Mi az átírási folyamat? Az RNS örökletes információkat kap
Az átírás megkezdése
Átírási megnyúlás
Az a pillanat, amikor az RNS-polimeráz a transzkripció iniciációjából az elongációba megy át, nincs pontosan meghatározva. Három jelentősebb biokémiai esemény jellemzi ezt az átmenetet az Escherichia coli RNS polimeráz esetében: a szigma faktor felszabadulása, az enzimmolekula első transzlokációja a templát mentén, valamint a transzkripciós komplex erős stabilizálódása, amely az RNS mellett polimeráz, magában foglalja a növekvő RNS-láncot és az átírt DNS-t. Ugyanezek a jelenségek jellemzőek az eukarióta RNS polimerázokra is. Az iniciációból az elongációba való átmenetet az enzim, a promoter, a transzkripciós iniciációs faktorok közötti kötések felszakadása kíséri, és egyes esetekben az RNS polimeráz elongációs kompetencia állapotba való átmenete (például a CTD domén foszforilációja RNS polimeráz II). Az elongációs fázis a növekvő transzkriptum felszabadulását követően ér véget, és az enzim disszociál a templátról (termináció).
A nyúlás alapvető nyúlási tényezők segítségével történik, amelyek szükségesek ahhoz, hogy a folyamat ne álljon le idő előtt.
A közelmúltban bizonyítékok jelentek meg, amelyek azt mutatják, hogy a szabályozó tényezők is szabályozhatják a megnyúlást. Az elongációs folyamat során az RNS-polimeráz a gén bizonyos részein megáll. Ez különösen jól látható kis szubsztrátumkoncentráció esetén. A mátrix egyes területein hosszú késések tapasztalhatók az RNS polimeráz előrehaladása, az ún. még optimális szubsztrátkoncentráció esetén is szünetek figyelhetők meg. Ezeknek a szüneteknek az időtartama nyúlási tényezőkkel szabályozható.
Felmondás
A baktériumoknak két transzkripciós terminációs mechanizmusa van:
- egy rho-függő mechanizmus, amelyben a Rho (rho) fehérje destabilizálja a DNS-templát és az mRNS közötti hidrogénkötéseket, felszabadítva az RNS-molekulát.
- rho-független, amelyben a transzkripció leáll, amikor az újonnan szintetizált RNS-molekula szárhurkot képez, majd több uracil (...UUUU), ami az RNS-molekula DNS-templátról való leválásához vezet.
A transzkripció terminációját eukariótákban kevésbé tanulmányozták. Az RNS levágásával zárul, ami után az enzim több adenint (...AAAA) ad a 3"-os végéhez, amelyek száma meghatározza az adott transzkriptum stabilitását.
Átírási gyárak
Számos kísérleti adat utal arra, hogy a transzkripció az úgynevezett transzkripciós gyárakban történik: hatalmas, egyes becslések szerint akár 10 Da tömegű komplexek, amelyek körülbelül 8 RNS-polimeráz II-t tartalmaznak, valamint a későbbi feldolgozáshoz és splicinghez, valamint korrekcióhoz szükséges összetevőket. az újonnan szintetizált átiratból. A sejtmagban állandó csere folyik az oldható és az aktivált RNS-polimeráz készletei között. Az aktív RNS-polimeráz részt vesz egy ilyen komplexben, amely viszont egy szerkezeti egység, amely a kromatin tömörítését szervezi. A legfrissebb adatok azt mutatják, hogy transzkripciós gyárak transzkripció hiányában is léteznek, a sejtben rögzülnek (még nem tisztázott, hogy kölcsönhatásba lépnek-e a sejt magmátrixával vagy sem) és önálló nukleáris alkompartmentet képviselnek. Az RNS-polimeráz I-et, II-t vagy III-at tartalmazó transzkripciós gyári komplexet tömegspektrometriával elemeztük.
Fordított átírás
Fordított átírási sémaEgyes vírusok (például az AIDS-et okozó HIV) képesek RNS-t DNS-vé átírni. A HIV-nek van egy DNS-be integrált RNS-genomja. Ennek eredményeként a vírus DNS-e kombinálható a gazdasejt genomjával. A DNS RNS-ből történő szintéziséért felelős fő enzimet reverseáznak nevezik. A reversetas egyik funkciója, hogy komplementer DNS-t (cDNS) hozzon létre a vírusgenomból. A kapcsolódó ribonukleáz H enzim hasítja az RNS-t, és a reverz enzim cDNS-t szintetizál a DNS kettős hélixből. A cDNS-t az integráz integrálja a gazdasejt genomjába. Az eredmény a vírusfehérjék szintézise a gazdasejt által, amelyek új vírusokat képeznek. HIV esetén a T-limfociták apoptózisa (sejthalála) is be van programozva. Más esetekben a sejt a vírusok terjesztője maradhat.
Egyes eukarióta sejtek tartalmazzák a telomeráz enzimet, amely szintén reverz transzkripciós aktivitást mutat. Segítségével a DNS-ben ismétlődő szekvenciák szintetizálódnak. A telomeráz gyakran aktiválódik a rákos sejtekben, hogy korlátlan ideig megkettőzze a genomot anélkül, hogy elveszítené a fehérjét kódoló DNS-szekvenciát.
Megjegyzések
Wikimédia Alapítvány. 2010.
Nézze meg, mi az „átírás (biológia)” más szótárakban:
- (a latin transcriptio szóból, szó szerint átírás), RNS-molekulák bioszintézise, ill. DNS-szelvények; a genetikai megvalósítás első szakasza. információ az élő sejtekben. A DNS-függő RNS-polimeráz enzim hajtja végre, a legtöbb tanulmányozott paradicsomba... ... Biológiai enciklopédikus szótár
biológia- BIOLÓGIA (a görög bio élet és logosz szóból, doktrínából) az életről szóló tudományok összessége, formáinak, tulajdonságainak, kapcsolatainak és kapcsolatainak a Földön való megnyilvánulásainak sokféleségében. A kifejezést először egyidejűleg és egymástól függetlenül javasolták 1802-ben... ... Ismeretelméleti és Tudományfilozófiai Enciklopédia
Az élettudomány, amely magában foglalja az élőlények természetére, szerkezetére, működésére és viselkedésére vonatkozó összes tudást. A biológia nemcsak a különféle organizmusok formáinak sokféleségével foglalkozik, hanem azok evolúciójával, fejlődésével és azokkal a kapcsolatokkal is, amelyek... ... Collier enciklopédiája
BIOLÓGIA- az életről szóló tudományok összessége, formáinak, tulajdonságainak, kapcsolatainak és kapcsolatainak sokféleségében a Földön. A kifejezést először egyidejűleg és egymástól függetlenül javasolta 1802-ben a kiváló francia tudós, J.B. Lamarck és a német...... Tudományfilozófia: Alapfogalmak szójegyzéke
I Átírás (a latin transcriptio rewriting szóból) szavak, szövegek írásbeli sokszorosítása, kiejtésük figyelembevételével meghatározott grafikai rendszer segítségével. T. lehet tudományos és gyakorlati. A tudományos T. nyelvészeti...
- (a latin transcriptio szóból, betűk átírása), RNS bioszintézis DNS-mátrixon; a genetikai megvalósítás első szakasza. információ, a vágás során a DNS nukleotid szekvenciája RNS nukleotid szekvencia formájában olvasható ki (lásd Genetikai kód) ... Kémiai enciklopédia
Elő-mRNS szárhurokkal. A bázisokban lévő nitrogénatomokat kékkel, az oxigénatomokat a molekula foszfátvázában pirossal jelöljük A ribonukleinsavak (RNS) nukleinsavak, ortofoszforsav-maradékot tartalmazó nukleotidok polimerjei... Wikipédia
Olyan tudomány, amelynek célja az életjelenségek természetének megértése a biológiai objektumok és rendszerek molekuláris szintet megközelítő, esetenként ezt a határt elérő szinten történő tanulmányozásával. A végső cél...... Nagy Szovjet Enciklopédia
A reverz transzkripció egyszálú RNS-templátból kettős szálú DNS előállításának folyamata. Ezt a folyamatot fordított transzkripciónak nevezik, mivel a genetikai információ átvitele „fordítva” történik, viszonylag ... ... Wikipédia
A "Vírus" kérés ide kerül átirányításra. Lát más jelentések is. ? Vírusok Rotavírus Tudományos besorolás Overkingdom ... Wikipédia
A genetikai kód megfejtése után felmerült a kérdés: hogyan kerül át az információ a DNS-ből a fehérjébe? Biokémiai vizsgálatok kimutatták, hogy a sejt DNS-ének nagy része a sejtmagban lokalizálódik, míg a fehérjeszintézis a citoplazmában történik. A DNS-nek és a fehérjeszintézisnek ez a területi szétválasztása közvetítő kereséséhez vezetett. Mivel a fehérjeszintézis riboszómák részvételével zajlott, az RNS-t közvetítő szerepként javasolták. Készült egy diagram, amely szemlélteti a genetikai információ áramlásának irányát egy sejtben:
DNS → RNS → fehérje
Ezt nevezik a molekuláris biológia központi dogmájának. F. Crick feltételezte, hogy a makromolekulák e séma szerinti szintézise a mátrixelv szerint történik. Sok évbe telt, mire bebizonyították ennek a posztulátumnak a helyességét.
Először azt feltételezték, hogy a riboszómális RNS ("egy gén - egy riboszóma - egy fehérje") közvetítő szerepet játszik. Hamarosan azonban világossá vált, hogy ez a feltételezés tarthatatlan. Kimutatták, hogy a fehérjeszintézis során a riboszómák száma nem változik, i.e. új RNS nem szintetizálódik, és ezért nem érkezik új információ. Hamarosan az instabil RNS egy részét fedezték fel a riboszómák összetételében, amelyek molekuláit Mg-kationok segítségével lazán tartják a riboszómán. Molekuláris hibridizáció segítségével kimutatták, hogy ennek az RNS-nek a molekulái a DNS bizonyos szakaszainak másolatai. Megkapta a nevet mátrix, vagy hírvivő RNS. Korábban hírvivő RNS-nek és hírvivő RNS-nek is nevezték. Ezeknek a molekuláknak a komplementaritása a DNS bizonyos szakaszaival azt jelzi, hogy a DNS-en lévő templáttípus szerint szintetizálták őket.
Fokozatosan tisztázódott az információátvitel teljes útja a DNS-ből a fehérjébe. Két szakaszból áll: átiratokÉs adások. A transzkripció szakaszában a genetikai információ beolvasásra kerül és a DNS-ből az mRNS-be kerül. A transzkripciós folyamat három szakaszban zajlik: megindítás, inicializálás, megnyúlásÉs megszüntetése. Az információ csak egy DNS láncból (+ lánc) kerül kiolvasásra, mivel a genetikai kód tulajdonságai alapján a komplementer DNS szakaszok nem kódolhatják ugyanazon fehérje szerkezetét a kód komplementer degenerációjának hiánya miatt. A transzkripciót az RNS-polimeráz enzim végzi, amely négy alegységből (ααββ") áll, és nem rendelkezik specifikussággal a DNS forrását illetően. A transzkripció kezdeti szakaszában - iniciáció - egy ötödik alegység, az ún. s-faktor , kapcsolódik az enzimhez, amely felismeri a DNS egy meghatározott szakaszát, promótert A promóterek nem íródnak át.Az s-faktor felismeri őket egy specifikus nukleotid szekvencia jelenléte bennük.A bakteriális promóterekben Pribnov-nak nevezik. blokk és TATAAT formájú (enyhe eltérésekkel) A promoterhez az RNS polimeráz enzim kapcsolódik Az mRNS lánc növekedése egy irányban történik, a transzkripció sebessége ≈ 45-50 nukleotid/másodperc. csak egy rövid, 8 nukleotidból álló lánc szintetizálódik, ami után az s-faktor elválik az RNS polimeráztól és megkezdődik az elongációs szakasz Az mRNS lánc meghosszabbítását egy tetramer fehérje végzi A szekció, amelyből információkat olvasunk ki transzkripciónak nevezik.Terminátorral végződik - egy specifikus nukleotidszekvenciával, amely a stop jel szerepét tölti be. A terminátorhoz érve az RNS polimeráz enzim működése leáll, és fehérjeterminációs faktorok segítségével leválik a mátrixból.

A baktériumsejtekben a keletkező mRNS molekulák azonnal templátként szolgálhatnak a fehérjeszintézishez, azaz. adás. Ribszómákhoz kapcsolódnak, amelyekhez a transzport RNS (tRNS) molekulák egyidejűleg aminosavakat szállítanak. A transzfer RNS-láncok körülbelül 70 nukleotidból állnak. Az egyszálú tRNS molekula komplementer párosítási helyekkel rendelkezik, amelyek aktív centrumokat tartalmaznak: a tRNS felismerésének helye a tRNS-szintetáz enzim által, amely a megfelelő aktivált aminosavat a tRNS-hez kapcsolja; akceptor - az a hely, amelyhez az aminosav kapcsolódik, és az antikodon hurok.

Antikodon az mRNS-molekula megfelelő kodonjával komplementer triplett. A kodon-antikodon kölcsönhatás a komplementer párosítás típusát követi, melynek során egy aminosavat adnak a növekvő fehérjelánchoz. A különböző mRNS-ek startkodonja az AUG kodon, amely a metionin aminosavnak felel meg. Ezért az aktivált metionin aminosavhoz kapcsolódó UAC antikodont tartalmazó tRNS az első, amely megközelíti a mátrixot. Az aminosavakat aktiváló és a tRNS-hez kötődő enzimeket aminoacil-tRNS szintetázoknak nevezzük. A fehérje bioszintézis minden szakaszát (iniciáció, elongáció, termináció) a fehérje transzlációs faktorok szolgálják. A prokarióták mindegyik szakaszában három van belőlük. Az mRNS-templát végén nonszensz kodonok vannak, amelyeket nem olvasnak be, és a transzláció végét jelzik.
Számos organizmus genomjában, a baktériumoktól az emberekig, olyan géneket és megfelelő tRNS-eket fedeztek fel, amelyek nem szabványos kodonolvasást végeznek. Ezt a jelenséget az ún sugárzott kétértelműség.

Lehetővé teszi, hogy elkerülje az mRNS-molekulák szerkezetében a transzkripció során fellépő hibák negatív következményeit. Így, amikor nonszensz kodonok jelennek meg az mRNS-molekulában, amelyek képesek idő előtt leállítani a transzkripciós folyamatot, akkor aktiválódik az elnyomási mechanizmus. Abból áll, hogy a tRNS egy szokatlan formája jelenik meg a sejtben a nonszensz kodonnal komplementer antikodonnal, aminek normális esetben nem kellene léteznie. Megjelenése a nonszensz kodonhoz hasonló összetételű tRNS antikodonban bázist helyettesítő gén hatásának eredménye. A helyettesítés eredményeként a nonszensz kodont szabályos szignifikáns kodonként olvassuk be. Az ilyen mutációkat szupresszor mutációknak nevezzük, mert elnyomják az eredeti mutációt, amely a nonszensz kodonhoz vezetett.
A szén formájú élet a fehérjemolekulák jelenléte miatt létezik. A fehérje bioszintézise pedig a sejtben az egyetlen lehetőség a génexpresszióra. Ennek a folyamatnak a megvalósításához azonban számos olyan folyamatot kell elindítani, amelyek a genetikai információ „kicsomagolásához”, a kívánt gén kereséséhez, kiolvasásához és reprodukálásához kapcsolódnak. A „transzkripció” kifejezés a biológiában kifejezetten arra a folyamatra utal, amely során az információ egy génből a hírvivő RNS-be kerül. Ez a bioszintézis kezdete, vagyis a genetikai információ közvetlen megvalósítása.
Genetikai információ tárolása
Az élő szervezetek sejtjeiben a genetikai információ a sejtmagban, a mitokondriumokban, a kloroplasztiszokban és a plazmidokban lokalizálódik. A mitokondriumok és a kloroplasztiszok kis mennyiségben tartalmaznak állati és növényi DNS-t, míg a bakteriális plazmidok a környezeti feltételekhez való gyors alkalmazkodásért felelős gének tárolóhelyei.
A vírustestekben az öröklődő információkat is RNS vagy DNS polimerek formájában tárolják. De megvalósításának folyamata összefügg az átírás szükségességével is. A biológiában ennek a folyamatnak rendkívüli jelentősége van, hiszen ez vezet az öröklődő információk megvalósításához, beindítva a fehérje bioszintézist.

Az állati sejtekben az örökletes információt a DNS polimerje képviseli, amely tömören van csomagolva a sejtmag belsejében. Ezért a fehérjeszintézis vagy bármely gén leolvasása előtt bizonyos szakaszoknak át kell menniük: a kondenzált kromatin feltekercselése és a kívánt gén „felszabadulása”, enzimmolekulák általi felismerése, transzkripció.
A biológiában és a biológiai kémiában ezeket a szakaszokat már tanulmányozták. Egy fehérje szintéziséhez vezetnek, amelynek elsődleges szerkezetét egyetlen gén kódolja.
Transzkripciós mintázat eukarióta sejtekben
Bár a biológiában a transzkripciót nem vizsgálták kellőképpen, szekvenciáját hagyományosan diagram formájában mutatják be. Kezdésből, megnyúlásból és befejezésből áll. Ez azt jelenti, hogy az egész folyamat három komponensű jelenségre oszlik.

Az iniciáció olyan biológiai és biokémiai folyamatok összessége, amelyek a transzkripció kezdetéhez vezetnek. Az elongáció lényege a molekulalánc folyamatos növekedése. A termináció olyan folyamatok összessége, amelyek az RNS-szintézis leállásához vezetnek. Egyébként a fehérjebioszintézissel összefüggésben a biológiában a transzkripció folyamatát általában a hírvivő RNS szintézisével azonosítják. Ennek alapján később egy polipeptidláncot szintetizálnak.

Megindítás, inicializálás
Az iniciáció a legkevésbé ismert transzkripciós mechanizmus a biológiában. Biokémiai szempontból mi ez, nem ismert. Vagyis a transzkripció kiváltásáért felelős specifikus enzimeket egyáltalán nem ismerik fel. Szintén ismeretlenek az intracelluláris szignálok és átvitelük módjai, amelyek egy új fehérje szintézisének szükségességét jelzik. Ez a citológia és a biokémia alapvető feladata.
Megnyúlás
Az iniciáció és a megnyúlás folyamatát még nem lehet időben szétválasztani, mivel nem lehet olyan laboratóriumi vizsgálatokat végezni, amelyek célja specifikus enzimek és trigger tényezők jelenlétének megerősítése. Ezért ez a határ nagyon feltételes. Az elongációs folyamat lényege a DNS templát szakasz alapján szintetizált növekedési lánc meghosszabbításában rejlik.
Úgy gondolják, hogy az elongáció az RNS polimeráz első transzlokációja és az első kadon RNS kiindulási helyéhez való kapcsolódása után kezdődik. Az elongáció során a kadonokat a 3"-5" szál irányában olvassuk le egy két szálra osztott, despiralizált DNS-szakaszon. Ezzel egyidejűleg a növekvő RNS-lánc új, a templát DNS-régióval komplementer nukleotidokkal egészül ki. Ebben az esetben a DNS 12 nukleotid szélességre, azaz 4 kadonra „bővül”.

Az RNS-polimeráz enzim a növekvő lánc mentén mozog, és „mögötte” a DNS fordítottan „keresztkötésbe” kerül egy kettős szálú szerkezetbe, a nukleotidok közötti hidrogénkötések helyreállításával. Ez részben választ ad arra a kérdésre, hogy milyen folyamatot nevezünk transzkripciónak a biológiában. Az elongáció a transzkripció fő fázisa, mivel ennek során a gén és a fehérjeszintézis között az úgynevezett közvetítő áll össze.
Felmondás
Az eukarióta sejtekben végbemenő transzkripciós folyamat kevéssé ismert. Eddig a tudósok a lényegét arra csökkentették, hogy leállítsák a DNS-olvasást az 5"-es végén, és egy csoport adeninbázist kapcsoljanak az RNS 3"-os végéhez. Ez utóbbi eljárás lehetővé teszi a keletkező RNS kémiai szerkezetének stabilizálását. A bakteriális sejtekben kétféle termináció létezik. Ez egy Rho-függő és Rho-független folyamat.

Az első a Rho fehérje jelenlétében következik be, és a DNS templát régiója és a szintetizált RNS közötti hidrogénkötések egyszerű megszakadására redukálódik. A második, Rho-független, a szárhurok megjelenése után következik be, ha van mögötte egy sor uracilbázis. Ez a kombináció az RNS leválását okozza a DNS-templátról. Nyilvánvaló, hogy a transzkripció terminációja enzimatikus folyamat, de specifikus biokatalizátort még nem találtak rá.
Vírus transzkripció
A vírustestek nem rendelkeznek saját fehérje-bioszintézis-rendszerrel, ezért nem tudnak szaporodni a sejtek kihasználása nélkül. De a vírusoknak megvan a saját genetikai anyaguk, amelyet meg kell valósítani, és integrálni kell a fertőzött sejtek génjeibe. Ehhez számos enzimük van (vagy kihasználják a sejtenzimrendszereket), amelyek átírják nukleinsavukat. Vagyis ez az enzim a vírus genetikai információi alapján szintetizálja a hírvivő RNS analógját. De ez egyáltalán nem RNS, hanem egy DNS polimer, amely komplementer például az emberi génekkel.
Ez teljesen sérti a biológia transzkripciójának hagyományos elveit, amint az a HIV-vírus példáján is látható. Reverz enzimje képes a humán nukleinsavval komplementer DNS-t szintetizálni vírus RNS-ből. Az RNS-ből komplementer DNS szintetizálásának folyamatát reverz transzkripciónak nevezik. A biológiában ez a definíciója annak a folyamatnak, amely a vírus örökletes információinak az emberi genomba való beépüléséért felelős.
Fürdőkádak helyreállítása Kolpinóban vk.com/restavraciya_vann_kolpino.
Átírás. Kezdete - a transzkripció kezdete, vége - a transzkripció vége, DNS - DNS.
A transzkripció az RNS-szintézis folyamata, amely a DNS-t templátként használja, és minden élő sejtben előfordul. Más szavakkal, ez a genetikai információ átvitele a DNS-ből az RNS-be.
A transzkripciót a DNS-függő RNS-polimeráz enzim katalizálja. Az RNS szintézis folyamata az 5"-től a 3"-ig terjedő irányban halad, vagyis a DNS-templát szál mentén az RNS-polimeráz 3"->5" irányba mozog.
A transzkripció az iniciáció, az elongáció és a termináció szakaszaiból áll.
Az átírás megkezdése
A transzkripció iniciációja egy összetett folyamat, amely az átírt szekvencia közelében található DNS-szekvenciától és a különböző fehérjefaktorok jelenlététől vagy hiányától függ.
Átírási megnyúlás
Az a pillanat, amikor az RNS-polimeráz a transzkripció iniciációjából az elongációba megy át, nincs pontosan meghatározva. Három jelentősebb biokémiai esemény jellemzi ezt az átmenetet az Escherichia coli RNS polimeráz esetében: a szigma faktor felszabadulása, az enzimmolekula első transzlokációja a templát mentén, valamint a transzkripciós komplex erős stabilizálódása, amely az RNS mellett polimeráz, magában foglalja a növekvő RNS-láncot és az átírt DNS-t. Ugyanezek a jelenségek jellemzőek az eukarióta RNS polimerázokra is. Az iniciációból az elongációba való átmenetet az enzim, a promoter, a transzkripciós iniciációs faktorok közötti kötések felszakadása, egyes esetekben az RNS polimeráz elongációs kompetencia állapotába való átmenete kíséri. Az elongációs fázis azután ér véget, hogy a növekvő transzkriptum felszabadul, és az enzim disszociál a templátról.
Az elongációs szakasz során körülbelül 18 nukleotidpár csavarodik fel a DNS-ben. A DNS-templátszál körülbelül 12 nukleotidja hibrid hélixet alkot az RNS-szál növekvő végével. Ahogy az RNS-polimeráz áthalad a templáton, előtte megtörténik a DNS kettős hélix feltekerése, mögötte pedig a DNS kettős hélix helyreállítása. Ezzel egyidejűleg a növekvő RNS-lánc következő láncszeme felszabadul a templáttal és RNS-polimerázzal alkotott komplexből. Ezeket a mozgásokat az RNS-polimeráz és a DNS relatív forgásának kell kísérnie. Nehéz elképzelni, hogy ez hogyan történhetne meg egy sejtben, különösen a kromatin transzkripció során. Ezért lehetséges, hogy az ilyen forgás megakadályozására a DNS mentén mozgó RNS-polimerázt topoizomerázok kísérik.
A nyúlás alapvető nyúlási tényezők segítségével történik, amelyek szükségesek ahhoz, hogy a folyamat ne álljon le idő előtt.
A közelmúltban bizonyítékok jelentek meg, amelyek azt mutatják, hogy a szabályozó tényezők is szabályozhatják a megnyúlást. Az elongációs folyamat során az RNS-polimeráz a gén bizonyos részein megáll. Ez különösen jól látható kis szubsztrátumkoncentráció esetén. A mátrix egyes területein hosszú késések tapasztalhatók az RNS polimeráz előrehaladása, az ún. még optimális szubsztrátkoncentráció esetén is szünetek figyelhetők meg. Ezeknek a szüneteknek az időtartama nyúlási tényezőkkel szabályozható.
| Triptofán operon |
A transzkripció a szintézis folyamatamolekulákRNS bekapcsolvaterületDNS, mátrixként használják. Az átírás jelentése az genetikai információ átvitele a DNS-ből az RNS-be.
Egy DNS-molekula két komplementer szálból áll, míg az RNS csak egyből. A transzkripció során csak az egyik DNS-szál szolgál templátként az RNS-szintézishez. Őt hívják szemantikai lánc. A kivétel a mitokondriális DNS, amelyben mindkét szál értelmes és különböző géneket tartalmaz. Amellett, hogy a nukleáris DNS kivételét képezik, egyes gének egy értelmetlen szálon is lokalizálhatók.
A transzkripció során az RNS molekula az 5"-től a 3"-ig terjedő irányban szintetizálódik (ami természetes az összes nukleinsav szintézisénél), míg a DNS-lánc mentén a szintézis ellenkező irányban megy végbe: 3"→5 ".
Az eukariótákban minden gén külön-külön íródik át. A kivétel ismét a mitokondriális DNS, amely egy közös többgénes transzkriptummá íródik át, amelyet aztán levágnak. Mivel a prokariótákban a gének csoportokat alkotnak, egy operont alkotva, az ilyen gének együtt íródnak át. Akárhogyan is transzkripton DNS szakasznak nevezzük, amely egy promoterből, egy átírt régióból és egy terminátorból áll.
Az átírásnak 3 szakasza van: iniciáció, megnyúlás, befejezés.
Megindítás, inicializálás A transzkripció lehetővé teszi az RNS-molekula szintézisének megkezdését. Az iniciálás során egy enzimkomplexet kapcsolnak a promoterhez. A fő az RNS-polimeráz (jelen esetben DNS-függő), amely viszont több alegységfehérjéből áll, és a folyamat katalizátoraként játszik szerepet. Az eukariótákban a transzkripció megindulását a DNS speciális szakaszai befolyásolják: fokozók (erősítik) és hangtompítók (elnyomják), amelyek általában magától a géntől bizonyos távolságra helyezkednek el. Különféle fehérjefaktorok befolyásolják a transzkripció beindulásának lehetőségét.
A prokariótáknak csak egyféle RNS-polimerázuk van, míg az eukariótáknak három. Az RNS polimeráz 1 háromféle riboszomális RNS szintetizálására szolgál (összesen 4 típusú rRNS létezik). Az RNS-polimeráz 2-t pre-mRNS (prekurzor hírvivő RNS) szintetizálására használják. Az RNS polimeráz-3 szintetizálja a riboszómális RNS egyik típusát, a transzport és a kis nukleáris RNS-t.
Az RNS-polimeráz képes felismerni specifikus nukleotidszekvenciákat, és azokhoz kapcsolódni. Ezek a sorozatok rövidek és univerzálisak minden élőlény számára.
Miután az RNS polimeráz a promoterhez kötődik, a DNS kettős hélix egy része feltekerődik, és a szakasz szálai közötti nukleotid kötések megszakadnak. Körülbelül 18 pár nukleotid bomlik fel.
A színpadon megnyúlás a szekvenciális addíció a szabad nukleotidok komplementaritása elve szerint történik a felszabadult DNS-szakaszhoz. Az RNS polimeráz a nukleotidokat egy poliribonukleotid láncba egyesíti.
Az RNS szintézis során körülbelül 12 nukleotidja komplementer és átmenetileg kapcsolódik DNS nukleotidokhoz. Ha az RNS-polimeráz előtte mozog, a DNS-láncok szétválnak, mögötte pedig enzimek segítségével „összevarrják”. Az RNS-lánc fokozatosan növekszik, és kimozdul az RNS-polimeráz komplexből.
Vannak olyan elongációs tényezők, amelyek megakadályozzák a transzkripció idő előtti leállását.
Felmondás A transzkripciós folyamat a terminátor régióban megy végbe, amelyet az RNS polimeráz speciális fehérjeterminációs faktoroknak köszönhetően ismer fel.
A szintetizált RNS-molekula 3"-os végéhez sok adenin nukleotid (poli-A) kapcsolódik, hogy megakadályozza annak enzimatikus lebomlását. Még korábban, amikor az 5"-es végét szintetizálták, az ún. sapka.
A legtöbb esetben a transzkripció nem termel kész RNS-t. A nyers RNS-nek még át kell mennie egy folyamaton feldolgozás, amelynél módosulása megváltozik és funkcionálisan aktívvá válik. Az eukariótákban minden RNS-típus saját módosulásokon megy keresztül. A poli-A és sapka kialakulását gyakran feldolgozásnak is nevezik.
Azt is ajánljuk
 Numerikus szimbolizmus János Jelenések könyvében
Numerikus szimbolizmus János Jelenések könyvében
 „Konfliktológia” képzés: arab-izraeli konfliktus
„Konfliktológia” képzés: arab-izraeli konfliktus
 Csoda a kígyóval, ami György szent nagy vértanúval történt
Csoda a kígyóval, ami György szent nagy vértanúval történt
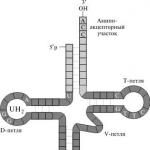 Az RNS örökletes információkat kap
Az RNS örökletes információkat kap
 Scaevola, Gaius Mucius etruszkokkal vívott háború hősének tiszteletbeli neve
Scaevola, Gaius Mucius etruszkokkal vívott háború hősének tiszteletbeli neve
 Hol van az állítmány? Mi az a „predikátum”? A predikátumok típusai. Igaz-e, hogy az egyszerű verbális állítmány olyan állítmány, amelynek lexikai és nyelvtani jelentését egyetlen ige fejezi ki?
Hol van az állítmány? Mi az a „predikátum”? A predikátumok típusai. Igaz-e, hogy az egyszerű verbális állítmány olyan állítmány, amelynek lexikai és nyelvtani jelentését egyetlen ige fejezi ki?