Vad är transkriptionsprocessen? RNA tar emot ärftlig information
Initiering av transkription
Transkriptionsförlängning
Det ögonblick då RNA-polymeras övergår från transkriptionsinitiering till förlängning är inte exakt bestämt. Tre stora biokemiska händelser kännetecknar denna övergång i fallet med Escherichia coli RNA-polymeras: frisättningen av sigmafaktorn, den första translokationen av enzymmolekylen längs mallen och den starka stabiliseringen av transkriptionskomplexet, som förutom RNA:t polymeras, inkluderar den växande RNA-kedjan och det transkriberade DNA:t. Samma fenomen är också karakteristiska för eukaryota RNA-polymeraser. Övergången från initiering till förlängning åtföljs av brott av bindningar mellan enzymet, promotorn, traoch i vissa fall av övergången av RNA-polymeras till ett tillstånd av förlängningskompetens (till exempel fosforylering av CTD-domänen i RNA-polymeras II). Förlängningsfasen slutar efter att det växande transkriptet har frigjorts och enzymet dissocierar från mallen (terminering).
Förlängning utförs med hjälp av grundläggande förlängningsfaktorer, som är nödvändiga för att processen inte ska stoppa i förtid.
Nyligen har bevis dykt upp som visar att regulatoriska faktorer också kan reglera förlängning. Under förlängningsprocessen pausar RNA-polymeras vid vissa delar av genen. Detta ses särskilt tydligt vid låga koncentrationer av substrat. I vissa områden av matrisen finns det långa förseningar i utvecklingen av RNA-polymeras, den så kallade. pauser observeras även vid optimala substratkoncentrationer. Varaktigheten av dessa pauser kan styras av förlängningsfaktorer.
Uppsägning
Bakterier har tvåsmer:
- en rho-beroende mekanism där Rho (rho)-proteinet destabiliserar vätebindningarna mellan DNA-mallen och mRNA:t och frisätter RNA-molekylen.
- rho-oberoende, där transkriptionen stoppas när den nysyntetiserade RNA-molekylen bildar en stam-loop, följt av flera uraciler (...UUUU), vilket leder till att RNA-molekylen lossnar från DNA-mallen.
Transkriptionsterminering i eukaryoter studeras mindre. Det slutar med att RNA skärs, varefter enzymet lägger till flera adeniner (...AAAA) till sin 3"-ände, vars antal bestämmer stabiliteten för ett givet transkript.
Transkriptionsfabriker
Det finns ett antal experimentella data som indikerar att transkription sker i de så kallade transkriptionsfabrikerna: enorma, enligt vissa uppskattningar, upp till 10 Da-komplex som innehåller cirka 8 RNA-polymeraser II och komponenter för efterföljande bearbetning och splitsning, såväl som korrigering av det nysyntetiserade transkriptet. I cellkärnan sker ett konstant utbyte mellan pooler av lösligt och aktiverat RNA-polymeras. Aktivt RNA-polymeras är involverat i ett sådant komplex, som i sin tur är en strukturell enhet som organiserar kromatinkomprimering. Nya data indikerar att transkriptionsfabriker existerar även i frånvaro av transkription, de är fixerade i cellen (det är ännu inte klart om de interagerar med cellens kärnmatris eller inte) och representerar en oberoende nukleär subkompartment. Transkriptionsfabrikskomplexet innehållande RNA-polymeras I, II eller III analyserades med masspektrometri.
Omvänd transkription
Schema för omvänd transkriptionVissa virus (som HIV, som orsakar AIDS) har förmågan att transkribera RNA till DNA. HIV har ett RNA-genom som är integrerat i DNA. Som ett resultat kan virusets DNA kombineras med värdcellens genom. Det huvudsakliga enzymet som är ansvarigt för att syntetisera DNA från RNA kallas reversease. En av funktionerna hos reversetas är att skapa komplementärt DNA (cDNA) från det virala genomet. Det associerade enzymet ribonukleas H klyver RNA och reversease syntetiserar cDNA från DNA-dubbelhelixen. cDNA:t integreras i värdcellens genom genom integras. Resultatet är syntesen av virala proteiner av värdcellen, som bildar nya virus. I fallet med HIV programmeras även apoptos (celldöd) av T-lymfocyter. I andra fall kan cellen förbli en spridare av virus.
Vissa eukaryota celler innehåller enzymet telomeras, som också uppvisar omvänd transkriptionsaktivitet. Med dess hjälp syntetiseras repeterande sekvenser i DNA. Telomeras aktiveras ofta i cancerceller för att på obestämd tid duplicera genomet utan att förlora den proteinkodande DNA-sekvensen.
Anteckningar
Wikimedia Foundation. 2010.
Se vad "Transcription (biologi)" är i andra ordböcker:
- (från latin transcriptio, lit. omskrivning), biosyntes av RNA-molekyler, resp. DNA-sektioner; det första steget av genetisk implementering. information i levande celler. Det utförs av enzymet DNA-beroende RNA-polymeras, till paradiset för de mest studerade... ... Biologisk encyklopedisk ordbok
biologi- BIOLOGI (från det grekiska ordet bioliv och logos, doktrin) helheten av vetenskaper om livet i alla mångfalden av manifestationer av dess former, egenskaper, samband och relationer på jorden. Termen föreslogs första gången samtidigt och oberoende 1802... ... Encyclopedia of Epistemology and Philosophy of Science
Livsvetenskapen som omfattar all kunskap om levande varelsers natur, struktur, funktion och beteende. Biologi handlar inte bara om den stora variationen av former av olika organismer, utan också om deras evolution, utveckling och de relationer som... ... Colliers uppslagsverk
BIOLOGI- en uppsättning vetenskaper om livet i alla mångfalden av manifestationer av dess former, egenskaper, samband och relationer på jorden. Termen föreslogs första gången samtidigt och oberoende av varandra 1802 av den framstående franske vetenskapsmannen J.B. Lamarck och tyska... ... Vetenskapsfilosofi: Ordlista över grundläggande termer
I Transcription (från latin transcriptio rewriting) skriftlig återgivning av ord och texter, med hänsyn till deras uttal med hjälp av ett visst grafiskt system. T. kan vara vetenskaplig och praktisk. Scientific T. används i språklig...
- (från latin transcriptio, bokstäver omskrivning), RNA-biosyntes på en DNA-matris; det första steget av genetisk implementering. information, under skärningen av nukleotidsekvensen av DNA läses i form av en nukleotidsekvens av RNA (se genetisk kod) ... Kemiskt uppslagsverk
Pre-mRNA med stamslinga. Kväveatomer i baserna är markerade med blått, syreatomer i fosfatryggraden i molekylen i rött Ribonukleinsyror (RNA) är nukleinsyror, polymerer av nukleotider som innehåller en ortofosforsyrarest ... Wikipedia
En vetenskap som syftar till att förstå livsfenomens natur genom att studera biologiska objekt och system på en nivå som närmar sig den molekylära nivån, och i vissa fall nå denna gräns. Det slutliga målet är... ... Stora sovjetiska encyklopedien
Omvänd transkription är processen för att producera dubbelsträngat DNA från en enkelsträngad RNA-mall. Denna process kallas omvänd transkription, eftersom överföringen av genetisk information sker "omvänt", relativt ... ... Wikipedia
Begäran "Virus" omdirigeras hit. Ser även andra betydelser. ? Virus Rotavirus Vetenskaplig klassificering Overkingdom ... Wikipedia
Efter att ha dechiffrerat den genetiska koden uppstod frågan: hur överförs information från DNA till protein? Biokemiska studier har fastställt att huvuddelen av DNA i en cell är lokaliserad i kärnan, medan proteinsyntes sker i cytoplasman. Denna territoriella separation av DNA och proteinsyntes ledde till sökandet efter en mellanhand. Eftersom proteinsyntes ägde rum med deltagande av ribosomer, föreslogs RNA för att spela rollen som en mellanhand. Ett diagram skapades som illustrerar riktningen för flödet av genetisk information i en cell:
DNA → RNA → protein
Det kallas molekylärbiologins centrala dogm. F. Crick postulerade att syntesen av makromolekyler enligt detta schema utförs enligt matrisprincipen. Det tog många år att bevisa riktigheten av detta postulat.
Först antog man att ribosomalt RNA ("en gen - en ribosom - ett protein") spelade rollen som en mellanhand. Det stod emellertid snart klart att detta antagande var ohållbart. Det har visat sig att under proteinsyntesen inte ändras antalet ribosomer, d.v.s. nytt RNA syntetiseras inte och därför tas ingen ny information emot. Snart upptäcktes en bråkdel av instabilt RNA i sammansättningen av ribosomer, vars molekyler hålls löst på ribosomen med hjälp av Mg-katjoner. Med hjälp av molekylär hybridisering visades det att molekylerna i detta RNA är kopior av vissa delar av DNA. Hon fick namnet matris, eller budbärar-RNA. Det kallades också tidigare budbärar-RNA och budbärar-RNA. Komplementariteten av dessa molekyler till vissa sektioner av DNA indikerade att de syntetiserades enligt en malltyp på DNA.
Efterhand klargjordes hela vägen för informationsöverföring från DNA till protein. Den består av två steg: transkriptioner Och sändningar. I transkriptionsstadiet läses genetisk information och överförs från DNA till mRNA. Transkriptionsprocessen sker i tre steg: initiering, förlängning Och uppsägning. Information läses endast från en DNA-kedja (+-kedja), eftersom, baserat på egenskaperna hos den genetiska koden, komplementära DNA-sektioner inte kan koda strukturen för samma protein på grund av avsaknaden av komplementär degeneration av koden. Transkription utförs av enzymet RNA-polymeras, som består av fyra subenheter (ααββ") och inte har specificitet vad gäller källan till DNA. Vid det inledande skedet av transkription - initiering - en femte subenhet, den så kallade s-faktorn , är fäst till enzymet, som känner igen en specifik sektion av DNA, promotor. Promotorer transkriberas inte. De känns igen av s-faktorn genom närvaron av en specifik nukleotidsekvens i dem. I bakteriella promotorer kallas det Pribnov block och har formen TATAAT (med små variationer) Enzymet RNA-polymeras är fäst till promotorn. Tillväxten av mRNA-kedjan sker i en riktning, transkriptionshastigheten är ≈ 45-50 nukleotider per sekund. Vid initieringsstadiet, endast en kort kedja på 8 nukleotider syntetiseras, varefter s-faktorn separeras från RNA-polymeras och förlängningsstadiet börjar. Förlängningen av mRNA-kedjan utförs av ett tetramerprotein. Avsnitt , från vilket information läses, är kallas transkripton.Det slutar med en terminator - en specifik nukleotidsekvens som spelar rollen som en stoppsignal. Efter att ha nått terminatorn slutar RNA-polymerasenzymet att fungera och separeras med hjälp av proteintermineringsfaktorer från matrisen.

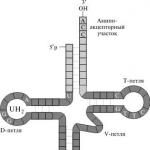
I bakterieceller kan de resulterande mRNA-molekylerna omedelbart fungera som mallar för proteinsyntes, d.v.s. utsända. De ansluter till ribosomer, till vilka transport-RNA (tRNA)-molekyler samtidigt levererar aminosyror. Transfer RNA-kedjor består av cirka 70 nukleotider. En enkelsträngad tRNA-molekyl har ställen för komplementär parning, som innehåller aktiva centra: ett ställe för igenkänning av tRNA av enzymet tRNA-syntetas, som fäster motsvarande aktiverade aminosyra till tRNA; acceptor - platsen till vilken aminosyran är fäst och antikodonloopen.

Antikodonär en triplett som är komplementär till motsvarande kodon i mRNA-molekylen. Kodon-antikodon-interaktionen följer typen av komplementär parning, under vilken en aminosyra läggs till den växande proteinkedjan. Startkodonet i olika mRNA är AUG-kodonet, motsvarande aminosyran metionin. Därför är tRNA:t med UAC-antikodonet, kopplat till den aktiverade aminosyran metionin, det första som närmar sig matrisen. Enzymer som aktiverar aminosyror och kopplar dem till tRNA kallas aminoacyl-tRNA-syntetaser. Alla stadier av proteinbiosyntes (initiering, förlängning, avslutning) betjänas av proteintranslationsfaktorer. Prokaryoter har tre av dem för varje steg. I slutet av mRNA-mallen finns det nonsenskodon som inte läses och markerar slutet på translationen.
I arvsmassan hos många organismer, från bakterier till människor, har gener och motsvarande tRNA:n som utför icke-standardiserad avläsning av kodon upptäckts. Detta fenomen kallas tvetydighet i sändningen.

Det låter dig undvika de negativa konsekvenserna av fel som uppstår i strukturen av mRNA-molekyler under transkription. Sålunda, när nonsenskodon dyker upp inuti mRNA-molekylen, som kan stoppa transkriptionsprocessen i förtid, aktiveras undertryckningsmekanismen. Den består i att en ovanlig form av tRNA dyker upp i cellen med ett antikodon som är komplementärt till nonsenskodonet, vilket normalt inte borde finnas. Dess utseende är resultatet av verkan av en gen som ersätter en bas i tRNA-antikodonet, som till sin sammansättning liknar nonsenskodonet. Som ett resultat av denna ersättning läses nonsenskodonet som ett vanligt signifikant kodon. Sådana mutationer kallas suppressormutationer, eftersom de undertrycker den ursprungliga mutationen som ledde till nonsenskodonet.
Liv i kolform existerar på grund av närvaron av proteinmolekyler. Och proteinbiosyntes i cellen är den enda möjligheten för genuttryck. Men för att implementera denna process är det nödvändigt att starta ett antal processer associerade med "uppackning" av genetisk information, sökning efter den önskade genen, läsning av den och reproducering av den. Termen "transkription" i biologi hänvisar specifikt till processen att överföra information från en gen till budbärar-RNA. Detta är början på biosyntes, det vill säga den direkta implementeringen av genetisk information.
Lagring av genetisk information
I cellerna hos levande organismer är genetisk information lokaliserad i kärnan, mitokondrier, kloroplaster och plasmider. Mitokondrier och kloroplaster innehåller en liten mängd djur- och växt-DNA, medan bakterieplasmider är lagringsplatsen för gener som är ansvariga för snabb anpassning till miljöförhållanden.
I virala kroppar lagras även ärftlig information i form av RNA eller DNA-polymerer. Men processen för dess implementering är också förknippad med behovet av transkription. Inom biologi är denna process av exceptionell betydelse, eftersom det är den som leder till implementeringen av ärftlig information, vilket utlöser proteinbiosyntes.

I djurceller representeras ärftlig information av en polymer av DNA, som är kompakt förpackad inuti kärnan. Därför, innan proteinsyntes eller läsning av någon gen, måste vissa stadier passera: avveckling av kondenserat kromatin och "frisättning" av den önskade genen, dess igenkänning av enzymmolekyler, transkription.
Inom biologi och biologisk kemi har dessa stadier redan studerats. De leder till syntesen av ett protein, vars primära struktur kodades i en enda gen.
Transkriptionsmönster i eukaryota celler
Även om transkription i biologi inte har studerats tillräckligt, presenteras dess sekvens traditionellt i form av ett diagram. Den består av initiering, förlängning och avslutning. Detta innebär att hela processen är uppdelad i tre komponentfenomen.

Initiering är en uppsättning biologiska och biokemiska processer som leder till början av transkription. Kärnan i förlängningen är den fortsatta tillväxten av molekylkedjan. Terminering är en uppsättning processer som leder till att RNA-syntesen upphör. Förresten, i samband med proteinbiosyntes identifieras transkriptionsprocessen i biologi vanligtvis med syntesen av budbärar-RNA. Baserat på det kommer en polypeptidkedja senare att syntetiseras.

Initiering
Initiering är den minst förstådda transkriptionsmekanismen inom biologi. Vad det är ur biokemisk synvinkel är okänt. Det vill säga att de specifika enzymer som är ansvariga för att utlösa transkription inte känns igen alls. Okänd är också de intracellulära signalerna och metoderna för deras överföring, vilket indikerar behovet av syntes av ett nytt protein. Detta är en grundläggande uppgift för cytologi och biokemi.
Förlängning
Det är ännu inte möjligt att separera processen för initiering och förlängning i tid på grund av omöjligheten att genomföra laboratoriestudier utformade för att bekräfta närvaron av specifika enzymer och triggerfaktorer. Därför är denna gräns mycket villkorad. Kärnan i förlängningsprocessen handlar om att förlänga den växande kedjan, syntetiserad på basis av DNA-mallsektionen.
Man tror att förlängning börjar efter den första translokationen av RNA-polymeras och början av fästningen av det första kadonet till startstället för RNA. Under förlängning avläses kadon i riktningen för 3"-5"-strängen på en despiraliserad DNA-sektion uppdelad i två strängar. Samtidigt tillsätts den växande RNA-kedjan med nya nukleotider som är komplementära till mall-DNA-regionen. I det här fallet "expanderas" DNA:t till en bredd av 12 nukleotider, det vill säga 4 kadon.

Enzymet RNA-polymeras rör sig längs den växande kedjan, och "bakom" den är DNA:t omvänt "tvärbunden" till en dubbelsträngad struktur med återställande av vätebindningar mellan nukleotider. Detta svarar delvis på frågan om vilken process som kallas transkription inom biologi. Det är förlängning som är transkriptionens huvudfas, eftersom den så kallade intermediären mellan genen och proteinsyntesen under dess förlopp samlas.
Uppsägning
Processen för transkriptionsterminering i eukaryota celler är dåligt förstådd. Hittills har forskare reducerat dess väsen till att stoppa DNA-läsning vid 5"-änden och fästa en grupp adeninbaser till 3-tumsänden av RNA. Den senare processen tillåter att den kemiska strukturen hos det resulterande RNA:t stabiliseras. Det finns två typer av terminering i bakterieceller. Det är en Rho-beroende och Rho-oberoende process.

Den första inträffar i närvaro av Rho-proteinet och reduceras till en enkel brytning av vätebindningar mellan mallregionen av DNA och det syntetiserade RNA:t. Den andra, Rho-oberoende, inträffar efter uppkomsten av stamslingan om det finns en uppsättning uracilbaser bakom den. Denna kombination gör att RNA:t lossnar från DNA-mallen. Det är uppenbart att transkriptionsterminering är en enzymatisk process, men specifika biokatalysatorer för det har ännu inte hittats.
Viral transkription
Virala kroppar har inte sitt eget proteinbiosyntessystem och kan därför inte föröka sig utan att utnyttja celler. Men virus har sitt eget genetiska material, som måste realiseras och även integreras i infekterade cellers gener. För att göra detta har de ett antal enzymer (eller utnyttjar cellenzymsystem) som transkriberar deras nukleinsyra. Det vill säga att detta enzym, baserat på virusets genetiska information, syntetiserar en analog av budbärar-RNA. Men det är inte alls RNA, utan en DNA-polymer, komplementär till till exempel mänskliga gener.
Detta bryter helt mot de traditionella principerna för transkription inom biologin, vilket kan ses i exemplet med HIV-viruset. Dess omvända enzymenzym kan syntetisera DNA som är komplementärt till human nukleinsyra från viralt RNA. Processen att syntetisera komplementärt DNA från RNA kallas omvänd transkription. Detta är definitionen i biologi av processen som är ansvarig för integreringen av virusets ärftliga information i det mänskliga genomet.
Restaurering av badkar i Kolpino vk.com/restavraciya_vann_kolpino.
Transkription. Börja - början av transkription, Slut - slutet av transkription, DNA - DNA.
Transkription är processen för RNA-syntes med DNA som mall och sker i alla levande celler. Det är med andra ord överföringen av genetisk information från DNA till RNA.
Transkription katalyseras av enzymet DNA-beroende RNA-polymeras. Processen för RNA-syntes fortsätter i riktningen från 5" till 3"-änden, det vill säga längs DNA-mallsträngen rör sig RNA-polymeras i riktningen 3"->5"
Transkription består av stadierna initiering, förlängning och avslutning.
Initiering av transkription
Transkriptionsinitiering är en komplex process som beror på DNA-sekvensen i närheten av den transkriberade sekvensen och på närvaron eller frånvaron av olika proteinfaktorer.
Transkriptionsförlängning
Det ögonblick då RNA-polymeras övergår från transkriptionsinitiering till förlängning är inte exakt bestämt. Tre stora biokemiska händelser kännetecknar denna övergång i fallet med Escherichia coli RNA-polymeras: frisättningen av sigmafaktorn, den första translokationen av enzymmolekylen längs mallen och den starka stabiliseringen av transkriptionskomplexet, som förutom RNA:t polymeras, inkluderar den växande RNA-kedjan och det transkriberade DNA:t. Samma fenomen är också karakteristiska för eukaryota RNA-polymeraser. Övergången från initiering till förlängning åtföljs av brott av bindningar mellan enzymet, promotorn, traoch i vissa fall av övergången av RNA-polymeras till ett tillstånd av förlängningskompetens. Förlängningsfasen slutar efter att det växande transkriptet har frigjorts och enzymet dissocierar från mallen.
Under förlängningsstadiet är ungefär 18 nukleotidpar otvinnade i DNA. Cirka 12 nukleotider av DNA-mallsträngen bildar en hybridhelix med den växande änden av RNA-strängen. När RNA-polymeras rör sig genom mallen sker avlindning av DNA-dubbelhelixen före den, och återställande av DNA-dubbelhelixen sker bakom den. Samtidigt frigörs nästa länk i den växande RNA-kedjan från komplexet med mallen och RNA-polymeraset. Dessa rörelser måste åtföljas av relativ rotation av RNA-polymeras och DNA. Det är svårt att föreställa sig hur detta kunde hända i en cell, speciellt under kromatintranskription. Därför är det möjligt att för att förhindra sådan rotation åtföljs RNA-polymeras som rör sig längs DNA av topoisomeraser.
Förlängning utförs med hjälp av grundläggande förlängningsfaktorer, som är nödvändiga för att processen inte ska stoppa i förtid.
Nyligen har bevis dykt upp som visar att regulatoriska faktorer också kan reglera förlängning. Under förlängningsprocessen pausar RNA-polymeras vid vissa delar av genen. Detta ses särskilt tydligt vid låga koncentrationer av substrat. I vissa områden av matrisen finns det långa förseningar i utvecklingen av RNA-polymeras, den så kallade. pauser observeras även vid optimala substratkoncentrationer. Varaktigheten av dessa pauser kan styras av förlängningsfaktorer.
| Tryptofan operon |
Transkription är syntesprocessenmolekylerRNA påområdeDNA, används som en matris. Betydelsen av transkriptionen är överföring av genetisk information från DNA till RNA.
En DNA-molekyl består av två komplementära strängar, medan RNA endast består av en. Under transkription fungerar endast en av DNA-strängarna som mall för RNA-syntes. De ringer henne semantisk kedja. Undantaget är mitokondrie-DNA, där båda strängarna är sensuella och innehåller olika gener. Förutom att vara ett undantag på nukleärt DNA, kan vissa gener vara lokaliserade på en nonsenssträng.
Under transkriptionen syntetiseras RNA-molekylen i riktningen från 5" till 3"-änden (vilket är naturligt för syntesen av alla nukleinsyror), medan syntesen längs DNA-kedjan fortsätter i motsatt riktning: 3"→5 ".
I eukaryoter transkriberas varje gen separat. Undantaget är återigen mitokondriellt DNA, som transkriberas till ett gemensamt multigentranskript, som sedan skärs. Eftersom gener i prokaryoter bildar grupper och bildar ett operon, transkriberas sådana gener tillsammans. I alla fall transkripton kallas en sektion av DNA som består av en promotor, en transkriberad region och en terminator.
Det finns 3 stadier i transkriptionen: initiering, förlängning, avslutning.
Initiering transkription gör att syntesen av RNA-molekylen kan börja. Initiering involverar bindning av ett komplex av enzymer till promotorn. Den viktigaste är RNA-polymeras (i detta fall DNA-beroende), som i sin tur består av flera subenhetsproteiner och spelar rollen som en katalysator för processen. Hos eukaryoter påverkas initieringen av transkription av speciella sektioner av DNA: förstärkare (stärka) och ljuddämpare (undertryckta), som vanligtvis är belägna en bit från själva genen. Det finns olika proteinfaktorer som påverkar möjligheten till transkriptionsinitiering.
Prokaryoter har bara en typ av RNA-polymeras, medan eukaryoter har tre. RNA-polymeras 1 används för att syntetisera tre typer av ribosomalt RNA (det finns totalt fyra typer av rRNA). RNA-polymeras 2 används för att syntetisera pre-mRNA (precursor messenger RNA). RNA-polymeras-3 syntetiserar en av typerna av ribosomalt RNA, transport och små nukleära.
RNA-polymeras kan känna igen specifika nukleotidsekvenser och fäster till dem. Dessa sekvenser är korta och universella för allt levande.
Efter att RNA-polymeras binder till promotorn lindas en sektion av DNA-dubbelhelixen upp och nukleotidbindningarna mellan strängarna i denna sektion bryts. Ungefär 18 par nukleotider löses upp.
På scenen förlängning sekventiell addition sker enligt principen om komplementaritet av fria nukleotider till den frigjorda DNA-sektionen. RNA-polymeras kombinerar nukleotider till en polyribonukleotidkedja.
Under RNA-syntes är cirka 12 av dess nukleotider komplementära och tillfälligt kopplade till DNA-nukleotider. När RNA-polymeras rör sig framför det divergerar DNA-kedjorna, och bakom det "sys" de med hjälp av enzymer. RNA-kedjan växer gradvis och rör sig ut ur RNA-polymeraskomplexet.
Det finns förlängningsfaktorer som förhindrar för tidig avbrytning av transkription.
Uppsägning Transkriptionsprocessen sker i terminatorregionen, som känns igen av RNA-polymeras tack vare speciella proteintermineringsfaktorer.
Många adeninnukleotider (poly-A) är fästa vid 3"-änden av den syntetiserade RNA-molekylen för att förhindra dess enzymatiska nedbrytning. Ännu tidigare, när 5"-änden syntetiserades, har den s.k. keps.
I de flesta fall producerar inte transkription färdigt RNA. Rå-RNA behöver fortfarande gå igenom en process bearbetning, vid vilken dess modifieringsförändringar inträffar och den blir funktionellt aktiv. Varje typ av RNA i eukaryoter genomgår sina egna modifieringar. Bildandet av poly-A och lock kallas också ofta för bearbetning.
Vi rekommenderar också
 Numerisk symbolik i Johannes Uppenbarelsebok
Numerisk symbolik i Johannes Uppenbarelsebok
 Träningsklass "Konfliktologi": Arabisk-israelisk konflikt
Träningsklass "Konfliktologi": Arabisk-israelisk konflikt
 Mirakel med ormen som hände den helige store martyren George
Mirakel med ormen som hände den helige store martyren George
 RNA tar emot ärftlig information
RNA tar emot ärftlig information
 Scaevola, hedersnamnet på krigets hjälte med etruskerna Gaius Mucius
Scaevola, hedersnamnet på krigets hjälte med etruskerna Gaius Mucius
 Var är predikatet? Vad är ett "predikat"? Typer av predikat. Är det sant att ett enkelt verbalt predikat är ett predikat vars lexikala och grammatiska betydelse uttrycks av ett verb
Var är predikatet? Vad är ett "predikat"? Typer av predikat. Är det sant att ett enkelt verbalt predikat är ett predikat vars lexikala och grammatiska betydelse uttrycks av ett verb