Regiunile funcționale ale ARNt. Structura și funcțiile ARNt, caracteristicile activării aminoacizilor
70-90N | pagina secundară - trifoi | CCA 3" const pentru tot ARNt |
prezența timinei, pseudouridină-psi, digirouridină DGU în bucla D - protecție împotriva ribonucleazelor? longeviv | O varietate de structuri primare ale ARNt - 61 + 1 - prin numărul de codoni + ARNt formilmetionină, anticodonul pisicii este același cu cel al ARNt metioninei. Varietate de structuri terțiare - 20 (în funcție de numărul de aminoacizi) | recunoaștere - formarea unei legături covalente m-y tARN și act | aminoacil-ARNt sintetazele atașează acte la tARN
Funcția ARNt este de a transfera aminoacizi din citoplasmă la ribozomi, în care are loc sinteza proteinelor.
ARNt-urile care leagă un aminoacid se numesc izoacceptor.
În total, 64 de ARNt diferite există simultan într-o celulă.
Fiecare ARNt se asociază numai cu propriul codon.
Fiecare ARNt își recunoaște propriul codon fără implicarea unui aminoacid. Aminoacizii legați de ARNt au fost modificați chimic, după care a fost analizată polipeptida rezultată, care conținea aminoacidul modificat. Cysteinil-tRNACys (R=CH2-SH) a fost redus la alanil-tRNACys (R=CH3).
Majoritatea ARNt-urilor, indiferent de secvența lor de nucleotide, au o structură secundară în formă de trifoi datorită prezenței a trei ac de păr în ea.
Caracteristicile structurale ale ARNt
Există întotdeauna patru nucleotide nepereche la 3 "capătul moleculei, iar trei dintre ele sunt în mod necesar CCA. Capetele 5" și 3" ale lanțului de ARN formează o tulpină acceptor. Lanțurile sunt ținute împreună datorită împerecherii complementare a șapte nucleotide 5 "- se termină cu șapte nucleotide situate în apropierea capătului de 3 ". 2. Toate moleculele au un ac de păr T? C, denumit astfel deoarece conține două reziduuri neobișnuite: ribotimidină (T) și pseudouridină (? Acul de păr este format dintr-un ac dublu -tulpină de cinci perechi de baze, inclusiv o pereche de GC, și o buclă de șapte nucleotide în lungime.
în același punct al buclei. 3. Într-un ac de păr anticodon, tulpina este întotdeauna reprezentată de o familie de perechi
temeiuri. Tripletul complementar codonului înrudit, anticodonul, este situat în buclă.
le, constând din șapte nucleotide. O ura invariantă-
cyl și o citozină modificată, iar o purină modificată se învecinează cu capătul său de 3 ", de regulă
adenina. 4. Un alt ac de păr constă dintr-o tulpină lungă de trei până la patru perechi de nucleotide și o buclă variabilă
dimensiune, care conțin adesea uracil într-o formă redusă - dihidrouracil (DU). Secvențele de nucleotide ale tulpinilor, numărul de nucleotide dintre tulpina anticodonului și tulpina T?C (bucla variabilă), precum și dimensiunea buclei și localizarea reziduurilor de dihidrouracil în bucla DU variază cel mai puternic.
[Singer, 1998].
Structura terțiară a ARNt
Structură în formă de L.
Atașarea aminoacizilor la ARNt
Pentru ca un aminoacid să formeze un lanț polipeptidic, acesta trebuie să fie atașat la ARNt de către enzima aminoacil-ARNt sintetaza. Această enzimă formează o legătură covalentă între gruparea carboxil aminoacidului și gruparea hidroxil riboză la capătul 3’ al ARNt cu participarea ATP. Aminoacil-ARNt sintetaza recunoaște un codon specific nu datorită prezenței unui anticodon pe ARNt, ci prin prezența unui situs de recunoaștere specific pe ARNt.
În total, există 21 de aminoacil-ARNt sintetaze diferite în celulă.
Aderarea are loc în două etape:
1. Gruparea carboxil a unui aminoacid este atașată de ATP a-fosfat. Adenilatul de aminoacil instabil rezultat este stabilizat prin legarea la enzimă.
2. Transferul grupării aminoacil a adenilatului de aminoacil la gruparea 2’ sau 3’-OH a ribozei terminale a ARNt
Unele aminoacil-ARNt sintetaze constau dintr-un singur lanț polipeptidic, în timp ce altele constau din două sau patru lanțuri identice, fiecare cu o greutate moleculară de 35 până la 115 kDa. Unele enzime dimerice și tetramerice sunt compuse din două tipuri de subunități. Nu există o corelație clară între dimensiunea moleculei de enzimă sau natura structurii și specificității subunității sale.
Specificitatea unei enzime este determinată de legarea sa puternică la capătul acceptor al ARNt, regiunea DU și bucla variabilă. Unele enzime par să nu recunoască tripletul anticodonului și catalizează reacția de aminoacetilare chiar și atunci când anticodonul este alterat. Cu toate acestea, unele enzime prezintă o activitate redusă în raport cu astfel de ARNt modificați și adaugă aminoacidul greșit atunci când înlocuiesc anticodonul.
70-90n | pagina secundară - trifoi | CCA 3" const pentru tot ARNt |
prezența timinei, pseudouridină-psi, digirouridină DGU în bucla D - protecție împotriva ribonucleazelor? longeviv | O varietate de structuri primare ale ARNt - 61 + 1 - prin numărul de codoni + ARNt formilmetionină, anticodonul pisicii este același cu cel al ARNt metioninei. Varietate de structuri terțiare - 20 (în funcție de numărul de aminoacizi)
Există două tipuri de tRNAFMet și tRNAMMet de legare a metioninei la procariote și tRNAIMet și tRNAMMet la eucariote. Se adaugă metionină la fiecare ARNt utilizând sinteza aminoacil-ARNt corespunzătoare. metionina atașată la tRNAFMet și tRNAIMet este formată de enzima metionil-ARNt-transformilază la Fmet-tRNAFMet. ARNt-urile încărcate cu formilmetionină recunosc codonul de inițiere AUG.
Literatură:
Din păcate, nu există bibliografie.
Manual. În ciuda faptului că ARNt este mult mai mic, o poveste despre structura, caracteristicile și funcționarea sa merită un capitol separat.
Deci, ARNt este un „adaptor”, care recunoaște secvența de trei litere a codului genetic la un capăt, potrivindu-l cu singurul aminoacid corespunzător fixat la celălalt capăt al ARNt. La sfârșitul ARN-ului de transfer care atinge ARN-ul mesager, există 3 nucleotide care formează anticodon. Numai dacă anticodonul este complementar regiunii ARNm se poate alătura ARN-ului de transfer. Dar chiar și în acest caz, ARNt nu se poate alătura ARNm-ului singur; are nevoie de ajutorul ribozomului, care este locul interacțiunii lor, precum și de un participant activ la traducere. De exemplu, ribozomul este cel care creează legături între aminoacizii aduși de ARNt, formând un lanț proteic.
Caracteristicile structurale ale ARNt sunt determinate de codul genetic, adică de regulile pentru construirea unei proteine în funcție de o genă pe care o citește ARN-ul de transfer. Acest cod funcționează în fiecare creatură vie de pe Pământ: crearea unui virus este scrisă în aceiași codoni de trei litere care sunt folosiți pentru a scrie „instrucțiunile de asamblare” ale unui delfin. S-a verificat experimental că genele unei creaturi vii, plasate în celula alteia, sunt perfect copiate și traduse în proteine care nu se pot distinge de genele care se reproduc în celulele gazdei. Uniformitatea codului genetic stă la baza producerii de E. coli modificată de către colonii de insulină și multe alte enzime umane care sunt utilizate ca medicamente pentru persoanele ale căror organisme nu sunt capabile să le producă sau să le producă insuficient. În ciuda diferenței evidente dintre oameni și E. coli, proteinele umane sunt create cu ușurință din planuri umane folosind un copiator E. coli. Nu este surprinzător că ARN-urile de transfer ale diferitelor creaturi diferă foarte puțin.

Fiecare codon din această listă, cu excepția a trei codoni de oprire, care semnalează finalizarea traducerii, ar trebui să fie recunoscut de ARN-ul de transfer. Recunoașterea se realizează prin atașarea unui anticodon la ARN mesager, care se poate lega doar de un codon din listă, astfel încât ARNt poate recunoaște doar un codon. Aceasta înseamnă că există cel puțin 61 de tipuri de aceste molecule în celulă. De fapt, sunt chiar mai multe, deoarece în unele situații pentru citirea ARN-ului mesager nu este suficient doar să existe anticodonul potrivit: sunt necesare alte condiții, în conformitate cu care se creează un ARNt special, modificat.
La prima vedere, o astfel de varietate de ARNt ar trebui să complice semnificativ procesul de traducere: la urma urmei, fiecare dintre aceste molecule va verifica codonul ARN-ului matricei substituit de ribozom pentru conformitatea cu anticodonul său - s-ar părea că atât de multă muncă mecanică inutilă. , atât de mult timp și energie pierdute. Dar ca urmare a evoluției s-au format și mecanisme celulare care previn această problemă. De exemplu, cantitatea de ARNt a fiecărei specii dintr-o celulă corespunde cu cât de des se găsește aminoacidul transportat de acea specie în proteinele construite. Există aminoacizi care sunt rar utilizați de celulă și există aceia care sunt adesea folosiți, iar dacă numărul de ARNt care îi poartă ar fi același, acest lucru ar complica foarte mult asamblarea proteinelor. Prin urmare, există puțini aminoacizi „rari” și ARNt-urile corespunzătoare lor în celulă, în timp ce aminoacizii care apar frecvent sunt produși în cantități mari.

Cu o asemenea varietate de molecule de ARNt, toate sunt foarte asemănătoare, prin urmare, având în vedere structura și funcțiile lor, vom studia în principal caracteristicile comune tuturor speciilor. Când te uiți la aspectul 3D al ARNt, arată ca o grămadă densă de atomi. Pare incredibil că această moleculă complicat încolăcită este rezultatul plierii unui lanț lung de nucleotide, dar așa se formează.
Este posibil să se urmărească etapele formării sale, începând de la prima: compilarea unei secvențe de nucleotide de către ARN polimerază în conformitate cu gena care conține informații despre acest ARN de transfer. Se numește ordinea în care aceste nucleotide se succed și numărul lor structura primară a ARNt. Se pare că este structura primară a ARNt care este codificată în gena citită de ARN polimerază. În general, structura primară este o secvență de molecule relativ simple de același tip, din care este compusă o moleculă de polimer mai complexă, pliată. De exemplu, structura primară a unei molecule de proteine este secvența simplă a aminoacizilor ei constituenți.

Orice lanț de nucleotide nu poate fi în stare desfășurată într-o celulă, pur și simplu întins într-o linie. Există prea multe părți încărcate pozitiv și negativ la marginile nucleotidelor, care formează cu ușurință legături de hidrogen între ele. Modul în care se formează aceleași legături între nucleotidele a două molecule de ADN, conectându-le într-o dublă helix, este descris în, iar pentru detalii despre legăturile de hidrogen, puteți urca în. Legăturile de hidrogen sunt mai puțin puternice decât legăturile dintre atomi din molecule, dar sunt suficiente pentru a răsuci în mod fantezist catena de ARNt și a o menține în acea poziție. La început, aceste legături se formează numai între unele nucleotide, pliind ARNt-ul într-o formă de frunză de trifoi. Rezultatul acestei plieri inițiale se numește structura secundara ARNt. Diagrama din stânga arată că doar unele nucleotide sunt legate prin legături de hidrogen, în timp ce altele rămân nepereche, formând inele și bucle. Diferențele dintre structura secundară a diferitelor tipuri de ARNt se datorează diferențelor în structura lor primară. Acest lucru se manifestă prin lungimi diferite de „frunze de trifoi” sau „tulpină” din cauza lungimii diferite ale lanțului inițial de nucleotide.
O altă diferență în structura primară a diferitelor ARNt-uri este că numai în unele poziții au aceleași nucleotide (în diagrama de mai sus sunt marcate cu primele litere ale numelui lor), în timp ce majoritatea nucleotidelor din diferite ARNt-uri diferă unele de altele. Schema de mai sus este comună tuturor ARNt-urilor, astfel încât diferite nucleotide sunt marcate cu numere.
Principalele părți funcționale ale ARNt sunt:
=) anticodon, adică secvența de nucleotide care este complementară unui singur codon de ARN mesager situat pe ac de păr anticodon
=) capăt acceptor de care se poate atasa un aminoacid. Este situat pe partea opusă a acului de păr anticodon.
În realitate, nici un ARNt nu arată ca în diagrama structurii secundare, deoarece doar unele nucleotide s-au unit pentru a-l forma, în timp ce restul au rămas nepereche. Datorită formării legăturilor de hidrogen între nucleotide din diferite părți ale frunzei de trifoi, se pliază mai mult într-o formă mult mai complexă. structura tertiaraîn formă de L. Puteți înțelege exact cum s-au curbat diferitele părți ale structurii secundare pentru a forma structura terțiară prin potrivirea culorilor din diagramele lor de mai jos. Actorul de păr anticodon, marcat cu albastru și gri, rămâne în partea de jos (merită să ne amintim că acest „de jos” este condiționat: este convenabil să descrii ARNt în această orientare spațială în schemele de traducere a proteinelor) și capătul acceptor (galben) este îndoit în lateral.

Așa arată ARNt-ul când este gata să atașeze un aminoacid. ARNt nu se poate combina singur cu aminoacidul, aceasta necesită participarea unei enzime speciale: aminoacil-ARNt sintetaze. Numărul de tipuri de sintetaze dintr-o celulă coincide cu numărul de tipuri de ARNt.
Uniformitatea formei tuturor tipurilor de ARNt este necesară pentru ca ribozomul să poată recunoaște oricare dintre ele, să le faciliteze andocarea cu ARNm și să se deplaseze în interiorul său de la un loc la altul. Dacă diferitele tipuri de ARNt ar fi semnificativ diferite unele de altele, acest lucru ar face munca ribozomului extrem de dificilă, reducând în mod critic rata sintezei proteinelor. Selecția naturală urmărește astfel să facă ARNt similare între ele. Dar, în același timp, există un alt factor care necesită existența unor diferențe vizibile între diferitele tipuri de ARNt: la urma urmei, este necesar să se recunoască fiecare tip și să se atașeze la el singurul aminoacid corespunzător. Evident, aceste diferențe ar trebui să fie sesizabile, dar nu prea semnificative, astfel încât munca de recunoaștere a speciilor de ARNt să se transforme într-un proces de bijuterii. Și tocmai acest lucru este realizat de sintetazele aminoacil-ARNt: fiecare dintre ele se poate lega doar de unul dintre cei 20 de aminoacizi și se poate atașa exact la acele tipuri de ARNt care corespund acestui aminoacid. Din tabelul cu codul genetic, se poate observa că fiecare aminoacid este codificat de mai multe secvențe de nucleotide, prin urmare, de exemplu, toate cele patru ARNt cu anticodoni CGA, CGG, CGU și CGC vor fi recunoscute de aceeași sintetază care leagă alanina. lor. Astfel de ARNt procesate de o sintetază sunt numite legate de.
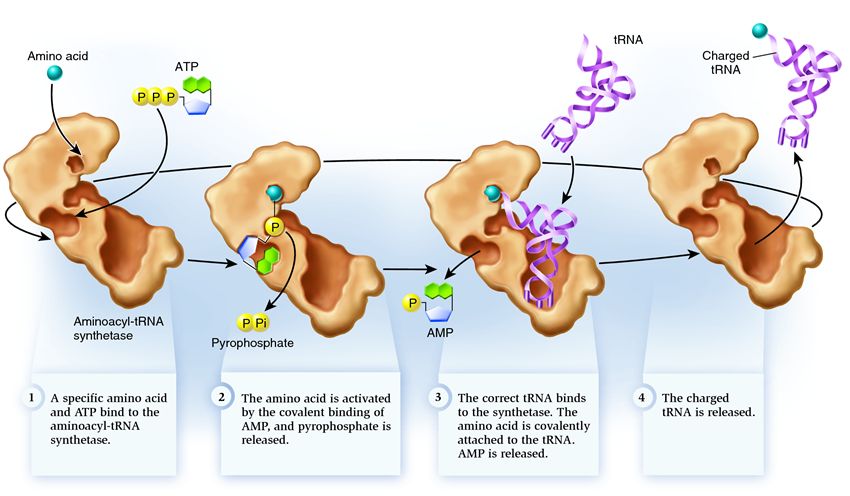
Sintetaza aparține unui grup de enzime a căror funcție este de a se lega de moleculele existente separat și de a le combina într-una singură:
1 . sintetaza conectează un aminoacid și o moleculă de ATP. Două grupuri de fosfat se desprind de ATP, eliberând energia necesară pentru următoarele activități. Adenozin monofosfatul (AMP) rămas din molecula distrusă se atașează de aminoacid, pregătindu-l pentru conectarea cu actorul de păr acceptor.
2 . sintetaza se atașează la sine unul dintre ARNt-urile înrudite corespunzător acestui aminoacid.

În această etapă, se verifică conformitatea ARN-ului de transfer cu sintetaza. Există mai multe moduri de recunoaștere și fiecare sintetază are o combinație unică a acestora. Cel puțin o nucleotidă anticodon este implicată în interacțiunea dintre sintetază și ARNt. Actorul de păr acceptor trebuie, de asemenea, verificat: se determină prezența nucleotidelor specifice pe acesta care sunt comune ARNt-urilor înrudite corespunzătoare aminoacidului dorit. Nucleotidele din alte părți ale ARNt pot participa, de asemenea, la potrivire prin legarea la anumite situsuri sintetaze. ARNt greșit se poate potrivi cu cel dorit într-un fel, dar din cauza potrivirii incomplete, se va alătura sintetazei încet și lejer, căzând ușor. Și ARNt-ul corect se va lipi rapid și ferm de sintetază, drept urmare structura sintetazei se modifică, demarând procesul aminoacilare , adică atașarea unui aminoacid la ARNt.

3 . aminoacilarea constă în înlocuirea moleculei de AMP atașată aminoacidului cu o moleculă de ARNt. După această înlocuire, AMP părăsește sintetaza și ARNt este reținut pentru o ultimă verificare a aminoacizilor. Dacă aminoacidul atașat este recunoscut ca fiind incorect, acesta va fi detașat din ARNt, locul aminoacidului în sintetază va fi gol și o altă moleculă se poate alătura acolo. Noul aminoacid va trece prin etapele de conectare cu ATP și ARNt și va fi, de asemenea, testat. Dacă nu s-au făcut greșeli, ARNt-ul încărcat cu aminoacizi este eliberat: este gata să-și joace rolul în traducerea proteinelor. Și sintetaza este gata să atașeze noi aminoacizi și ARNt, iar ciclul va începe din nou.
Multe depind de funcționarea corectă a aminoacil-ARNt sintetazei: dacă apare o defecțiune în această etapă, atunci aminoacidul greșit va fi atașat la tARN. Și va fi încorporat în proteina care crește pe ribozom, deoarece ARNt și ribozomul nu au funcția de a verifica corespondența codonului și aminoacidului. Consecințele erorii pot fi minore sau catastrofale, iar prin selecție naturală, creaturile cu enzime care nu au funcția unor astfel de verificări au fost înlocuite cu altele mai adaptabile, cu diverse opțiuni de potrivire între aminoacid și ARNt. Prin urmare, în celulele moderne, sintetaza se combină cu aminoacidul greșit în medie într-un caz din 50 de mii și cu ARNt greșit doar o dată la 100 de mii de atașamente.

Unii aminoacizi diferă între ei doar prin câțiva atomi. Dacă te uiți la schemele lor, devine evident că probabilitatea de a confunda arginina cu alanina este mult mai mică decât confuzia izoleucinei cu leucină sau valină. Prin urmare, fiecare sintetază care se leagă de unul dintre aminoacizii asemănători unul cu celălalt are mecanisme suplimentare de verificare. Iată un exemplu de astfel de adaptare în izoleucină-ARNt sintetaza:

Fiecare sintază are centru sinteticîn care un aminoacid este atașat de un ARNt. Actorul de păr acceptor al ARNt capturat de sintetază merge acolo, la fel ca și aminoacidul care este gata să reacționeze cu acesta. Lucrarea unor sintetaze se termină imediat după conectarea aminoacidului cu ARNt. Dar Ile-ARNt sintetaza are o șansă crescută de a face greșeli datorită existenței altor aminoacizi asemănătoare izoleucinei. Prin urmare, și ea are centru corecţional: din denumire este clar ce rol joacă în procesul de conectare a ARNt și a aminoacizilor. Figura din dreapta arată că poziția capătului acului de păr acceptor ARNt în centrul sintetic al sintetazei Ile-ARNt conferă acestui ac de păr o îndoire nenaturală. Cu toate acestea, sintetaza menține ARNt-ul în această poziție până când aminoacidul este atașat de acesta. După ce a apărut această conexiune, nevoia de a găsi ac de păr acceptor în centrul sintetic este epuizată, iar ARNt-ul se îndreaptă, ajungând la capătul său cu aminoacidul atașat în centrul de corecție.

Desigur, centrul sintetic joacă, de asemenea, un rol în eliminarea aminoacizilor care nu sunt potriviți pentru sintetază. Pentru a intra în ea, molecula trebuie să îndeplinească o serie de condiții, inclusiv să aibă dimensiunea potrivită. În ciuda faptului că leucina și izoleucina conțin același număr de atomi, din cauza diferențelor în structura spațială, leucina este mai mare. Prin urmare, nu poate pătrunde în centrul sintetic, a cărui dimensiune corespunde izoleucinei mai compacte, și pur și simplu sare în Ile-ARNt sintetaza.
Dar valina, care este cea mai mică dintre aceste trei molecule cu o structură atomică similară, ia cu ușurință locul izoleucinei în centrul sintetic, iar sintetaza o atașează de ARNt. În acest caz intră în joc centrul de corecție al sintetazei. Dacă actorul de păr acceptor de îndreptare este încărcat corect și poartă izoleucină, atunci nu se poate strânge în interiorul centrului de corecție: este pur și simplu prea mic pentru această moleculă. Astfel, ARNt-ul îndreptat nu mai este ținut de nimic și este detașat de sintetază. Dar dacă valina este atașată la ARNt, ea se strecoară în centrul de corecție, menținând astfel ARNt-ul conectat la acesta în sintetază. O astfel de ședere excesiv de lungă a ARNt în interior este un semnal de eroare pentru sintetază, schimbându-i configurația spațială. Ca rezultat:
=) valină este detașată de ARNt și îndepărtată din sintetază
=) Actorul de păr acceptor revine la locul sintetic, așteptând atașarea la aminoacid
=) sintetaza se leagă de un nou aminoacid, „încarcă” tARN cu acesta și verifică din nou dacă izoleucina a fost folosită pentru aceasta.
Un mecanism similar de recunoaștere dublă este utilizat de alte sintetaze.
Proprietățile fizice și chimice ale ADN-ului
Diverși factori care rup legăturile de hidrogen (creșterea temperaturii peste 80 C, modificări ale pH-ului și ale forței ionice, acțiunea ureei etc.) provoacă denaturarea ADN-ului, adică. modificarea aranjamentului spațial al lanțurilor de ADN fără a rupe legăturile covalente. Helixul dublu al ADN-ului în timpul denaturării este împărțit complet sau parțial în lanțurile sale componente. Denaturarea ADN-ului este însoțită de o creștere a absorbției optice în regiunea UV a bazelor purinice și pirimidinice. Acest fenomen se numește efect hipercromic . Denaturarea reduce, de asemenea, vâscozitatea ridicată inerentă soluțiilor de ADN nativ. Când structura originală a ADN-ului dublu catenar este restabilită, ca urmare a renaturarii, absorbția la 260 nm de către bazele azotate scade datorită „protejării” acestora. Acest fenomen se numește efect hipocrom .
„Desfășurarea” fiecărui ADN în lanțurile sale componente se realizează într-un anumit interval de temperatură. Punctul de mijloc al acestui interval se numește punct de topire. Temperatura de topire a ADN-ului depinde în condiții standard (un anumit pH și putere ionică) de raportul bazelor azotate. Perechile G-C care conțin trei legături de hidrogen sunt mai puternice, prin urmare, cu cât conținutul de perechi G-C în ADN este mai mare, cu atât este mai mare punctul de topire.
Funcțiile ADN-ului. În secvența de nucleotide din moleculele de ADN, informația genetică este codificată. Principalele funcții ale ADN-ului sunt, în primul rând, de a asigura reproducerea lui însuși într-o serie de generații de celule și generații de organisme și, în al doilea rând, de a asigura sinteza proteinelor. Aceste funcții se datorează faptului că moleculele de ADN servesc ca matrice în primul caz pentru replicare, adică. copierea informațiilor în moleculele de ADN fiice, în al doilea - pentru transcriere, adică. pentru a recoda informațiile în structura ARN.
Orez. 5 Curba de topire (denaturarea ADN)
Catenele complementare de ADN separate în timpul denaturarii pot, în anumite condiții, să se reconecteze într-o dublă helix. Acest proces se numește RENATURARE. Dacă denaturarea nu a avut loc complet și cel puțin câteva baze nu și-au pierdut interacțiunea prin legăturile de hidrogen, renaturarea are loc foarte repede.
Citoplasma celulelor conține trei tipuri funcționale principale de ARN. Acestea sunt ARN-uri mesager - ARNm care acționează ca șabloane pentru sinteza proteinelor, ARN ribozomal - ARNr care acționează ca componente structurale ale ribozomilor și ARN de transfer - ARNt implicați în traducerea (traducerea) informațiilor ARNm în secvența de aminoacizi din proteină.
Tabelul 2 arată diferențele dintre ADN și ARN în ceea ce privește structura, localizarea în celulă și funcțiile.
Tabelul 2 Diferențele dintre ADN și ARN
ARN de transfer, ARNt-acid ribonucleic, a cărui funcție este de a transporta AA la locul de sinteză a proteinelor. Are o lungime tipică de 73 până la 93 de nucleotide și o dimensiune de aproximativ 5 nm. ARNt-urile sunt, de asemenea, direct implicate în creșterea lanțului polipeptidic, alăturându-se - fiind într-un complex cu un aminoacid - codonului ARNm și asigurând conformarea complexului necesar formării unei noi legături peptidice. Fiecare aminoacid are propriul său ARNt. ARNt este un ARN monocatenar, dar în forma sa funcțională are o conformație cu frunză de trifoi. AA se atașează covalent la capătul de 3" al moleculei folosind enzima aminoacil-ARNt sintetază, specifică fiecărui tip de ARNt. La situsul C, există un anticodon corespunzător AA-te. ARNt-urile sunt sintetizate de ARN polimeraza obișnuită în cazul cazului. a procariotelor și de către ARN polimeraza III în cazul eucariotelor Transcrierile genelor ARNt sunt supuse procesării în mai multe etape, ceea ce duce la formarea unei structuri spațiale tipice ARNt.
Procesarea ARNt implică 5 pași cheie:
îndepărtarea secvenței de nucleotide lider de 5";
îndepărtarea secvenței 3’-terminale;
adăugarea unei secvențe CCA la capătul de 3";
excizia intronilor (la eucariote și arhei);
modificări ale nucleotidelor individuale.
Transportul ARNt se efectuează de-a lungul unei căi dependente de Ran, cu participarea factorului de transport exportin t, care recunoaște str-ru secundar și terțiar caracteristic al ARNt matur: secțiuni scurte dublu catenare și procesate corect 5 "- și 3" se termină. Acest mecanism asigură că numai ARNt-urile mature sunt exportate din nucleu.
62. Traducere - recunoașterea codonului ARNm
Translația este o sinteză de proteine efectuată de ribozomi din aminoacizi pe un șablon ARNm (sau și ARN). Elementele constitutive ale procesului de traducere: aminoacizi, ARNt, ribozomi, ARNm, enzime pentru aminoacilarea ARNt, factori de translație proteici (factori proteici de inițiere, alungire, terminare - proteine extraribozomale specifice necesare proceselor de translație), surse de energie ATP și GTP , ioni de magneziu (stabilizează structura ribozomului). 20 de aminoacizi sunt implicați în sinteza proteinelor. Pentru ca un aminoacid să-și „recunoaște” locul în viitorul lanț polipeptidic, acesta trebuie să se lege de un ARN de transfer (ARNt) care îndeplinește o funcție de adaptor. ARNt care se leagă de aminoacid recunoaște apoi codonul corespunzător de pe ARNm. Recunoașterea codonului ARNm:
Interacțiunea codon-anticodon se bazează pe principiile complementarității și antiparalelismului:
3'----C - G-A*------5' ARNt anticodon
5'-----G-C-Y*------codon ARNm 3'
Ipoteza oscilării a fost propusă de F. Crick:
Baza 3’ a codonului ARNm are o împerechere nestrictă cu baza 5’ a anticodonului ARNt: de exemplu, Y (ARNm) poate interacționa cu A și G (ARNt)
Unele ARNt se pot asocia cu mai mult de un codon.
63. Caracteristicile elementelor constitutive ale procesului de traducere. Traducerea (traducere-traducere) este procesul de sinteză a proteinelor din aminoacizi pe matricea ARN-ului informațional (de matrice) (ARNm, ARNm) realizat de ribozom.
Sinteza proteinelor este baza vieții celulare. Pentru a efectua acest proces în celulele tuturor organismelor există organite speciale - ribozomi- complexe ribonucleoproteice construite din 2 subunităţi: mari şi mici. Funcția ribozomilor este de a recunoaște trei litere (trei nucleotide) codoni ARNm, comparându-i cu anticodonii ARNt corespunzători purtători aminoaciziși adăugarea acestor aminoacizi la lanțul proteic în creștere. Deplasându-se de-a lungul moleculei de ARNm, ribozomul sintetizează o proteină în conformitate cu informațiile conținute în molecula de ARNm.
Pentru recunoașterea AK-t în celulă, există „adaptoare” speciale, transfera molecule de ARN(ARNt). Aceste molecule în formă de trifoi au un situs (anticodon) complementar unui codon ARNm, precum și un alt situs la care este atașat aminoacidul corespunzător acelui codon. Atașarea aminoacizilor la ARNt se realizează într-o reacție dependentă de energie de către enzimele aminoacil-ARNt sintetaze, iar molecula rezultată se numește aminoacil-ARNt. Astfel, specificitatea translației este determinată de interacțiunea dintre codonul ARNm și anticodonul ARNt, precum și specificitatea sintetazelor aminoacil-ARNt care atașează aminoacizii strict la ARNt-urile lor corespondente (de exemplu, codonul GGU va corespunde unui ARNt care conține anticodonul CCA și numai glicină AK).
ribozom procariot
ARNr 5S și 23S ARNr 16S
34 veverițe 21 veverițe
Ribozomii procarioți au o constantă de sedimentare de 70S, motiv pentru care se numesc particule 70S. Sunt construite din două subunități diferite: subunități 30S și 50S. Fiecare subunitate este un complex de ARNr și proteine ribozomale.
Particula 30S conține o moleculă de ARNr 16S și, în majoritatea cazurilor, o moleculă de proteină din mai mult de 20 de specii (21) . Subunitatea 50S constă din două molecule de ARNr (23S și 5S). Constă din peste 30 de proteine diferite (34), reprezentate de asemenea, de regulă, printr-un singur exemplar. Majoritatea proteinelor ribozomale îndeplinesc o funcție structurală.
ribozom eucariot
5S; ARNr 5,8S și 28S ARNr 18S
cel puțin 50 de proteine cel puțin 33 de proteine
Ribozomul este format din subunități mari și mici. Baza structurii fiecărei subunități este un ARNr pliat complex. Proteinele ribozomilor au fost atașate la schela ARNr.
Coeficientul de sedimentare al unui ribozom eucariot complet este de aproximativ 80 de unități Svedberg (80S), iar coeficientul de sedimentare al subparticulelor sale este de 40S și 60S.
Subunitatea mai mică 40S constă dintr-o moleculă de ARNr 18S și 30-40 molecule de proteine. Subunitatea mare 60S conține trei tipuri de ARNr cu coeficienți de sedimentare de 5S, 5,8S și 28S și proteine 40-50 (de exemplu, ribozomii hepatocitelor de șobolan includ 49 de proteine).
Regiunile funcționale ale ribozomilor
P - situsul peptidil pentru ARNt peptidil
A - situsul aminoacil pentru ARNt aminoacil
E - situs pentru eliberarea ARNt din ribozom
Ribozomul conține 2 situsuri funcționale pentru interacțiunea cu ARNt: aminoacil (acceptor) și peptidil (donator). Aminoacil-ARNt intră în situsul acceptor al ribozomului și interacționează pentru a forma legături de hidrogen între tripleții codon și anticodon. După formarea legăturilor de hidrogen, sistemul avansează 1 codon și ajunge în locul donor. În același timp, un nou codon apare în situsul acceptor liber și aminoacil-t-ARN-ul corespunzător este atașat de acesta.
Ribozomi: structură, funcție
Ribozomii sunt centrii citoplasmatici ai biosintezei proteinelor. Ele constau din subunități mari și mici, care diferă în coeficienții de sedimentare (rata de sedimentare în timpul centrifugării), exprimate în unități de Svedberg - S.
Ribozomii sunt prezenți atât în celulele eucariote, cât și în cele procariote, deoarece îndeplinesc o funcție importantă biosinteza proteinelor. Fiecare celulă conține zeci, sute de mii (până la câteva milioane) din aceste mici organele rotunjite. Este o particulă de ribonucleoproteină rotunjită. Diametrul său este de 20-30 nm. Ribozomul este format din subunități mari și mici, care diferă prin coeficienții de sedimentare (viteza de sedimentare în timpul centrifugării), exprimate în unități Svedberg - S. Aceste subunități sunt combinate în prezența unei catene de m-ARN (matrice, sau informațional, ARN). Un complex dintr-un grup de ribozomi uniți de o singură moleculă de ARNm ca un șir de margele se numește polizom. Aceste structuri sunt fie localizate liber în citoplasmă, fie atașate de membranele ER granulare (în ambele cazuri, sinteza proteinelor se desfășoară activ pe ele).
Polizomii ER granulare formează proteine care sunt excretate din celulă și utilizate pentru nevoile întregului organism (de exemplu, enzime digestive, proteine din laptele matern uman). În plus, ribozomii sunt prezenți pe suprafața interioară a membranelor mitocondriale, unde au, de asemenea, un rol activ în sinteza moleculelor de proteine.
ARN de transfer (ARNt) joacă un rol important în procesul de utilizare a informațiilor ereditare de către celulă. Livrând aminoacizii necesari la locul de asamblare al lanțurilor peptidice, ARNt acționează ca un mediator de translație.
Moleculele de ARNt sunt lanțuri de polinucleotide sintetizate pe secvențe specifice de ADN. Ele constau dintr-un număr relativ mic de nucleotide -75-95. Ca urmare a conexiunii complementare a bazelor care se află în diferite părți ale lanțului polinucleotidic ARNt, acesta capătă o structură asemănătoare cu forma unei frunze de trifoi (Fig. 3.26).
Orez. 3.26. Structura unei molecule tipice de ARNt.
Are patru părți principale care îndeplinesc funcții diferite. acceptor„Tulpina” este formată din două părți terminale conectate complementare ale ARNt. Este format din șapte perechi de baze. Capătul 3’ al acestei tulpini este oarecum mai lung și formează o regiune monocatenară care se termină cu o secvență CCA cu o grupare OH liberă. În acest scop este atașat un aminoacid transportabil. Celelalte trei ramuri sunt secvențe de nucleotide pereche complementare care se termină în secțiuni nepereche care formează bucle. Mijlocul acestor ramuri - anticodon - este format din cinci perechi de nucleotide și conține un anticodon în centrul buclei sale. Antidonul este trei nucleotide complementare codonului ARNm, care codifică aminoacidul transportat de acest ARNt la locul sintezei peptidei.
Între ramurile acceptor și anticodon sunt două ramuri laterale. În buclele lor, ele conțin baze modificate - dihidrouridină (bucla D) și tripletul TψC, unde \y este pseudouriaină (bucla T^C).
Între ramurile aiticodonă și T^C există o buclă suplimentară, care include de la 3-5 până la 13-21 nucleotide.
În general, diferitele tipuri de ARNt sunt caracterizate de o anumită constanță a secvenței de nucleotide, care constă cel mai adesea din 76 de nucleotide. Variația numărului lor se datorează în principal modificării numărului de nucleotide din bucla suplimentară. Regiunile complementare care susțin structura ARNt sunt de obicei conservate. Structura primară a ARNt, determinată de secvența nucleotidelor, formează structura secundară a ARNt, care are forma unei frunze de trifoi. La rândul său, structura secundară determină o structură terțiară tridimensională, care se caracterizează prin formarea a două elice duble perpendiculare (Fig. 3.27). Una dintre ele este formată din ramurile acceptor și TψC, cealaltă din ramurile anticodon și D.
La capătul uneia dintre helixele duble se află aminoacidul transportat, la capătul celeilalte se află anticodonul. Aceste zone sunt cele mai îndepărtate unele de altele. Stabilitatea structurii terțiare a ARNt este menținută datorită apariției unor legături suplimentare de hidrogen între bazele lanțului polinucleotidic, situate în diferite părți ale acestuia, dar apropiate spațial în structura terțiară.
Diferite tipuri de ARNt au o structură terțiară similară, deși cu unele variații.
Orez. 3.27. Organizarea spațială a ARNt:
I - structura secundară a ARNt sub forma unei „frunze de trifoi”, determinată de structura sa primară (secvența nucleotidelor din lanț);
II - proiecția bidimensională a structurii terțiare a ARNt;
III - dispunerea moleculei de ARNt în spațiu
ANEXĂ (în cazul în care cineva nu înțelege acest lucru)
Dinții de fulger – nucleotide (Adenină-Timină/Uracil/, Guanine-Citazină). Toate fulgerele sunt ADN.

Pentru a transfera informații din ADN, trebuie să rupeți 2 fire. Legătura dintre A-T și G-C este hidrogen, prin urmare este ușor ruptă de enzima helicază:

Pentru a preveni formarea nodurilor (De exemplu, am răsucit un prosop):



Topoizomeraza taie o catenă de ADN la originea replicării, astfel încât lanțul să nu se răsucească.

Când un fir este liber, al doilea se poate roti cu ușurință în jurul axei sale, eliberând astfel tensiunea în timpul „desfășurării”. Nodurile nu apar, se economisește energia.
Apoi, este necesar un primer ARN pentru a începe colectarea ARN. O proteină care asamblează ARNm nu poate asambla doar prima nucleotidă, are nevoie de o bucată de ARN pentru a începe (este scris în detaliu acolo, o voi scrie mai târziu). Această piesă se numește primer ARN. Și această proteină atașează deja prima nucleotidă la ea.


La descrierea structurii acizilor nucleici se iau în considerare diferite niveluri de organizare a macromoleculelor: structura primară și secundară.
Structura primară a acizilor nucleici este compoziția de nucleotide și o anumită secvență de unități de nucleotide din lanțul polimeric.
Structura secundară a ARN-ului. Molecula de acid ribonucleic este construită dintr-un singur lanț polinucleotidic.
Structura secundară a ARN-ului
Secțiuni separate ale lanțului de ARN formează bucle spiralizate - „agrafe de păr”, datorită legăturilor de hidrogen dintre bazele azotate complementare A-U și G-C. Secțiunile lanțului de ARN din astfel de structuri elicoidale sunt antiparalele, dar nu întotdeauna complet complementare; ele conțin resturi de nucleotide nepereche sau chiar bucle monocatenar care nu se potrivesc în dubla helix. Prezența regiunilor spiralizate este caracteristică tuturor tipurilor de ARN.
Rolul principal al ARN-ului este participarea directă la biosinteza proteinelor.
Sunt cunoscute trei tipuri de ARN celular, care diferă prin localizarea lor în celulă, compoziție, dimensiune și proprietăți care determină rolul lor specific în formarea macromoleculelor proteice:
- ARN-urile informaționale (matriceale) transmit informații codificate în ADN despre structura proteinei de la nucleul celular la ribozomi, unde se realizează sinteza proteinelor; structura primară a tuturor ARNm-urilor, indiferent de unicitatea secvenței lor de codare, are aceeași structură a capetelor 5’ și 3’.
Deci, la capătul 5’ există o nucleotidă modificată 7-metilguanozin-5’-trifosfat (capac). Câteva zeci de nucleotide separă capacul de codonul de inițiere, de obicei tripletul -AUG-. Regiunea de codificare este urmată de unul dintre codonii de terminare -UGA-, -UUA-, -UAG-. La capătul 3’ al majorității ARNm, există o secvență de nucleotide de 100-200 de resturi de adenozin monofosfat.
- ARN-urile de transfer colectează aminoacizi în citoplasma celulei și îi transferă în ribozom; Moleculele de ARN de acest tip „învață” din secțiunile corespunzătoare ale lanțului de ARN mesager care aminoacizi ar trebui să participe la sinteza proteinelor.
Structura spațială a oricărui ARNt, indiferent de diferențele în secvența de nucleotide, este descrisă de modelul universal de trifoi. Fiecare moleculă de ARNt are secțiuni de lanț care nu sunt implicate în formarea legăturilor de hidrogen între resturile de nucleotide.
Acestea includ, în special, situsul responsabil pentru legarea de aminoacid la capătul 3’ al moleculei și anticodon, un triplet specific de nucleotide care interacționează complementar cu codonul ARNm.
- ARN-ul ribozomal asigură sinteza proteică a unei anumite structuri, citind informații din ARN (matrice) informațional. ARNr formează complexe cu proteine numite ribozomi.
Fiecare ribozom este format din două subunități - mici (40S) și mari (60S). Subunitățile ribozomilor diferă nu numai prin setul de ARNr, ci și prin numărul și structura proteinelor.
Data publicării: 2015-02-03; Citește: 2729 | Încălcarea drepturilor de autor ale paginii

ARN este un polimer ai cărui monomeri sunt ribonucleotide.
Spre deosebire de ADN, ARN-ul este format nu din două, ci dintr-un singur lanț de polinucleotide (excepție - unii virusuri care conțin ARN au ARN dublu catenar). Nucleotidele ARN sunt capabile să formeze legături de hidrogen între ele. Lanțurile de ARN sunt mult mai scurte decât lanțurile de ADN.
Monomerul ARN - nucleotidă (ribonucleotidă) - constă din reziduuri a trei substanțe: 1) o bază azotată, 2) o monozaharidă cu cinci atomi de carbon (pentoză) și 3) acid fosforic. Bazele azotate ale ARN aparțin și ele claselor de pirimidine și purine.
Bazele pirimidinice ale ARN - uracil, citozină, baze purinice - adenină și guanină.
31. Tipuri de ARN și caracteristici ale structurii sale
Monozaharida nucleotidă ARN este reprezentată de riboză.
Există trei tipuri de ARN: 1) informație (matrice) ARN - ARNm (ARNm), 2) ARN de transfer - ARNt, 3) ARN ribozomal - ARNr.
Toate tipurile de ARN sunt polinucleotide neramificate, au o conformație spațială specifică și participă la procesele de sinteză a proteinelor.
Informațiile despre structura tuturor tipurilor de ARN sunt stocate în ADN. Procesul de sinteză a ARN pe un șablon de ADN este denumit în mod obișnuit transcripție.

ARN-urile de transfer conțin de obicei 76 (de la 75 la 95) nucleotide; greutate moleculară - 25.000–30.000.
ARNt reprezintă aproximativ 10% din conținutul total de ARN din celulă. Funcțiile ARNt: 1) transportul aminoacizilor la locul sintezei proteinelor, la ribozomi, 2) mediator de translație. În celulă se găsesc aproximativ 40 de tipuri de ARNt, fiecare dintre ele având o secvență de nucleotide caracteristică doar pentru aceasta. În același timp, toate ARNt-urile au mai multe regiuni complementare intramoleculare, datorită cărora ARNt-urile capătă o conformație asemănătoare unei frunze de trifoi.
Orice ARNt are o buclă pentru contactul cu ribozomul (1), o buclă anticodon (2), o buclă pentru contactul cu enzima (3), o tulpină acceptor (4) și un anticodon (5). Aminoacidul este atașat la capătul 3’ al tulpinii acceptoare. Anticodon - trei nucleotide care „recunosc” un codon ARNm.
Trebuie subliniat faptul că un anumit ARNt poate transporta un aminoacid strict definit corespunzător anticodonului său. Specificitatea conexiunii dintre aminoacizi și ARNt se realizează datorită proprietăților enzimei aminoacil-ARNt sintetaza.
ARN-urile ribozomale conțin 3000–5000 de nucleotide; greutate moleculară - 1.000.000–1.500.000.
ARNr reprezintă 80-85% din conținutul total de ARN din celulă. În complex cu proteinele ribozomale, ARNr formează ribozomi - organele care realizează sinteza proteinelor. În celulele eucariote, sinteza ARNr are loc în nucleol. Funcțiile ARNr: 1) o componentă structurală necesară a ribozomilor și, astfel, asigurarea funcționării ribozomilor; 2) asigurarea interacțiunii ribozomului și ARNt; 3) legarea inițială a ribozomului și a codonului inițiator ARNm și determinarea cadrului de citire, 4) formarea centrului activ al ribozomului.
ARN-urile mesager sunt diverse în ceea ce privește conținutul de nucleotide și greutatea moleculară (de la 50.000 la 4.000.000).
Ponderea ARNm reprezintă până la 5% din conținutul total de ARN din celulă. Funcțiile ARNm: 1) transfer de informații genetice de la ADN la ribozomi, 2) o matrice pentru sinteza unei molecule de proteine, 3) determinarea secvenței de aminoacizi a structurii primare a unei molecule de proteine.
Citeste si
ARN este un polimer ai cărui monomeri sunt nucleotide.
Cele trei baze azotate sunt aceleași ca în ADN (adenină, guanină, citozină); al patrulea - uracil - este prezent în molecula de ARN în loc de timină. Nucleotidele ARN conțin riboză în loc de deoxiriboză. În lanțul ARN...
trei tipuri principale de ARN: informativ(ARNm) sau matrice(ARNm), ribozomal(ARNr) și transport(ARNt). Ele diferă prin dimensiunea moleculară și funcție. Toate tipurile de ARN sunt sintetizate pe ADN cu participarea enzimelor - ARN polimeraze. ARN mesager reprezintă 2-3% din tot ARN-ul celular, ribozomal - 80-85, transport - aproximativ 15%.
ARNm.
citește informații ereditare dintr-o regiune ADN și, sub forma unei secvențe copiate de baze azotate, o transferă la ribozomi, unde este sintetizată o anumită proteină. Fiecare dintre moleculele de ARNm în ordinea nucleotidelor și ca mărime corespunde genei din ADN din care a fost transcris. În medie, ARNm conține 1500 de nucleotide (75-3000). Fiecare triplet (trei nucleotide) de pe un ARNm se numește codon. Depinde de codon care aminoacid va apărea într-un loc dat în timpul sintezei proteinelor.
(ARNt) are o greutate moleculară relativ mică de aproximativ 24-29 mii.
D și conține de la 75 la 90 de nucleotide în moleculă. Până la 10% din toate nucleotidele ARNt sunt baze minore, ceea ce, aparent, îl protejează de acțiunea enzimelor hidrolitice.Rolul ARNt este acela de a transfera aminoacizi la ribozomi și de a participa la procesul de sinteză a proteinelor. Fiecare aminoacid se atașează de un ARNt specific. O serie de aminoacizi au mai mult de un ARNt. Până în prezent, au fost descoperite peste 60 de ARNt care diferă în structura lor primară (secvența de bază).
Structura secundară a tuturor ARNt-urilor este prezentată sub forma unei frunze de trifoi cu o tulpină dublu catenară și trei monocatenar). La capătul unuia dintre lanțuri există un situs acceptor - tripletul CCA, de adenină căreia este atașat un aminoacid specific.
(ARNr). Conțin 120-3100 de nucleotide. ARN-ul ribozomal se acumulează în nucleu, în nucleoli.
Proteinele ribozomale sunt transportate la nucleoli din citoplasmă, iar acolo are loc formarea spontană a subparticulelor ribozomale prin combinarea proteinelor cu ARNr-ul corespunzător. Subparticulele ribozomului sunt transportate împreună sau separat prin porii membranei nucleare în citoplasmă. Ribozomi sunt organite cu dimensiunea de 20-30 nm.
Sunt construite din două subparticule de dimensiuni și forme diferite. În anumite etape ale sintezei proteinelor în celulă, ribozomii sunt împărțiți în subparticule.
ARN-ul ribozomal servește ca cadru pentru ribozomi și facilitează legarea inițială a ARNm de ribozom în timpul biosintezei proteinelor.
Întrebarea 6 Legăturile care formează structurile primare și secundare ale ADN-ului și ARN-ului. Tipuri de ARN
Codul genetic este o modalitate de codificare a secvenței de aminoacizi a proteinelor folosind o secvență de nucleotide, caracteristică tuturor organismelor vii.
Proprietăți: 1) cod genetic triplet(fiecare aminoacid este codificat de trei nucleotide); 2) nesuprapune(tripleții învecinați nu au nucleotide comune); 3) degenerat(cu excepția metioninei și a triptofanului, toți aminoacizii au mai mult de un codon); 4) universal(în mare parte la fel pentru toate organismele vii); 5) în codoni pentru un aminoacid, primele două nucleotide sunt de obicei aceleași, iar a treia variază; 6) are o ordine liniară de citire și se caracterizează prin coliniaritate, T.
e. coincidenţa ordinului codonilor din ARNm cu ordinea aminoacizilor din lanţul polipeptidic sintetizat.
Data publicării: 2014-12-08; Citește: 11268 | Încălcarea drepturilor de autor ale paginii
studopedia.org - Studopedia.Org - 2014-2018 (0,001 s) ...
Citoplasma celulelor conține trei tipuri principale funcționale de ARN:
- ARN mesager (ARNm) care acționează ca modele pentru sinteza proteinelor;
- ARN ribozomal (ARNr) care acționează ca componente structurale ale ribozomilor;
- transfer ARN-uri (ARNt) implicate în traducerea (traducerea) informațiilor ARNm în secvența de aminoacizi a unei molecule de proteine.
În nucleul celulelor se găsește ARN nuclear, constituind de la 4 până la 10% din totalul ARN celular.
Cea mai mare parte a ARN-ului nuclear este reprezentată de precursori cu molecule înalte ai ARN-ului ribozomal și de transfer. Precursorii ARNr cu greutate moleculară mare (ARN 28 S, 18 S și 5 S) sunt localizați în principal în nucleol.
ARN-ul este principalul material genetic în unele virusuri animale și vegetale (ARN genomic). Majoritatea virusurilor ARN sunt caracterizate prin transcrierea inversă a genomului lor ARN, dirijată de transcriptază inversă.
Toți acizii ribonucleici sunt polimeri ai ribonucleotidelor conectați, ca într-o moleculă de ADN, prin legături 3′,5′-fosforodiester.
Spre deosebire de ADN, care are o structură dublu catenară, ARN-ul este o moleculă polimerică liniară monocatenară.
structura ARNm. ARNm este cea mai eterogenă clasă de ARN în ceea ce privește dimensiunea și stabilitatea.
structura ARNt.
ARN-urile de transfer acționează ca mediatori (adaptori) în timpul traducerii ARNm. Ele reprezintă aproximativ 15% din ARN-ul celular total. Fiecare dintre cei 20 de aminoacizi proteinogeni are propriul său ARNt. Pentru unii aminoacizi codificați de doi sau mai mulți codoni, există mai mulți ARNt.
ARNt-urile sunt molecule monocatenar relativ mici constând din 70-93 nucleotide. Greutatea lor moleculară este (2,4-3,1) .104 kDa.
Structura secundară a ARNt se formează datorită formării numărului maxim de legături de hidrogen între perechile complementare intramoleculare de baze azotate.
Ca rezultat al formării acestor legături, lanțul polinucleotidic ARNt se răsucește cu formarea de ramuri spiralizate care se termină în bucle de nucleotide nepereche. Imaginea spațială a structurilor secundare ale tuturor ARNt-urilor are forma unei foi de trifoi.
Patru ramuri obligatorii se disting în „frunză de trifoi”, ARNt-urile mai lungi, în plus, conțin o a cincea ramură scurtă (suplimentară).
Funcția adaptor a ARNt este asigurată de o ramură acceptor, la capătul 3’ al căreia un rest de aminoacid este atașat printr-o legătură eterică și o ramură de anticodon opusă ramurii acceptoare, în vârful căreia există o buclă care conține un anticodon.
Un anticodon este un triplet specific de nucleotide care este complementar în direcția antiparalelă cu codonul ARNm care codifică aminoacidul corespunzător.
Ramura T care poartă bucla pseudouridină (bucla TyC) asigură interacțiunea ARNt cu ribozomii.
Ramura D, care poartă bucla dehidrouridină, asigură interacțiunea ARNt cu aminoacil-ARNt sintetaza corespunzătoare.

Structura secundară a ARNt
Funcțiile celei de-a cincea ramuri suplimentare sunt încă puțin înțelese; cel mai probabil, echivalează lungimea diferitelor molecule de ARNt.
Structura terțiară a ARNt este foarte compactă și se formează prin reunirea ramurilor individuale ale frunzei de trifoi datorită legăturilor suplimentare de hidrogen pentru a forma o structură de „cot cot” în formă de L.
ARN de transport, structură și mecanism funcțional.
În acest caz, brațul acceptor care leagă aminoacidul este situat la un capăt al moleculei, iar anti-codonul este la celălalt.

Structura terțiară a ARNt (conform A.S. Spirin)
Structura ARNr și a ribozomilor. ARN-urile ribozomale formează coloana vertebrală la care proteinele specifice se leagă pentru a forma ribozomi. Ribozomii sunt organite nucleoproteice care asigură sinteza proteinelor din ARNm.
Numărul de ribozomi dintr-o celulă este foarte mare: de la 104 la procariote la 106 la eucariote. Ribozomii sunt localizați în principal în citoplasmă, la eucariote, în plus, în nucleol, în matricea mitocondrială și în stroma cloroplastelor. Ribozomii constau din două subparticule: mari și mici. După mărime și greutate moleculară, toți ribozomii studiați sunt împărțiți în 3 grupe - ribozomi 70S ai procariotelor (coeficient de sedimentare S), constând din subparticule mici 30S și 50S mari; Ribozomi eucarioți 80S, constând din subunități 40S mici și 60S mari.
Mica subunitate a ribozomilor 80S este formată dintr-o moleculă de ARNr (18S) și 33 de molecule de diferite proteine.
Subunitatea mare este formată din trei molecule de ARNr (5S, 5.8S și 28S) și aproximativ 50 de proteine.
Structura secundară a ARNr se formează datorită secțiunilor scurte dublu catenare ale moleculei - ac de păr (aproximativ 2/3 din ARNr), 1/3 - este reprezentată de secțiuni monocatenar bogate în nucleotide purinice.
Butoane sociale pentru Joomla
Proteinele formează baza vieții. Funcțiile lor în celulă sunt foarte diverse. Cu toate acestea, proteinele „nu se pot” reproduce. Și toate informațiile despre structura proteinelor sunt conținute în gene (ADN).
În organismele superioare, proteinele sunt sintetizate în citoplasma celulei, iar ADN-ul este ascuns în spatele învelișului nucleului. Prin urmare, ADN-ul nu poate servi direct ca șablon pentru sinteza proteinelor. Acest rol este îndeplinit de un alt acid nucleic - ARN.
Molecula de ARN este o polinucleotidă neramificată cu o structură terțiară.
Este format dintr-un lanț de polinucleotide și, deși nucleotidele complementare incluse în el sunt, de asemenea, capabile să formeze legături de hidrogen între ele, aceste legături apar între nucleotidele unui lanț. Lanțurile de ARN sunt mult mai scurte decât lanțurile de ADN. Dacă conținutul de ADN într-o celulă este relativ constant, atunci conținutul de ARN fluctuează foarte mult. Cea mai mare cantitate de ARN din celule este observată în timpul sintezei proteinelor.
ARN-ul joacă un rol major în transmiterea și implementarea informațiilor ereditare.
În conformitate cu funcția și caracteristicile structurale, se disting mai multe clase de ARN celular.
Există trei clase principale de ARN celular.
- Informațional (ARNm) sau matrice (ARNm). Moleculele sale sunt cele mai diverse în ceea ce privește dimensiunea, greutatea moleculară (de la 0,05x106 la 4x106) și stabilitate.
Ele reprezintă aproximativ 2% din cantitatea totală de ARN din celulă. Toate ARNm sunt purtători de informații genetice de la nucleu la citoplasmă, la locul sintezei proteinelor. Ele servesc ca o matrice (desen de lucru) pentru sinteza unei molecule de proteine, deoarece determină secvența de aminoacizi (structura primară) a unei molecule de proteine.

- ARN ribozomal (ARNr).
Ele reprezintă 80-85% din conținutul total de ARN din celulă.
31. Structura ARN-ului. Tipuri de ARN, caracteristici structurale și funcții. Structura secundară a ARNt
ARN-ul ribozomal este format din 3-5 mii de nucleotide. Se sintetizează în nucleolii nucleului. În complex cu proteinele ribozomale, ARNr formează ribozomi - organele pe care sunt asamblate moleculele proteice. Semnificația principală a ARNr este că asigură legarea inițială a ARNm și a ribozomului și formează centrul activ al ribozomului, în care se formează legături peptidice între aminoacizi în timpul sintezei lanțului polipeptidic.
- ARN de transfer (ARNt).
Moleculele de ARNt conțin de obicei 75-86 nucleotide. Greutatea moleculară a moleculelor de ARNt este de aproximativ 25 mii. Moleculele de ARNt joacă rolul de intermediari în biosinteza proteinelor - furnizează aminoacizi la locul sintezei proteinelor, adică la ribozomi. Celula conține mai mult de 30 de tipuri de ARNt. Fiecare tip de ARNt are propria sa secvență unică de nucleotide.
Cu toate acestea, toate moleculele au mai multe regiuni complementare intramoleculare, datorită prezenței cărora toți ARNt-urile au o structură terțiară asemănătoare cu forma unei frunze de trifoi.
Structura secundară a ARN-ului- caracteristic ARNt, monocatenar, în formă de „frunză de trifoi”.
Include:
- elice duble relativ scurte - tulpini,
- secțiuni monocatenare - bucle.
Există 4 tulpini (acceptor, anticodon, dihidrouridil, pseudouridil) și 3 bucle.

„Stem-loop” - un element al structurii secundare a ARN-ului, schematic

„Pseudoknot” - un element al structurii secundare a ARN-ului, schematic

Tulpina acceptor conține capetele 3’ și 5’ ale lanțului polinucleotidic, capătul 5’ cu un rest de acid guanilic, capătul 3’ este un triplet CCA și servește la formarea unei legături esterice cu AA.
Tulpina anticodonului își recunoaște codonul pe ARNm din ribozomi prin principiul complementarității.
Tulpina pseudouridil servește la atașarea ribozomului.
Tulpina dihidrouridil servește la legarea la aminoacil-ARNt sintetaza.
Butoane sociale pentru Joomla
Recomandăm și noi
 Sursă de alimentare comutată: reparație și perfecționare
Sursă de alimentare comutată: reparație și perfecționare
 Telecomanda luminii
Telecomanda luminii
 Lecții de înot pentru copii preșcolari
Lecții de înot pentru copii preșcolari
 Note pentru master - alarme de uz casnic
Note pentru master - alarme de uz casnic
 Elice de ceas pe Atmega8
Elice de ceas pe Atmega8
 Exemple de aplicații pentru dispozitive și relee, cum să alegeți și să conectați corect un releu Microcontroler și circuite simple de comutare a releului
Exemple de aplicații pentru dispozitive și relee, cum să alegeți și să conectați corect un releu Microcontroler și circuite simple de comutare a releului