridin (? Hårnålen består av en dubbel -strängad stam av fem parade baser, inklusive G-C-paret, och en slinga med sju nukleotider lång.
på samma punkt i slingan. 3. I en antikodon hårnål representeras alltid stammen av en familj av parade
grunder. Tripletten som är komplementär till det relaterade kodonet, antikodonet, finns i slingan.
le, bestående av sju nukleotider. En oföränderlig ura-
cyl och ett modifierat cytosin, och en modifierad purin gränsar till dess 3 "ände, som regel
adenin. 4. En annan hårnål består av en tre till fyra par långa nukleotider och en variabel slinga
storlek, ofta innehållande uracil i reducerad form - dihydrouracil (DU). Nukleotidsekvenserna för stjälkarna, antalet nukleotider mellan antikodonstammen och T?C-stammen (variabel loop), såväl som storleken på slingan och lokaliseringen av dihydrouracilrester i DU-slingan varierar kraftigast.
[Singer, 1998].
Tertiär struktur av tRNA
L-formad struktur.
Bindning av aminosyror till tRNA
För att en aminosyra ska bilda en polypeptidkedja måste den bindas till tRNA med enzymet aminoacyl-tRNA-syntetas. Detta enzym bildar en kovalent bindning mellan aminosyrakarboxylgruppen och riboshydroxylgruppen vid 3'-änden av tRNA med deltagande av ATP. Aminoacyl-tRNA-syntetas känner igen ett specifikt kodon inte på grund av närvaron av ett antikodon på tRNA:t, utan genom närvaron av ett specifikt igenkänningsställe på tRNA:t.
Totalt finns det 21 olika aminoacyl-tRNA-syntetaser i cellen.
Anslutningen sker i två steg:
1. Karboxylgruppen i en aminosyra är bunden till ATP a-fosfat. Det resulterande instabila aminoacyladenylatet stabiliseras genom bindning till enzymet.
2. Överföring av aminoacylgruppen i aminoacyladenylat till 2'- eller 3'-OH-gruppen i den terminala ribosen av tRNA
Vissa aminoacyl-tRNA-syntetaser består av en enda polypeptidkedja, medan andra består av två eller fyra identiska kedjor, var och en med en molekylvikt på 35 till 115 kDa. Vissa dimera och tetramera enzymer är sammansatta av två typer av subenheter. Det finns ingen tydlig korrelation mellan storleken på enzymmolekylen eller arten av dess underenhetsstruktur och specificitet.
Specificiteten för ett enzym bestäms av dess starka bindning till acceptoränden av tRNA, DU-regionen och den variabla slingan. Vissa enzymer verkar inte känna igen antikodontripletten och katalyserar aminoacetyleringsreaktionen även när antikodonet förändras. Vissa enzymer visar dock minskad aktivitet i förhållande till sådana modifierade tRNA och tillför fel aminosyra när antikodonet ersätts.
70-90n | sekundär sida - klöverblad | CCA 3"-konst för alla tRNA |
förekomsten av tymin, pseudouridin-psi, digirouridin DGU i D-loopen - skydd mot ribonukleaser? långlivade | En mängd olika primära strukturer av tRNA - 61 + 1 - med antalet kodoner + formylmetionin tRNA, kattens antikodon är detsamma som för metionin tRNA. Olika tertiära strukturer - 20 (beroende på antalet aminosyror)
Det finns två typer av tRNA-bindande metionin tRNAFMet och tRNAMMet i prokaryoter och tRNAIMet och tRNAMMet i eukaryoter. Metionin tillsätts till varje tRNA med användning av lämplig aminoacyl-tRNA-syntes. metionin fäst till tRNAFMet och tRNAIMet bildas av enzymet metionyl-tRNA-transformylas till Fmet-tRNAFMet. tRNA laddade med formylmetionin känner igen initieringskodonet AUG.
Litteratur:
Tyvärr finns det ingen bibliografi.
Lärobok. Trots det faktum att tRNA är mycket mindre, förtjänar en berättelse om dess struktur, egenskaper och funktion ett separat kapitel.
Så, tRNA är en "adapter", som känner igen sekvensen på tre bokstäver av den genetiska koden i ena änden, och matchar den med den enda motsvarande aminosyran fixerad i den andra änden av tRNA. I slutet av överförings-RNA:t som rör budbärar-RNA:t finns det 3 nukleotider som bildas antikodon. Endast om antikodonet är komplementärt till mRNA-regionen kan överförings-RNA:t ansluta sig till det. Men även i det här fallet kan tRNA inte gå med i mRNA på egen hand, det behöver hjälp av ribosomen, som är platsen för deras interaktion, såväl som en aktiv deltagare i översättningen. Till exempel är det ribosomen som skapar bindningar mellan aminosyror från tRNA och bildar en proteinkedja.
De strukturella egenskaperna hos tRNA bestäms av den genetiska koden, det vill säga reglerna för att konstruera ett protein enligt en gen som överförings-RNA:t läser. Den här koden fungerar i alla levande varelser på jorden: skapandet av ett virus skrivs med samma trebokstavskodon som används för att skriva "monteringsinstruktionerna" för en delfin. Det har experimentellt verifierats att generna från en levande varelse, placerad i en annans cell, kopieras perfekt och översätts till proteiner som inte går att skilja från generna som reproducerar i värdens celler. Den genetiska kodens enhetlighet är grunden för produktionen av modifierad E. coli av kolonier av insulin och många andra mänskliga enzymer som används som läkemedel för människor vars kroppar inte kan producera dem, eller producerar otillräckligt. Trots den uppenbara skillnaden mellan människor och E. coli, skapas mänskliga proteiner lätt från mänskliga ritningar med hjälp av en E. coli kopiator. Inte överraskande skiljer sig överförings-RNA från olika varelser väldigt lite.

Varje kodon från denna lista, förutom tre stoppa kodon, som signalerar fullbordandet av translationen, bör kännas igen av överförings-RNA:t. Igenkänning utförs genom att fästa ett antikodon till budbärar-RNA, som bara kan binda till ett kodon från listan, så tRNA kan bara känna igen ett kodon. Det betyder att det finns minst 61 typer av dessa molekyler i cellen. Faktum är att det finns ännu fler av dem, eftersom det i vissa situationer för att läsa budbärar-RNA inte räcker att bara ha rätt antikodon: andra villkor krävs, i enlighet med vilka ett speciellt, modifierat tRNA skapas.
Vid första anblicken borde en sådan mängd tRNA komplicera translationsprocessen avsevärt: trots allt kommer var och en av dessa molekyler att kontrollera matris-RNA-kodonet som ersatts av det av ribosomen för överensstämmelse med dess antikodon - det verkar som att så mycket meningslöst mekaniskt arbete , så mycket bortkastad tid och energi. Men som ett resultat av evolutionen har även cellulära mekanismer bildats som förhindrar detta problem. Till exempel motsvarar mängden tRNA för varje art i en cell hur ofta aminosyran som bärs av den arten finns i de proteiner som byggs. Det finns aminosyror som sällan används av cellen, och det finns de som ofta används, och om antalet tRNA som bär dem var detsamma skulle detta avsevärt komplicera sammansättningen av proteiner. Därför finns det få "sällsynta" aminosyror och deras motsvarande tRNA i cellen, medan frekvent förekommande aminosyror produceras i stora mängder.

Med en sådan variation av tRNA-molekyler är de alla väldigt lika, därför kommer vi, med tanke på deras struktur och funktioner, huvudsakligen studera egenskaperna som är gemensamma för alla arter. När du tittar på 3D-layouten av tRNA ser det ut som en tät hög med atomer. Det verkar otroligt att denna intrikat lindade molekyl är resultatet av veckningen av en lång kedja av nukleotider, men det är så den bildas.
Det är möjligt att spåra stadierna av dess bildning, från det allra första: sammanställningen av en nukleotidsekvens av RNA-polymeras i enlighet med genen som innehåller information om detta överförings-RNA. Ordningen i vilken dessa nukleotider följer varandra och deras nummer kallas primär struktur av tRNA. Det visar sig att det är den primära strukturen av tRNA som kodas i genen som läses av RNA-polymeras. I allmänhet är den primära strukturen en sekvens av relativt enkla molekyler av samma typ, av vilka en mer komplex, veckad polymermolekyl är sammansatt. Till exempel är den primära strukturen för en proteinmolekyl den enkla sekvensen av dess ingående aminosyror.

Någon kedja av nukleotider kan inte vara i ett ovikt tillstånd i en cell, utan bara sträckt ut i en linje. Det finns för många positivt och negativt laddade delar vid kanterna av nukleotiderna, som lätt bildar vätebindningar med varandra. Hur samma bindningar bildas mellan nukleotiderna i två DNA-molekyler, som förbinder dem till en dubbelspiral, beskrivs i, och för detaljer om vätebindningar, kan du klättra in. Vätebindningar är mindre starka än bindningarna mellan atomer i molekyler, men de räcker för att tvinna tRNA-tråden intrikat och hålla den i den positionen. Till en början bildas dessa bindningar endast mellan vissa nukleotider, vilket viker tRNA:t till en klöverbladsform. Resultatet av denna initiala vikning kallas sekundär struktur tRNA. Diagrammet till vänster visar att endast vissa nukleotider är sammanlänkade med vätebindningar, medan andra förblir oparade och bildar ringar och slingor. Skillnader mellan den sekundära strukturen för olika typer av tRNA beror på skillnader i deras primära struktur. Detta visar sig i olika längder av "klöverblad" eller "stjälk" på grund av olika längder på den initiala kedjan av nukleotider.
En annan skillnad i den primära strukturen för olika tRNA är att de endast i vissa positioner har samma nukleotider (i diagrammet ovan är de markerade med de första bokstäverna i deras namn), medan de flesta nukleotiderna i olika tRNA skiljer sig från varandra. Schemat ovan är gemensamt för alla tRNA, så olika nukleotider är markerade med siffror.
De huvudsakliga funktionella delarna av tRNA är:
=) antikodon det vill säga nukleotidsekvensen som är komplementär till ett enda kodon av budbärar-RNA beläget på antikodon hårnål
=) acceptor slut till vilken en aminosyra kan fästas. Den är belägen på motsatt sida av antikodonhårnålen.
I verkligheten ser inte ett enda tRNA ut som det gör i det sekundära strukturdiagrammet, eftersom bara några nukleotider gick samman för att bilda det, medan resten förblev oparade. På grund av bildandet av vätebindningar mellan nukleotider från olika delar av klöverbladet viks det ytterligare till ett mycket mer komplext tertiär struktur i form av ett L. Du kan förstå exakt hur de olika delarna av den sekundära strukturen kröks för att bilda den tertiära strukturen genom att matcha färgerna i deras diagram nedan. Antikodon-hårnålen, markerad i blått och grått, finns kvar längst ner (det är värt att komma ihåg att denna "botten" är villkorad: det är bekvämt att avbilda tRNA i denna rumsliga orientering i proteinöversättningsscheman) och acceptoränden (gul) är böjd åt sidan.

Så här ser tRNA ut när det är redo att fästa en aminosyra. tRNA kan inte kombineras med aminosyran på egen hand, detta kräver deltagande av ett speciellt enzym: aminoacyl-tRNA-syntetaser. Antalet syntetastyper i en cell sammanfaller med antalet tRNA-typer.
Likformigheten i formen av alla typer av tRNA är nödvändig för att ribosomen ska kunna känna igen någon av dem, underlätta deras dockning med mRNA och flytta inom sig själv från en plats till en annan. Om olika typer av tRNA skilde sig signifikant från varandra skulle detta göra ribosomens arbete extremt svårt, vilket kritiskt minskar hastigheten för proteinsyntesen. Naturligt urval syftar alltså till att göra tRNA liknande varandra. Men samtidigt finns det en annan faktor som kräver att det finns märkbara skillnader mellan olika typer av tRNA: trots allt är det nödvändigt att känna igen varje typ och fästa den enda motsvarande aminosyran till den. Uppenbarligen bör dessa skillnader vara märkbara, men inte för betydande, så att arbetet med att känna igen tRNA-arter förvandlas till en smyckesprocess. Och det är just detta som utförs av aminoacyl-tRNA-syntetaser: var och en av dem kan binda till endast en av de 20 aminosyrorna och fästa den exakt till de typer av tRNA som motsvarar denna aminosyra. Av tabellen med den genetiska koden kan man se att varje aminosyra kodas av flera nukleotidsekvenser, därför kommer till exempel alla fyra tRNA med antikodonen CGA, CGG, CGU och CGC att kännas igen av samma syntetas som fäster alanin till dem. Sådana tRNA som bearbetas av ett syntetas kallas relaterad.
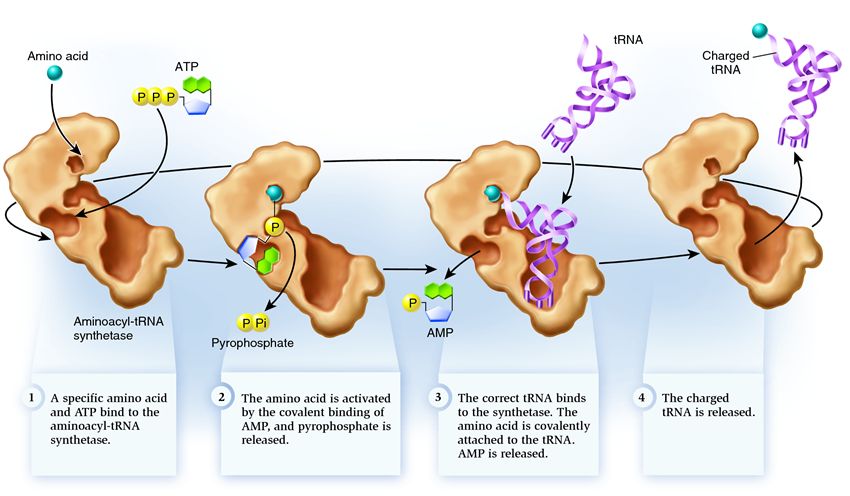
Syntetas tillhör en grupp enzymer vars funktion är att binda till separata existerande molekyler och kombinera dem till en:
1 . syntetas förbinder en aminosyra och en ATP-molekyl. Två fosfatgrupper bryter sig från ATP och frigör den energi som behövs för följande aktiviteter. Adenosinmonofosfatet (AMP) som finns kvar från den förstörda molekylen fäster vid aminosyran och förbereder den för anslutning till acceptorns hårnål.
2 . syntetas fäster till sig själv en av de relaterade tRNA:n som motsvarar denna aminosyra.

I detta skede kontrolleras överförings-RNA:s överensstämmelse med syntetaset. Det finns flera sätt att känna igen, och varje syntetas har en unik kombination av dem. Åtminstone en antikodonnukleotid är involverad i interaktionen mellan syntetas och tRNA. Acceptorhårnålen måste också kontrolleras: närvaron av specifika nukleotider på den som är gemensamma för relaterade tRNA som motsvarar den önskade aminosyran bestäms. Nukleotider från andra delar av tRNA kan också delta i matchning genom att binda till vissa syntetasställen. Fel tRNA kan matcha det önskade på något sätt, men på grund av ofullständig matchning kommer det att gå ihop med syntetaset långsamt och löst och lätt falla av. Och det korrekta tRNA kommer att fästa vid syntetaset snabbt och bestämt, som ett resultat av vilket syntetasets struktur förändras, vilket startar processen aminoacylering det vill säga bindningen av en aminosyra till tRNA.

3 . Aminoacylering består i att ersätta AMP-molekylen fäst vid aminosyran med en tRNA-molekyl. Efter denna ersättning lämnar AMP syntetaset och tRNA hålls uppe för en sista aminosyrakontroll. Om den bifogade aminosyran upptäcks som felaktig kommer den att lossna från tRNA:t, aminosyrans plats i syntetaset blir tom och ytterligare en molekyl kan ansluta sig dit. Den nya aminosyran kommer att gå igenom stadierna av anslutning till ATP och tRNA, och kommer också att testas. Om inga misstag gjordes frigörs det aminosyraladdade tRNA:t: det är redo att spela sin roll i proteinöversättning. Och syntetaset är redo att fästa nya aminosyror och tRNA, och cykeln kommer att börja på nytt.
Mycket beror på hur aminoacyl-tRNA-syntetaset fungerar korrekt: om ett fel inträffar i detta skede kommer fel aminosyra att fästas till tRNA:t. Och det kommer att byggas in i proteinet som växer på ribosomen, eftersom tRNA och ribosomen inte har funktionen att kontrollera överensstämmelsen mellan kodonet och aminosyran. Konsekvenserna av fel kan vara små eller katastrofala och genom naturligt urval har varelser med enzymer som inte har funktionen av sådana kontroller ersatts av mer anpassningsbara med olika alternativ för matchning mellan aminosyra och tRNA. Därför, i moderna celler, kombineras syntetas med fel aminosyra i genomsnitt i ett fall av 50 tusen, och med fel tRNA bara en gång av 100 tusen bilagor.

Vissa aminosyror skiljer sig från varandra med endast ett fåtal atomer. Om du tittar på deras scheman blir det uppenbart att sannolikheten för att förväxla arginin med alanin är mycket mindre än att förväxla isoleucin för leucin eller valin. Därför har varje syntetas som binder till en av aminosyrorna som liknar varandra ytterligare verifieringsmekanismer. Här är ett exempel på en sådan anpassning i isoleucin-tRNA-syntetas:

Varje syntas har syntetiskt centrum där en aminosyra är bunden till ett tRNA. Acceptorhårnålen för tRNA:t som fångas av syntetaset går dit, liksom aminosyran som är redo att reagera med den. Arbetet med vissa syntetaser slutar omedelbart efter anslutningen av aminosyran och tRNA. Men Ile-tRNA-syntetas har en ökad chans att göra misstag på grund av att det finns andra isoleucinliknande aminosyror. Därför har hon också kriminalvårdscentral: av namnet är det tydligt vilken roll det spelar i processen att koppla samman tRNA och aminosyror. Figuren till höger visar att positionen för änden av tRNA-acceptor-hårnålen i det syntetiska centrumet av Ile-tRNA-syntetas ger denna hårnål en onaturlig böjning. Syntetaset håller emellertid tRNA:t i denna position tills aminosyran är fäst vid den. Efter att denna koppling har inträffat, är behovet av att hitta acceptorhårnålen i det syntetiska centret uttömt, och tRNA:t rätar ut och får sin ände med aminosyran fäst vid den in i korrigeringscentret.

Det syntetiska centret spelar förstås också en roll för att sålla bort aminosyror som inte är lämpliga för syntetas. För att komma in i det måste molekylen uppfylla ett antal villkor, bland annat ha rätt storlek. Trots att leucin och isoleucin innehåller samma antal atomer, på grund av skillnader i den rumsliga strukturen, är leucin större. Därför kan den inte tränga in i det syntetiska centret, vars storlek motsvarar det mer kompakta isoleucinet, och studsar helt enkelt av Ile-tRNA-syntetaset.
Men valin, som är den minsta av dessa tre molekyler med liknande atomstruktur, tar lätt isoleucins plats i det syntetiska centret, och syntetas fäster det till tRNA. Det är i det här fallet som syntetasets korrektionscentrum kommer in i bilden. Om hårnålen för utriktningsacceptorn är korrekt laddad och bär isoleucin, kan den inte klämmas in i korrigeringscentret: den är helt enkelt för liten för denna molekyl. Det uträtade tRNA:t hålls alltså inte längre av någonting, och det lösgörs från syntetaset. Men om valin är fäst vid tRNA, glider det in i korrigeringscentret och håller därigenom tRNA:t kopplat till det i syntetaset. En sådan överdrivet lång vistelse av tRNA inuti är en felsignal för syntetaset, vilket ändrar dess rumsliga konfiguration. Som ett resultat:
=) valin lösgörs från tRNA och avlägsnas från syntetaset
=) Acceptorhårnålen återvänder till det syntetiska stället, i väntan på bindning till aminosyran
=) syntetas binder till en ny aminosyra, "laddar" tRNA med den och kontrollerar igen om isoleucin användes för detta.
En liknande dubbeligenkänningsmekanism används av andra syntetaser.
Fysikaliska och kemiska egenskaper hos DNA
Olika faktorer som bryter vätebindningar (temperaturhöjning över 80 C, förändringar i pH och jonstyrka, inverkan av urea etc.) orsakar DNA-denaturering, d.v.s. förändring i det rumsliga arrangemanget av DNA-kedjor utan att bryta kovalenta bindningar. Den dubbla helixen av DNA under denaturering är helt eller delvis uppdelad i dess komponentkedjor. DNA-denaturering åtföljs av en ökning av optisk absorption i UV-regionen av purin- och pyrimidinbaser. Detta fenomen kallas hyperkrom effekt . Denaturering minskar också den höga viskositeten som är inneboende i naturliga DNA-lösningar. När den ursprungliga dubbelsträngade DNA-strukturen återställs, som ett resultat av renaturering, minskar absorptionen vid 260 nm av kvävehaltiga baser på grund av deras "avskärmning". Detta fenomen kallas hypokrom effekt .
"Avlindningen" av varje DNA till dess komponentkedjor utförs inom ett visst temperaturintervall. Mittpunkten av detta intervall kallas smältpunkten. Smälttemperaturen för DNA beror under standardförhållanden (ett visst pH och jonstyrka) på förhållandet mellan kvävehaltiga baser. G-C-par som innehåller tre vätebindningar är starkare, därför, ju högre innehållet av G-C-par i DNA, desto högre smältpunkt.
Funktioner av DNA. I sekvensen av nukleotider i DNA-molekyler kodas genetisk information. DNA:s huvudsakliga funktioner är för det första att säkerställa reproduktionen av sig själv i en serie av cellgenerationer och generationer av organismer, och för det andra att säkerställa syntesen av proteiner. Dessa funktioner beror på att DNA-molekyler fungerar som en matris i det första fallet för replikering, d.v.s. kopiera information i dotter-DNA-molekyler, i den andra - för transkription, dvs. för att omkoda information till RNA-strukturen.
Ris. 5 Smältkurva (DNA-denaturering)
Komplementära DNA-strängar som separeras under denaturering kan under vissa förhållanden återförenas till en dubbelhelix. Denna process kallas RENATURING. Om denaturering inte har skett fullständigt och åtminstone ett fåtal baser inte har förlorat interaktion med vätebindningar, fortskrider renatureringen mycket snabbt.
Cellernas cytoplasma innehåller tre huvudsakliga funktionella typer av RNA. Dessa är budbärar-RNA – mRNA som fungerar som mallar för proteinsyntes, ribosomala RNA – rRNA som fungerar som strukturella komponenter i ribosomer och transfer-RNA – tRNA som är involverade i translationen (translationen) av mRNA-information till aminosyrasekvensen i proteinet.
Tabell 2 visar skillnaderna mellan DNA och RNA vad gäller struktur, lokalisering i cellen och funktioner.
Tabell 2 Skillnader mellan DNA och RNA
Överför RNA, tRNA-ribonukleinsyra, vars funktion är att transportera AA till platsen för proteinsyntes. Den har en typisk längd på 73 till 93 nukleotider och en storlek på cirka 5 nm. tRNA är också direkt involverade i tillväxten av polypeptidkedjan, förenar sig - är i ett komplex med en aminosyra - till mRNA-kodonet och ger den konformation av komplexet som är nödvändigt för bildandet av en ny peptidbindning. Varje aminosyra har sitt eget tRNA. tRNA är ett enkelsträngat RNA, men i sin funktionella form har det en klöverbladskonformation. AA fäster kovalent till 3"-änden av molekylen med hjälp av enzymet aminoacyl-tRNA-syntetas, specifikt för varje typ av tRNA. På plats C finns ett antikodon som motsvarar AA-te. tRNA syntetiseras av vanligt RNA-polymeras i fallet av prokaryoter och av RNA-polymeras III i fallet med eukaryoter. Transkript av tRNA-gener genomgår flerstegsbearbetning, vilket leder till bildandet av en rumslig struktur som är typisk för tRNA.
tRNA-bearbetning involverar 5 nyckelsteg:
avlägsnande av 5"-ledarnukleotidsekvensen;
avlägsnande av den 3'-terminala sekvensen;
lägga till en CCA-sekvens vid 3"-änden;
excision av introner (i eukaryoter och archaea);
modifieringar av enskilda nukleotider.
Transport av tRNA utförs längs en Ran-beroende väg med deltagande av transportfaktorn exportin t, som känner igen den karakteristiska sekundära och tertiära str-ru av moget tRNA: korta dubbelsträngade sektioner och korrekt bearbetade 5 "- och 3" slutar. Denna mekanism säkerställer att endast mogna tRNA exporteras från kärnan.
62. Translation - mRNA-kodonigenkänning
Translation är en proteinsyntes som utförs av ribosomer från aminosyror på en mRNA (eller och RNA) mall. Översättningsprocessens beståndsdelar: aminosyror, tRNA, ribosomer, mRNA, enzymer för aminoacylering av tRNA, proteintranslationsfaktorer (proteinfaktorer för initiering, förlängning, avslutning - specifika extraribosomala proteiner som behövs för translationsprocesser), ATP- och GTP-energikällor magnesiumjoner (stabiliserar ribosomstrukturen). 20 aminosyror är involverade i proteinsyntesen. För att en aminosyra ska "känna igen" sin plats i den framtida polypeptidkedjan måste den binda till ett transfer-RNA (tRNA) som utför en adapterfunktion. tRNA:t som binder till aminosyran känner sedan igen motsvarande kodon på mRNA:t. mRNA-kodonigenkänning:
Kodon-antikodon-interaktionen är baserad på principerna om komplementaritet och antiparallelism:
3'----C - G-A*------5' tRNA antikodon
5'-----G-C-Y*------3' mRNA kodon
Wobble-hypotesen föreslogs av F. Crick:
3'-basen av mRNA-kodonet har en icke-strikt parning med 5'-basen av tRNA-antikodonet: till exempel kan Y (mRNA) interagera med A och G (tRNA)
Vissa tRNA kan paras ihop med mer än ett kodon.
63. Egenskaper för de ingående delarna av översättningsprocessen. Translation (translatio-translation) är processen för proteinsyntes från aminosyror på matrisen av informativt (matris) RNA (mRNA, mRNA) som utförs av ribosomen.
Proteinsyntes är grunden för cellliv. För att utföra denna process i cellerna i alla organismer finns det speciella organeller - ribosomer- ribonukleoproteinkomplex byggda av 2 subenheter: stora och små. Ribosomernas funktion är att känna igen tre bokstäver (tre-nukleotider) kodon mRNA, jämför dem med motsvarande tRNA-antikodon som bär aminosyror och tillsatsen av dessa aminosyror till den växande proteinkedjan. När den rör sig längs mRNA-molekylen, syntetiserar ribosomen ett protein i enlighet med informationen i mRNA-molekylen.
För igenkänning av AK-t i cellen finns det speciella "adaptrar", överföra RNA-molekyler(tRNA). Dessa klöverbladsformade molekyler har ett ställe (antikodon) som är komplementärt till ett mRNA-kodon, såväl som ett annat ställe till vilket aminosyran som motsvarar det kodonet är fäst. Bindningen av aminosyror till tRNA utförs i en energiberoende reaktion av enzymer aminoacyl-tRNA-syntetaser, och den resulterande molekylen kallas aminoacyl-tRNA. Sålunda bestäms translationens specificitet av interaktionen mellan mRNA-kodonet och tRNA-antikodonet, såväl som specificiteten hos aminoacyl-tRNA-syntetaser som binder aminosyror strikt till deras motsvarande tRNA (till exempel kommer GGU-kodonet att motsvara en tRNA som innehåller CCA-antikodonet och endast AK-glycin).
prokaryot ribosom
5S och 23S rRNA 16S rRNA
34 ekorrar 21 ekorrar
Prokaryota ribosomer har en sedimentationskonstant på 70S, vilket är anledningen till att de kallas 70S-partiklar. De är byggda av två olika underenheter: 30S och 50S underenheter. Varje subenhet är ett komplex av rRNA och ribosomala proteiner.
30S-partikeln innehåller en 16S rRNA-molekyl och i de flesta fall en proteinmolekyl från mer än 20 arter (21) . 50S-subenheten består av två rRNA-molekyler (23S och 5S). Den består av mer än 30 olika proteiner (34), som också representeras, som regel, av en kopia. De flesta av de ribosomala proteinerna har en strukturell funktion.
eukaryot ribosom
5S; 5,8S och 28S rRNA 18S rRNA
minst 50 proteiner minst 33 proteiner
Ribosomen består av stora och små underenheter. Grunden för strukturen för varje subenhet är ett komplext veckat rRNA. Ribosomproteiner fästes till rRNA-ställningen.
Sedimentationskoefficienten för en komplett eukaryot ribosom är cirka 80 Svedberg-enheter (80S), och sedimentationskoefficienten för dess subpartiklar är 40S och 60S.
Den mindre 40S-subenheten består av en 18S rRNA-molekyl och 30-40 proteinmolekyler. Den stora 60S-subenheten innehåller tre typer av rRNA med sedimentationskoefficienter på 5S, 5.8S och 28S och 40-50 proteiner (till exempel inkluderar råtthepatocytribosomer 49 proteiner).
Funktionella regioner av ribosomer
P - peptidylställe för peptidyl-tRNA
A - aminoacylställe för aminoacyl-tRNA
E - plats för frisättning av tRNA från ribosomen
Ribosomen innehåller 2 funktionella ställen för interaktion med tRNA: aminoacyl (acceptor) och peptidyl (donator). Aminoacyl-tRNA går in i acceptorstället i ribosomen och interagerar för att bilda vätebindningar mellan kodon och antikodontripletter. Efter bildandet av vätebindningar flyttar systemet fram 1 kodon och hamnar på donatorstället. Samtidigt uppträder ett nytt kodon i det lediga acceptorstället, och motsvarande aminoacyl-t-RNA fästs till det.
Ribosomer: struktur, funktion
Ribosomer är de cytoplasmatiska centra för proteinbiosyntes. De består av stora och små underenheter, som skiljer sig i sedimentationskoefficienter (sedimentationshastighet under centrifugering), uttryckt i enheter av Svedberg - S.
Ribosomer finns i både eukaryota och prokaryota celler, eftersom de fyller en viktig funktion i proteinbiosyntes. Varje cell innehåller tiotals, hundratusentals (upp till flera miljoner) av dessa små rundade organeller. Det är en rundad ribonukleoproteinpartikel. Dess diameter är 20-30 nm. Ribosomen består av stora och små subenheter, som skiljer sig i sedimentationskoefficienter (sedimentationshastighet under centrifugering), uttryckt i Svedberg-enheter - S. Dessa subenheter kombineras i närvaro av en sträng av m-RNA (matris, eller informations-RNA). Ett komplex av en grupp ribosomer förenade av en enda mRNA-molekyl som en sträng av pärlor kallas polysom. Dessa strukturer är antingen fritt belägna i cytoplasman eller fästa vid membranen i den granulära ER (i båda fallen fortsätter proteinsyntesen aktivt på dem).
Polysomer av granulär ER bildar proteiner som utsöndras från cellen och används för hela organismens behov (till exempel matsmältningsenzymer, proteiner från human bröstmjölk). Dessutom finns ribosomer på den inre ytan av mitokondriella membran, där de också deltar aktivt i syntesen av proteinmolekyler.
Transfer-RNA (tRNA) spelar en viktig roll i processen att använda ärftlig information av cellen. Genom att leverera de nödvändiga aminosyrorna till sammansättningsstället för peptidkedjor, fungerar tRNA som en translationell mediator.
tRNA-molekyler är polynukleotidkedjor syntetiserade på specifika DNA-sekvenser. De består av ett relativt litet antal nukleotider -75-95. Som ett resultat av den komplementära kopplingen av baser som är belägna i olika delar av tRNA-polynukleotidkedjan, får den en struktur som liknar ett klöverblad till formen (Fig. 3.26).
Ris. 3,26. Strukturen hos en typisk tRNA-molekyl.
Den har fyra huvuddelar som utför olika funktioner. acceptor"Stjälken" bildas av två komplementärt sammankopplade terminala delar av tRNA:t. Den består av sju baspar. 3'-änden av denna stam är något längre och bildar en enkelsträngad region som slutar med en CCA-sekvens med en fri OH-grupp. En transportabel aminosyra är fäst till detta ändamål. De återstående tre grenarna är komplementära parade nukleotidsekvenser som slutar i oparade sektioner som bildar loopar. Mitten av dessa grenar - antikodon - består av fem par nukleotider och innehåller ett antikodon i mitten av dess loop. Antikodonet är tre nukleotider som är komplementära till mRNA-kodonet, som kodar för aminosyran som transporteras av detta tRNA till platsen för peptidsyntes.
Mellan acceptor- och antikodongrenarna finns två sidogrenar. I sina loopar innehåller de modifierade baser - dihydrouridin (D-loop) och TψC-tripletten, där \y är pseudouriain (T^C-loop).
Mellan aitikodon- och T^C-grenarna finns en extra loop, som inkluderar från 3-5 till 13-21 nukleotider.
I allmänhet kännetecknas olika typer av tRNA av en viss beständighet i nukleotidsekvensen, som oftast består av 76 nukleotider. Variationen i deras antal beror huvudsakligen på förändringen av antalet nukleotider i den extra slingan. Komplementära regioner som stöder tRNA-strukturen är vanligtvis konserverade. Den primära strukturen av tRNA, bestäms av sekvensen av nukleotider, bildar den sekundära strukturen av tRNA, som har formen av ett klöverblad. Den sekundära strukturen orsakar i sin tur en tredimensionell tertiär struktur, som kännetecknas av bildandet av två vinkelräta dubbla helixar (Fig. 3.27). En av dem bildas av acceptor- och TψC-grenarna, den andra av antikodon- och D-grenarna.
I slutet av en av de dubbla helixarna finns den transporterade aminosyran, i slutet av den andra finns antikodonet. Dessa områden ligger längst bort från varandra. Stabiliteten hos den tertiära strukturen av tRNA bibehålls på grund av uppkomsten av ytterligare vätebindningar mellan baserna i polynukleotidkedjan, belägna i olika delar av den, men rumsligt nära i den tertiära strukturen.
Olika typer av tRNA har en liknande tertiär struktur, men med vissa variationer.
Ris. 3,27. Rumslig organisation av tRNA:
I - den sekundära strukturen av tRNA i form av ett "klöverblad", bestämt av dess primära struktur (sekvensen av nukleotider i kedjan);
II - tvådimensionell projektion av den tertiära strukturen av tRNA;
III - layout av tRNA-molekylen i rymden
BILAGA (ifall någon inte förstår detta)
Blixtande tänder - nukleotider (adenin-tymin / uracil /, guanin-cytazin). Alla blixtar är DNA.

För att överföra information från DNA måste du bryta 2 strängar. Bindningen mellan A-T och G-C är väte, därför bryts den lätt av Helicase-enzymet:

För att förhindra att knutar bildas (Som ett exempel vred jag en handduk):



Topoisomeras skär en DNA-sträng vid replikationsursprunget så att kedjan inte vrids.

När en tråd är ledig kan den andra lätt rotera runt sin axel och därigenom avlasta spänningen under "avlindning". Noder visas inte, energi sparas.
Sedan behövs en RNA-primer för att börja samla RNA. Ett protein som sätter ihop mRNA kan inte bara sätta ihop den första nukleotiden, det behöver en bit RNA för att starta (det är skrivet i detalj där, jag skriver det senare). Denna bit kallas RNA-primern. Och detta protein fäster redan den första nukleotiden till det.


När strukturen av nukleinsyror beskrivs, beaktas olika nivåer av organisering av makromolekyler: primär och sekundär struktur.
Nukleinsyrors primära struktur är nukleotidsammansättningen och en viss sekvens av nukleotidenheter i polymerkedjan.
Sekundär struktur av RNA. Ribonukleinsyramolekylen är byggd av en enda polynukleotidkedja.
Sekundär struktur av RNA
Separata sektioner av RNA-kedjan bildar spiraliserade slingor - "hårnålar", på grund av vätebindningar mellan de komplementära kvävebaserna A-U och G-C. Sektioner av RNA-kedjan i sådana spiralstrukturer är antiparallella, men inte alltid helt komplementära, de innehåller oparade nukleotidrester eller till och med enkelsträngade slingor som inte passar in i dubbelhelixen. Närvaron av spiraliserade regioner är karakteristisk för alla typer av RNA.
Huvudrollen för RNA är direkt deltagande i proteinbiosyntes.
Tre typer av cellulärt RNA är kända, som skiljer sig åt i deras placering i cellen, sammansättning, storlek och egenskaper som bestämmer deras specifika roll i bildandet av proteinmakromolekyler:
- informations-(matris)RNA:n överför information kodad i DNA om proteinets struktur från cellkärnan till ribosomerna, där proteinsyntesen utförs; den primära strukturen för alla mRNA, oavsett unikheten hos deras kodande sekvens, har samma struktur för 5'- och 3'-ändarna.
Så vid 5'-änden finns en modifierad nukleotid 7-metylguanosin-5'-trifosfat (cap). Flera tiotals nukleotider separerar locket från initieringskodonet, vanligtvis -AUG- tripletten. Den kodande regionen följs av ett av termineringskodonen -UGA-, -UUA-, -UAG-. I 3'-änden av de flesta mRNA finns en nukleotidsekvens på 100-200 adenosinmonofosfatrester.
- transfer RNA samlar aminosyror i cellens cytoplasma och överför dem till ribosomen; RNA-molekyler av denna typ "lär sig" från motsvarande delar av budbärar-RNA-kedjan vilka aminosyror som bör delta i proteinsyntesen.
Den rumsliga strukturen för varje tRNA, oavsett skillnader i nukleotidsekvensen, beskrivs av den universella klöverbladsmodellen. Varje tRNA-molekyl har kedjesektioner som inte är involverade i bildandet av vätebindningar mellan nukleotidrester.
Dessa inkluderar i synnerhet det ställe som är ansvarigt för bindning till aminosyran vid 3'-änden av molekylen och antikodonet, en specifik triplett av nukleotider som interagerar komplementärt med mRNA-kodonet.
- Ribosomalt RNA tillhandahåller proteinsyntes av en viss struktur, läser information från informations-(matris)-RNA. rRNA bildar komplex med proteiner som kallas ribosomer.
Varje ribosom består av två underenheter - liten (40S) och stor (60S). Ribosomsubenheter skiljer sig inte bara i uppsättningen av rRNA, utan också i antalet och strukturen av proteiner.
Publiceringsdatum: 2015-02-03; Läst: 2729 | Sidans upphovsrättsintrång

RNA är en polymer vars monomerer är ribonukleotider.
Till skillnad från DNA, bildas RNA inte av två, utan av en polynukleotidkedja (undantag - vissa RNA-innehållande virus har dubbelsträngat RNA). RNA-nukleotider kan bilda vätebindningar med varandra. RNA-kedjor är mycket kortare än DNA-kedjor.
RNA-monomeren - nukleotid (ribonukleotid) - består av rester av tre ämnen: 1) en kvävebas, 2) en monosackarid med fem kolatomer (pentos) och 3) fosforsyra. De kvävehaltiga baserna av RNA tillhör också klasserna pyrimidiner och puriner.
Pyrimidinbaser av RNA - uracil, cytosin, purinbaser - adenin och guanin.
31. Typer av RNA och egenskaper hos dess struktur
RNA-nukleotidmonosackariden representeras av ribos.
Det finns tre typer av RNA: 1) information (matris) RNA - mRNA (mRNA), 2) överförings-RNA - tRNA, 3) ribosomalt RNA - rRNA.
Alla typer av RNA är ogrenade polynukleotider, har en specifik rumslig konformation och deltar i processerna för proteinsyntes.
Information om strukturen hos alla typer av RNA lagras i DNA. Processen för RNA-syntes på en DNA-mall kallas vanligtvis transkription.

Transfer-RNA innehåller vanligtvis 76 (från 75 till 95) nukleotider; molekylvikt - 25 000–30 000.
tRNA står för cirka 10 % av det totala RNA-innehållet i cellen. Funktioner av tRNA: 1) transport av aminosyror till platsen för proteinsyntes, till ribosomer, 2) translationsmediator. Cirka 40 typer av tRNA finns i cellen, var och en av dem har en nukleotidsekvens som är karakteristisk endast för den. Samtidigt har alla tRNA flera intramolekylära komplementära regioner, på grund av vilka tRNA får en konformation som liknar ett klöverblad i form.
Varje tRNA har en loop för kontakt med ribosomen (1), en antikodonloop (2), en loop för kontakt med enzymet (3), en acceptorstam (4) och ett antikodon (5). Aminosyran är fäst vid 3'-änden av acceptorstammen. Antikodon - tre nukleotider som "känner igen" ett mRNA-kodon.
Det bör betonas att ett visst tRNA kan transportera en strikt definierad aminosyra som motsvarar dess antikodon. Specificiteten för kopplingen av aminosyror och tRNA uppnås på grund av egenskaperna hos enzymet aminoacyl-tRNA-syntetas.
Ribosomala RNA innehåller 3000–5000 nukleotider; molekylvikt - 1 000 000–1 500 000.
rRNA står för 80–85 % av det totala RNA-innehållet i cellen. I komplex med ribosomala proteiner bildar rRNA ribosomer - organeller som utför proteinsyntes. I eukaryota celler sker rRNA-syntes i nukleolen. Funktioner av rRNA: 1) en nödvändig strukturell komponent av ribosomer och därmed säkerställande av ribosomernas funktion; 2) säkerställande av interaktionen mellan ribosomen och tRNA; 3) den initiala bindningen av ribosomen och mRNA-initiatorkodonet och bestämningen av läsramen, 4) bildandet av ribosomens aktiva centrum.
Messenger-RNA är olika i nukleotidinnehåll och molekylvikt (från 50 000 till 4 000 000).
Andelen mRNA står för upp till 5 % av det totala RNA-innehållet i cellen. Funktioner hos mRNA: 1) överföring av genetisk information från DNA till ribosomer, 2) en matris för syntes av en proteinmolekyl, 3) bestämning av aminosyrasekvensen för en proteinmolekyls primära struktur.
Läs också
RNA är en polymer vars monomerer är nukleotider.
De tre kvävehaltiga baserna är desamma som i DNA (adenin, guanin, cytosin); den fjärde - uracil - finns i RNA-molekylen istället för tymin. RNA-nukleotider innehåller ribos istället för deoxiribos. I RNA-kedjan...
tre huvudtyper av RNA: informativt(mRNA), eller matris(mRNA), ribosomalt(rRNA), och transport(tRNA). De skiljer sig åt i molekylstorlek och funktion. Alla typer av RNA syntetiseras på DNA med deltagande av enzymer - RNA-polymeraser. Messenger-RNA utgör 2-3% av allt cellulärt RNA, ribosomalt - 80-85, transport - cirka 15%.
mRNA.
den läser ärftlig information från ett DNA-segment och, i form av en kopierad sekvens av kvävebaser, överför den till ribosomer, där ett visst protein syntetiseras. Var och en av mRNA-molekylerna i nukleotidernas storlek och storlek motsvarar genen i DNA från vilken den transkriberades. I genomsnitt innehåller mRNA 1500 nukleotider (75-3000). Varje triplett (tre nukleotider) på ett mRNA kallas ett kodon. Det beror på kodonet vilken aminosyra som kommer att dyka upp på en given plats under proteinsyntesen.
(tRNA) har en relativt låg molekylvikt på cirka 24-29 tusen.
D och innehåller från 75 till 90 nukleotider i molekylen. Upp till 10% av alla tRNA-nukleotider är mindre baser, vilket uppenbarligen skyddar det från inverkan av hydrolytiska enzymer.TRNA:s roll är att de överför aminosyror till ribosomer och deltar i processen för proteinsyntes. Varje aminosyra fäster till ett specifikt tRNA. Ett antal aminosyror har mer än ett tRNA. Hittills har mer än 60 tRNA upptäckts som skiljer sig åt i sin primära struktur (bassekvens).
Den sekundära strukturen av alla tRNA presenteras i form av ett klöverblad med en dubbelsträngad stam och tre enkelsträngade). I slutet av en av kedjorna finns ett acceptorställe - CCA-tripletten, till vars adenin en specifik aminosyra är fäst.
(rRNA). De innehåller 120-3100 nukleotider. Ribosomalt RNA ackumuleras i kärnan, i nukleolerna.
Ribosomala proteiner transporteras till nukleolerna från cytoplasman och där sker spontan bildning av ribosomala subpartiklar genom att kombinera proteiner med motsvarande rRNA. Ribosomens underpartiklar transporteras tillsammans eller separat genom porerna i kärnmembranet in i cytoplasman. Ribosomerär organeller 20-30 nm stora.
De är byggda av två underpartiklar av olika storlekar och former. Vid vissa stadier av proteinsyntesen i cellen delas ribosomerna in i subpartiklar.
Ribosomalt RNA fungerar som ett ramverk för ribosomer och underlättar den initiala bindningen av mRNA till ribosomen under proteinbiosyntes.
Fråga 6 Bindningarna som bildar de primära och sekundära strukturerna av DNA och RNA. Typer av RNA
Den genetiska koden är ett sätt att koda för aminosyrasekvensen hos proteiner med hjälp av en sekvens av nukleotider, som är karakteristisk för alla levande organismer.
Egenskaper: 1) genetisk kod trilling(varje aminosyra kodas av tre nukleotider); 2) ej överlappande(intilliggande tripletter har inte gemensamma nukleotider); 3) degenererad(med undantag för metionin och tryptofan har alla aminosyror mer än ett kodon); 4) universell(för det mesta samma för alla levande organismer); 5) i kodon för en aminosyra är de två första nukleotiderna vanligtvis desamma och den tredje varierar; 6) har en linjär läsordning och kännetecknas av kolinearitet, t.
e. sammanfallande ordningen av kodonen i mRNA:t med ordningen för aminosyrorna i den syntetiserade polypeptidkedjan.
Publiceringsdatum: 2014-12-08; Läs: 11268 | Sidans upphovsrättsintrång
studopedia.org - Studopedia.Org - 2014-2018. (0,001 s) ...
Cellernas cytoplasma innehåller tre huvudsakliga funktionella typer av RNA:
- budbärar-RNA (mRNA) som fungerar som mallar för proteinsyntes;
- ribosomalt RNA (rRNA) som fungerar som strukturella komponenter i ribosomer;
- överföra RNA (tRNA) involverade i translationen (translationen) av mRNA-information till aminosyrasekvensen för en proteinmolekyl.
I cellkärnan finns nukleärt RNA, som utgör från 4 till 10 % av det totala cellulära RNA:t.
Huvuddelen av nukleärt RNA representeras av högmolekylära prekursorer av ribosomalt och transport-RNA. Prekursorer för högmolekylära rRNA (28 S, 18 S och 5 S RNA) är huvudsakligen lokaliserade i nukleolus.
RNA är det huvudsakliga genetiska materialet i vissa djur- och växtvirus (genomiskt RNA). De flesta RNA-virus kännetecknas av omvänd transkription av deras RNA-genom, styrd av omvänt transkriptas.
Alla ribonukleinsyror är polymerer av ribonukleotider kopplade, som i en DNA-molekyl, med 3',5'-fosforodiesterbindningar.
Till skillnad från DNA, som har en dubbelsträngad struktur, är RNA en enkelsträngad linjär polymer molekyl.
mRNA-struktur. mRNA är den mest heterogena klassen av RNA vad gäller storlek och stabilitet.
tRNA struktur.
Transfer-RNA fungerar som mediatorer (adaptrar) under mRNA-translation. De står för cirka 15 % av det totala cellulära RNA:t. Var och en av de 20 proteinogena aminosyrorna har sitt eget tRNA. För vissa aminosyror som kodas av två eller flera kodon, finns det flera tRNA.
tRNA är relativt små enkelsträngade molekyler som består av 70-93 nukleotider. Deras molekylvikt är (2,4-3,1) 0,104 kDa.
Den sekundära strukturen av tRNA bildas på grund av bildandet av det maximala antalet vätebindningar mellan intramolekylära komplementära par av kvävehaltiga baser.
Som ett resultat av bildandet av dessa bindningar vrids tRNA-polynukleotidkedjan med bildandet av spiraliserade grenar som slutar i loopar av oparade nukleotider. Den rumsliga bilden av de sekundära strukturerna av alla tRNA har formen av ett klöverblad.
Fyra obligatoriska grenar urskiljs i "klöverbladet", längre tRNA innehåller dessutom en kort femte (ytterligare) gren.
Adaptorfunktionen hos tRNA tillhandahålls av en acceptorgren, till vars 3'-ände en aminosyrarest är fäst med en eterbindning, och en antikodongren mittemot acceptorgrenen, i toppen av vilken det finns en slinga som innehåller ett antikodon.
Ett antikodon är en specifik triplett av nukleotider som är komplementär i antiparallell riktning till mRNA-kodonet som kodar för motsvarande aminosyra.
T-grenen som bär pseudouridinslingan (TyC-loop) säkerställer interaktionen av tRNA med ribosomer.
D-grenen, som bär dehydrouridinslingan, säkerställer interaktionen av tRNA med motsvarande aminoacyl-tRNA-syntetas.

Sekundär struktur av tRNA
Funktionerna för den femte ytterligare grenen är fortfarande dåligt förstådda, troligen utjämnar den längden på olika tRNA-molekyler.
Den tertiära strukturen av tRNA är mycket kompakt och bildas genom att sammanföra individuella grenar av klöverbladet på grund av ytterligare vätebindningar för att bilda en L-formad "armbågsböj"-struktur.
Transport-RNA, struktur och funktionell mekanism.
I det här fallet är acceptorarmen som binder aminosyran placerad i ena änden av molekylen och antikodonet i den andra.

Tertiär struktur av tRNA (enligt A.S. Spirin)
Strukturen av rRNA och ribosomer. Ribosomala RNA bildar ryggraden till vilken specifika proteiner binder för att bilda ribosomer. Ribosomer är nukleoproteinorganeller som tillhandahåller proteinsyntes från mRNA.
Antalet ribosomer i en cell är mycket stort: från 104 i prokaryoter till 106 i eukaryoter. Ribosomer är lokaliserade främst i cytoplasman, i eukaryoter, dessutom i nukleolen, i mitokondriella matrisen och i kloroplasternas stroma. Ribosomer består av två subpartiklar: stora och små. Efter storlek och molekylvikt är alla studerade ribosomer indelade i 3 grupper - 70S ribosomer av prokaryoter (S-sedimentationskoefficient), bestående av små 30S och stora 50S subpartiklar; 80S eukaryota ribosomer, bestående av 40S små och 60S stora subenheter.
Den lilla subenheten av 80S-ribosomer bildas av en rRNA-molekyl (18S) och 33 molekyler av olika proteiner.
Den stora subenheten bildas av tre rRNA-molekyler (5S, 5.8S och 28S) och cirka 50 proteiner.
Den sekundära strukturen av rRNA bildas på grund av korta dubbelsträngade sektioner av molekylen - hårnålar (ca 2/3 av rRNA), 1/3 - representeras av enkelsträngade sektioner rika på purinnukleotider.
Sociala knappar för Joomla
Proteiner utgör grunden för livet. Deras funktioner i cellen är mycket olika. Proteiner "kan" dock inte reproducera sig. Och all information om proteiners struktur finns i gener (DNA).
I högre organismer syntetiseras proteiner i cellens cytoplasma, och DNA är gömt bakom skalet av kärnan. Därför kan DNA inte direkt tjäna som mall för proteinsyntes. Denna roll utförs av en annan nukleinsyra - RNA.
RNA-molekylen är en ogrenad polynukleotid med en tertiär struktur.
Den bildas av en polynukleotidkedja, och även om de komplementära nukleotiderna som ingår i den också kan bilda vätebindningar mellan sig, uppstår dessa bindningar mellan nukleotiderna i en kedja. RNA-kedjor är mycket kortare än DNA-kedjor. Om innehållet av DNA i en cell är relativt konstant, så fluktuerar innehållet av RNA kraftigt. Den största mängden RNA i celler observeras under proteinsyntes.
RNA spelar en stor roll vid överföring och implementering av ärftlig information.
I enlighet med funktionen och strukturella egenskaper särskiljs flera klasser av cellulärt RNA.
Det finns tre huvudklasser av cellulärt RNA.
- Informations- (mRNA) eller matris (mRNA). Dess molekyler är de mest olika vad gäller storlek, molekylvikt (från 0,05x106 till 4x106) och stabilitet.
De utgör cirka 2 % av den totala mängden RNA i cellen. Alla mRNA är bärare av genetisk information från kärnan till cytoplasman, till platsen för proteinsyntesen. De fungerar som en matris (arbetsritning) för syntesen av en proteinmolekyl, eftersom de bestämmer aminosyrasekvensen (primär struktur) för en proteinmolekyl.

- Ribosomalt RNA (rRNA).
De utgör 80–85 % av det totala RNA-innehållet i cellen.
31. Strukturen av RNA. RNA-typer, strukturella egenskaper och funktioner. Sekundär struktur av tRNA
Ribosomalt RNA består av 3–5 tusen nukleotider. Det syntetiseras i kärnan i kärnan. I komplex med ribosomala proteiner bildar rRNA ribosomer - organeller på vilka proteinmolekyler är sammansatta. Den huvudsakliga betydelsen av rRNA är att det tillhandahåller den initiala bindningen av mRNA och ribosom och bildar det aktiva centrumet av ribosomen, där peptidbindningar bildas mellan aminosyror under syntesen av polypeptidkedjan.
- Överför RNA (tRNA).
tRNA-molekyler innehåller vanligtvis 75-86 nukleotider. Molekylvikten för tRNA-molekyler är cirka 25 tusen. tRNA-molekyler spelar rollen som mellanhänder i proteinbiosyntes - de levererar aminosyror till platsen för proteinsyntes, det vill säga till ribosomer. Cellen innehåller mer än 30 typer av tRNA. Varje typ av tRNA har sin egen unika nukleotidsekvens.
Men alla molekyler har flera intramolekylära komplementära regioner, på grund av närvaron av vilka alla tRNA har en tertiär struktur som liknar ett klöverblad i form.
Sekundär struktur av RNA- karakteristisk för tRNA, enkelsträngad, formad som ett "klöverblad".
Inkluderar:
- relativt korta dubbla helixar - stjälkar,
- enkelsträngade sektioner - slingor.
Det finns 4 stammar (acceptor, antikodon, dihydrouridyl, pseudouridyl) och 3 slingor.

"Stam-loop" - ett element i den sekundära strukturen av RNA, schematiskt

"Pseudoknot" - ett element i den sekundära strukturen av RNA, schematiskt

Acceptorstammen innehåller 3'- och 5'-ändarna av polynukleotidkedjan, 5'-änden med en guanilinsyrarest, 3'-änden är en CCA-triplett och tjänar till att bilda en esterbindning med AA.
Antikodonstammen känner igen sitt kodon på mRNA i ribosomer genom komplementaritetsprincipen.
Pseudouridylstammen tjänar till att fästa vid ribosomen.
Dihydrouridylstammen tjänar till att binda till aminoacyl-tRNA-syntetaset.
Sociala knappar för Joomla
Vi rekommenderar också
 Byte av strömförsörjning: reparation och förfining
Byte av strömförsörjning: reparation och förfining
 Fjärrkontroll av ljus
Fjärrkontroll av ljus
 Simlektioner för förskolebarn
Simlektioner för förskolebarn
 Anteckningar för befälhavaren - hemhushållslarm
Anteckningar för befälhavaren - hemhushållslarm
 Klockpropeller på Atmega8
Klockpropeller på Atmega8
 Applikationsexempel på enheter och reläer, hur man väljer och ansluter ett relä korrekt Mikrokontroller och relä enkla kopplingskretsar
Applikationsexempel på enheter och reläer, hur man väljer och ansluter ett relä korrekt Mikrokontroller och relä enkla kopplingskretsar