Funkcionalne regije tRNA. Zgradba in funkcije tRNA, značilnosti aktivacije aminokislin
70-90N | sekundarna stran - deteljica | CCA 3" const za vso tRNA |
prisotnost timina, psevdouridina-psi, digirouridina DGU v D-zanki - zaščita pred ribonukleazami? dolgoživi | Različne primarne strukture tRNA - 61 + 1 - po številu kodonov + formilmetionin tRNA, je mačji antikodon enak antikodonu metioninske tRNA. Različne terciarne strukture - 20 (glede na število aminokislin) | prepoznavanje - tvorba kovalentne vezi m-y tRNA in akt | aminoacil-tRNA sintetaze vežejo dejanja na tRNA
Naloga tRNA je prenos aminokislin iz citoplazme v ribosome, v katerih poteka sinteza beljakovin.
tRNA, ki vežejo eno aminokislino, se imenujejo izoakceptorji.
Skupno v celici hkrati obstaja 64 različnih tRNA.
Vsaka tRNA se pari samo s svojim kodonom.
Vsaka tRNA prepozna svoj kodon brez vpletenosti aminokisline. Aminokisline, vezane na tRNA, smo kemično modificirali, nato pa analizirali nastali polipeptid, ki je vseboval modificirano aminokislino. Cisteinil-tRNACys (R=CH2-SH) je bil reduciran v alanil-tRNACys (R=CH3).
Večina tRNA ima, ne glede na njihovo nukleotidno zaporedje, sekundarno strukturo v obliki deteljnega lista zaradi prisotnosti treh lasnic v njej.
Strukturne značilnosti tRNA
Na 3 "koncu molekule" so vedno štirje neparni nukleotidi in trije od njih so nujno CCA. 5" in 3" konca verige RNA tvorita akceptorsko steblo. Verige se držita skupaj zaradi komplementarnega parjenja sedem nukleotidov 5" - konec s sedmimi nukleotidi, ki se nahajajo blizu 3" konca. 2. Vse molekule imajo zatič T? C, ki je tako označena, ker vsebuje dva nenavadna ostanka: ribotimidin (T) in psevdouridin (? Lasnica je sestavljena iz dvojnega -verižno steblo petih parnih baz, vključno s parom G-C, in zanko dolžine sedmih nukleotidov.
na isti točki v zanki. 3. V antikodonski lasnici je steblo vedno predstavljeno z družino parnih
razlogov. Triplet, komplementarni sorodnemu kodonu, antikodonu, se nahaja v zanki.
le, sestavljen iz sedmih nukleotidov. Invariantna ura-
cyl in modificiran citozin, modificiran purin pa se praviloma bliža njegovemu 3" koncu
adenin. 4. Druga lasnica je sestavljena iz tri do štiri pare dolgih nukleotidov in spremenljive zanke
velikost, ki pogosto vsebuje uracil v zmanjšani obliki - dihidrouracil (DU). Najmočneje se razlikujejo nukleotidna zaporedja stebel, število nukleotidov med steblom antikodona in deblom T?C (variabilna zanka), pa tudi velikost zanke in lokalizacija ostankov dihidrouracila v zanki DU.
[Pevec, 1998].
Terciarna struktura tRNA
Struktura v obliki črke L.
Vezava aminokislin na tRNA
Da lahko aminokislina tvori polipeptidno verigo, mora biti vezana na tRNA z encimom aminoacil-tRNA sintetazo. Ta encim tvori kovalentno vez med aminokislinsko karboksilno skupino in ribozno hidroksilno skupino na 3' koncu tRNA s sodelovanjem ATP. Aminoacil-tRNA sintetaza ne prepozna specifičnega kodona zaradi prisotnosti antikodona na tRNA, temveč zaradi prisotnosti specifičnega prepoznavnega mesta na tRNA.
Skupno je v celici 21 različnih sintetaz aminoacil-tRNA.
Priključitev poteka v dveh fazah:
1. Karboksilna skupina aminokisline je vezana na ATP a-fosfat. Nastali nestabilen aminoacil adenilat se stabilizira z vezavo na encim.
2. Prenos aminoacilne skupine aminoacil adenilata na 2' ali 3'-OH skupino končne riboze tRNA
Nekatere sintetaze aminoacil-tRNA so sestavljene iz ene polipeptidne verige, druge pa iz dveh ali štirih enakih verig, vsaka z molekulsko maso od 35 do 115 kDa. Nekateri dimerni in tetramerni encimi so sestavljeni iz dveh vrst podenot. Ni jasne korelacije med velikostjo molekule encima ali naravo njegove strukture in specifičnosti podenote.
Specifičnost encima je določena z njegovo močno vezavo na akceptorski konec tRNA, regijo DU in spremenljivo zanko. Zdi se, da nekateri encimi ne prepoznajo antikodonskega tripleta in katalizirajo reakcijo aminoacetilacije, tudi če je antikodon spremenjen. Vendar pa nekateri encimi kažejo zmanjšano aktivnost glede na tako modificirane tRNA in pri zamenjavi antikodona dodajo napačno aminokislino.
70-90n | sekundarna stran - deteljica | CCA 3" const za vso tRNA |
prisotnost timina, psevdouridina-psi, digirouridina DGU v D-zanki - zaščita pred ribonukleazami? dolgoživi | Različne primarne strukture tRNA - 61 + 1 - po številu kodonov + formilmetionin tRNA, je mačji antikodon enak antikodonu metioninske tRNA. Različne terciarne strukture - 20 (glede na število aminokislin)
Obstajata dve vrsti tRNA, ki vežeta metionin tRNAFMet in tRNAMMet pri prokariotih in tRNAIMet in tRNAMMet pri evkariontih. Metionin se doda vsaki tRNA z uporabo ustrezne sinteze aminoacil-tRNA. metionin, vezan na tRNAFMet in tRNAIMet, tvori encim metionil-tRNA-transformilaza v Fmet-tRNAFMet. tRNA, obremenjene s formilmetioninom, prepoznajo iniciacijski kodon AUG.
Literatura:
Žal ni bibliografije.
Učbenik. Kljub temu, da je tRNA veliko manjša, si zgodba o njeni zgradbi, značilnostih in delovanju zasluži ločeno poglavje.
Torej je tRNA "prilagoditelj", ki prepozna tričrkovno zaporedje genetske kode na enem koncu in ga ujema z edino ustrezno aminokislino, ki je fiksirana na drugem koncu tRNA. Na koncu prenosne RNA, ki se dotakne nosilne RNA, so 3 nukleotidi, ki tvorijo antikodon. Le če je antikodon komplementaren regiji mRNA, se mu lahko pridruži prenosna RNA. Toda tudi v tem primeru se tRNA ne more sama pridružiti mRNA, potrebuje pomoč ribosoma, ki je mesto njihove interakcije, in aktivnega udeleženca pri prevajanju. Na primer, ribosom je tisti, ki ustvarja vezi med aminokislinami, ki jih prinaša tRNA, in tvorijo beljakovinsko verigo.
Strukturne značilnosti tRNA določa genetska koda, torej pravila za konstruiranje proteina glede na gen, ki ga bere prenosna RNA. Ta koda deluje v vsakem živem bitju na Zemlji: ustvarjanje virusa je zapisano v istih tričrkovnih kodonih, ki se uporabljajo za pisanje "navodil za sestavljanje" delfina. Eksperimentalno je bilo dokazano, da so geni enega živega bitja, nameščeni v celico drugega, odlično kopirani in prevedeni v beljakovine, ki se ne razlikujejo od genov, ki se razmnožujejo v celicah gostitelja. Enotnost genetske kode je osnova za proizvodnjo modificirane E. coli s kolonijami inzulina in mnogih drugih človeških encimov, ki se uporabljajo kot zdravila za ljudi, katerih telo jih ne zmore ali jih proizvaja premalo. Kljub očitni razliki med človekom in E. coli se človeške beljakovine zlahka ustvarijo iz človeških načrtov z uporabo kopirnega stroja E. coli. Ni presenetljivo, da se prenosne RNA različnih bitij zelo malo razlikujejo.

Vsak kodon s tega seznama, razen treh zaustavitveni kodoni, ki signalizira zaključek translacije, mora biti prepoznana s prenosno RNA. Prepoznavanje se izvede tako, da se na nosilno RNA pritrdi antikodon, ki se lahko veže le na en kodon s seznama, zato lahko tRNA prepozna samo en kodon. To pomeni, da je v celici vsaj 61 vrst teh molekul. Pravzaprav jih je še več, saj v nekaterih situacijah za branje sporočilne RNK ni dovolj le pravi antikodon: potrebni so drugi pogoji, v skladu s katerimi se ustvari posebna, modificirana tRNA.
Na prvi pogled bi morala takšna raznolikost tRNA bistveno zapletati proces prevajanja: navsezadnje bo vsaka od teh molekul preverila, ali je kodon matriksne RNA, ki ga nadomesti ribosom, skladnost s svojim antikodonom - zdi se, da je toliko nesmiselnega mehanskega dela , toliko izgubljenega časa in energije. Toda kot posledica evolucije so se oblikovali tudi celični mehanizmi, ki preprečujejo to težavo. Na primer, količina tRNA vsake vrste v celici ustreza temu, kako pogosto se aminokislina, ki jo nosi ta vrsta, nahaja v beljakovinah, ki se gradijo. Obstajajo aminokisline, ki jih celica redko uporablja, in so tiste, ki se pogosto uporabljajo, in če bi bilo število tRNA, ki jih nosijo, enako, bi to močno otežilo sestavljanje beljakovin. Zato je v celici malo "redkih" aminokislin in njihovih ustreznih tRNA, pogosto se pojavljajo pa se proizvajajo v velikih količinah.

S tako raznolikostjo molekul tRNA so si vse zelo podobne, zato bomo glede na njihovo strukturo in funkcije preučevali predvsem značilnosti, ki so skupne vsem vrstam. Ko pogledate 3D postavitev tRNA, je videti kot gost kup atomov. Zdi se neverjetno, da je ta zapleteno zvita molekula posledica zlaganja dolge verige nukleotidov, a tako nastane.
Možno je slediti fazam njegovega nastanka, začenši od prve: sestavljanje nukleotidnega zaporedja z RNA polimerazo v skladu z genom, ki vsebuje informacije o tej prenosni RNA. Imenuje se vrstni red, v katerem si ti nukleotidi sledijo in njihovo število primarna struktura tRNA. Izkazalo se je, da je primarna struktura tRNA kodirana v genu, ki ga bere RNA polimeraza. Na splošno je primarna struktura zaporedje sorazmerno preprostih molekul iste vrste, iz katerih je sestavljena bolj zapletena, zložena polimerna molekula. Na primer, primarna struktura beljakovinske molekule je preprosto zaporedje njenih sestavnih aminokislin.

Nobena veriga nukleotidov v celici ne more biti v razgrnjenem stanju, preprosto raztegnjena v vrstici. Na robovih nukleotidov je preveč pozitivno in negativno nabitih delov, ki med seboj zlahka tvorijo vodikove vezi. Kako nastanejo enake vezi med nukleotidi dveh molekul DNK, ki ju povezujejo v dvojno vijačnico, je opisano v, za podrobnosti o vodikovih vezi pa se lahko povzpnete. Vodikove vezi so manj močne kot vezi med atomi v molekulah, vendar so dovolj, da namišljeno zasukajo verigo tRNA in jo obdržijo v tem položaju. Sprva se te vezi tvorijo le med nekaterimi nukleotidi, ki zložijo tRNA v obliko listov detelje. Rezultat tega začetnega zlaganja se imenuje sekundarna struktura tRNA. Diagram na levi kaže, da so le nekateri nukleotidi povezani z vodikovimi vezmi, drugi pa ostanejo neparni in tvorijo obroče in zanke. Razlike med sekundarno strukturo različnih vrst tRNA so posledica razlik v njihovi primarni strukturi. To se kaže v različnih dolžinah »listov detelje« ali »peclja« zaradi različnih dolžin začetne verige nukleotidov.
Druga razlika v primarni strukturi različnih tRNA je, da imajo le na nekaterih položajih enake nukleotide (na zgornjem diagramu so označeni s prvimi črkami svojih imen), medtem ko se večina nukleotidov v različnih tRNA med seboj razlikuje. Zgornja shema je skupna vsem tRNA, zato so različni nukleotidi označeni s številkami.
Glavni funkcionalni deli tRNA so:
=) antikodon, to je nukleotidno zaporedje, ki je komplementarno enemu kodonu sporočilne RNA, ki se nahaja na antikodonska lasnica
=) akceptorskega konca na katero se lahko veže aminokislina. Nahaja se na nasprotni strani antikodonske lasnice.
V resnici niti ena tRNA ni videti tako, kot je v diagramu sekundarne strukture, ker so se le nekateri nukleotidi združili, da jo tvorijo, medtem ko so ostali ostali neparni. Zaradi tvorbe vodikovih vezi med nukleotidi iz različnih delov deteljnega lista se dodatno zloži v veliko bolj kompleksno terciarna struktura v obliki črke L. Natančno lahko razumete, kako so se različni deli sekundarne strukture ukrivili, da tvorijo terciarno strukturo, tako da uskladite barve na njihovih spodnjih diagramih. Antikodonski lasnik, označen z modro in sivo, ostane na dnu (vredno je spomniti, da je to "dno" pogojno: priročno je prikazati tRNA v tej prostorski orientaciji v shemah prevajanja beljakovin), konec akceptorja (rumen) je upognjen na stran.

Takole izgleda tRNA, ko je pripravljena pritrditi aminokislino. tRNA se ne more sama združiti z aminokislino, to zahteva sodelovanje posebnega encima: aminoacil-tRNA sintetaze. Število vrst sintetaze v celici sovpada s številom tipov tRNA.
Enakomernost oblike vseh vrst tRNA je nujna, da ribosom lahko prepozna katero koli od njih, olajša njihovo spajanje z mRNA in se v sebi premika z enega mesta na drugo. Če bi se različne vrste tRNA med seboj bistveno razlikovale, bi to izredno otežilo delo ribosoma, kar bi kritično zmanjšalo hitrost sinteze beljakovin. Naravna selekcija si tako prizadeva narediti tRNA podobne med seboj. Toda hkrati obstaja še en dejavnik, ki zahteva obstoj opaznih razlik med različnimi vrstami tRNA: navsezadnje je treba vsako vrsto prepoznati in ji pritrditi edino ustrezno aminokislino. Očitno bi morale biti te razlike opazne, vendar ne preveč bistvene, da se delo prepoznavanja vrst tRNA spremeni v nakitni proces. In prav to izvajajo aminoacil-tRNA sintetaze: vsaka od njih se lahko veže samo na eno od 20 aminokislin in jo pritrdi natančno na tiste vrste tRNA, ki ustrezajo tej aminokislini. Iz tabele z genetsko kodo je razvidno, da je vsaka aminokislina kodirana z več nukleotidnimi zaporedji, zato bo na primer vse štiri tRNA z antikodoni CGA, CGG, CGU in CGC prepoznala ista sintetaza, ki veže alanin njim. Takšne tRNA, ki jih obdela ena sintetaza, se imenujejo povezane.
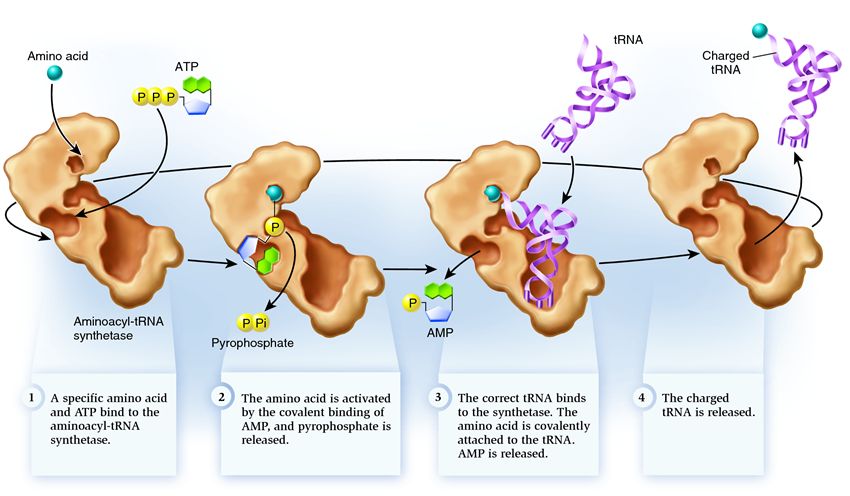
Sintetaza spada v skupino encimov, katerih naloga je, da se vežejo na ločeno obstoječe molekule in jih združijo v eno:
1 . sintetaza povezuje aminokislino in molekulo ATP. Dve fosfatni skupini se odcepita od ATP in sprostita energijo, potrebno za naslednje dejavnosti. Adenozin monofosfat (AMP), ki ostane od uničene molekule, se pritrdi na aminokislino in jo pripravi za povezavo z akceptorskim lasnikom.
2 . sintetaza nase veže eno od sorodnih tRNA, ki ustreza tej aminokislini.

Na tej stopnji se preveri skladnost prenosne RNA s sintetazo. Obstaja več načinov prepoznavanja in vsaka sintetaza ima edinstveno kombinacijo. Vsaj en antikodonski nukleotid je vključen v interakcijo med sintetazo in tRNA. Preveriti je treba tudi akceptorski lasnik: določi se prisotnost specifičnih nukleotidov na njem, ki so skupni sorodnim tRNA, ki ustrezajo želeni aminokislini. Nukleotidi iz drugih delov tRNA lahko sodelujejo tudi pri ujemanju z vezavo na določena mesta sintetaze. Napačna tRNA se lahko na nek način ujema z želeno, vendar se bo zaradi nepopolnega ujemanja počasi in ohlapno pridružila sintetazi ter zlahka odpadla. In pravilna tRNA se bo hitro in trdno prilepila na sintetazo, zaradi česar se struktura sintetaze spremeni in začne proces aminoacilacija , to je vezava aminokisline na tRNA.

3 . aminoacilacija sestoji iz zamenjave molekule AMP, vezane na aminokislino, z molekulo tRNA. Po tej zamenjavi AMP zapusti sintetazo in tRNA se zadrži za zadnji pregled aminokislin. Če je vezana aminokislina prepoznana kot napačna, se bo ločila od tRNA, mesto aminokisline v sintetazi bo prazno in tam se lahko pridruži druga molekula. Nova aminokislina bo šla skozi faze povezovanja z ATP in tRNA ter bo tudi testirana. Če ni bilo napak, se tRNA, nabita z aminokislinami, sprosti: pripravljena je igrati svojo vlogo pri prevajanju beljakovin. In sintetaza je pripravljena na pritrditev novih aminokislin in tRNA in cikel se bo začel na novo.
Veliko je odvisno od pravilnega delovanja aminoacil-tRNA sintetaze: če na tej stopnji pride do okvare, bo na tRNA pritrjena napačna aminokislina. In vgrajen bo v beljakovino, ki raste na ribosomu, ker tRNA in ribosom nimata funkcije preverjanja ujemanja kodona in aminokisline. Posledice napake so lahko manjše ali katastrofalne, z naravno selekcijo pa so bitja z encimi, ki nimajo funkcije tovrstnih preverjanj, izpodrinila bolj prilagodljiva z različnimi možnostmi ujemanja med aminokislino in tRNA. Zato se v sodobnih celicah sintetaza v povprečju kombinira z napačno aminokislino v enem primeru od 50 tisoč, z napačno tRNA pa le enkrat na 100 tisoč priponk.

Nekatere aminokisline se med seboj razlikujejo le za nekaj atomov. Če pogledate njihove sheme, postane očitno, da je verjetnost zamenjave arginina z alaninom veliko manjša kot zamenjava izolevcina za levcin ali valin. Zato ima vsaka sintetaza, ki se veže na eno od podobnih aminokislin, dodatne mehanizme preverjanja. Tukaj je primer takšne prilagoditve v izolevcin-tRNA sintetazi:

Vsaka sintaza ima sintetični center pri katerem je aminokislina vezana na tRNA. Tja gre akceptorski lasnik tRNA, ki ga ujame sintetaza, prav tako aminokislina, ki je pripravljena reagirati z njo. Delo nekaterih sintetaz se konča takoj po povezavi aminokisline in tRNA. Toda sintetaza Ile-tRNA ima povečano možnost za napake zaradi obstoja drugih izolevcinu podobnih aminokislin. Zato ima tudi popravni center: že iz imena je jasno, kakšno vlogo ima v procesu povezovanja tRNA in aminokislin. Slika na desni prikazuje, da položaj konca lasnice akceptorja tRNA v sintetičnem središču Ile-tRNA sintetaze daje tej lasnici nenaraven upogib. Vendar sintetaza zadrži tRNA v tem položaju, dokler se nanjo ne pritrdi aminokislina. Po vzpostavitvi te povezave je potreba po iskanju akceptorskega lasnega zatiča v sintetičnem središču izčrpana in tRNA se izravna, tako da svoj konec z aminokislino, ki je nanjo pritrjeno, dobi v korekcijski center.

Seveda ima sintetični center tudi vlogo pri odkrivanju aminokislin, ki niso primerne za sintetazo. Da bi prišla vanj, mora molekula izpolnjevati številne pogoje, vključno s pravo velikostjo. Kljub temu, da levcin in izolevcin vsebujeta enako število atomov, je zaradi razlik v prostorski strukturi levcin večji. Zato ne more prodreti v sintetični center, katerega velikost ustreza bolj kompaktnemu izolevcinu, in se preprosto odbije od Ile-tRNA sintetaze.
Toda valin, ki je najmanjša od teh treh molekul s podobno atomsko strukturo, zlahka prevzame mesto izolevcina v sintetičnem središču, sintetaza pa ga pritrdi na tRNA. V tem primeru pride v poštev korekcijski center sintetaze. Če je lasnica za ravnanje pravilno napolnjena in nosi izolevcin, se ne more stisniti v središče korekcije: preprosto je premajhna za to molekulo. Tako izravnane tRNA nič več ne drži in se loči od sintetaze. Če pa je valin vezan na tRNA, zdrsne v korekcijski center, s čimer ohranja tRNA povezano z njim v sintetazi. Tako predolgo bivanje tRNA v notranjosti je signal napake za sintetazo, ki spreminja njeno prostorsko konfiguracijo. Kot rezultat:
=) valin se loči od tRNA in odstrani iz sintetaze
=) akceptorska lasnica se vrne na sintetično mesto in čaka na pritrditev na aminokislino
=) sintetaza se veže na novo aminokislino, z njo »napolni« tRNA in ponovno preveri, ali je bil za to uporabljen izolevcin.
Podoben mehanizem dvojnega prepoznavanja uporabljajo druge sintetaze.
Fizikalne in kemijske lastnosti DNK
Različni dejavniki, ki prekinjajo vodikove vezi (povečanje temperature nad 80 C, spremembe pH in ionske jakosti, delovanje sečnine ipd.), povzročajo denaturacijo DNK, t.j. sprememba prostorske razporeditve verig DNK brez prekinitve kovalentnih vezi. Dvojna vijačnica DNK je med denaturacijo v celoti ali delno razdeljena na njene sestavne verige. Denaturacijo DNK spremlja povečanje optične absorpcije v UV območju purinskih in pirimidinskih baz. Ta pojav se imenuje hiperkromni učinek . Denaturacija zmanjša tudi visoko viskoznost, ki je značilna za naravne raztopine DNK. Ko se prvotna struktura dvoverižne DNK obnovi, se zaradi renaturacije zmanjša absorpcija dušikovih baz pri 260 nm zaradi njihovega "zaščita". Ta pojav se imenuje hipokromni učinek .
"Razvijanje" vsake DNK v njene sestavne verige poteka v določenem temperaturnem območju. Sredina tega intervala se imenuje tališče. Temperatura taljenja DNK je pri standardnih pogojih (določen pH in ionska jakost) odvisna od razmerja dušikovih baz. G-C pari, ki vsebujejo tri vodikove vezi, so močnejši, zato je višja kot je vsebnost G-C parov v DNK, višje je tališče.
Funkcije DNK. V zaporedju nukleotidov v molekulah DNK so kodirane genetske informacije. Glavne funkcije DNK so, prvič, zagotoviti razmnoževanje same sebe v vrsti celičnih generacij in generacij organizmov, in drugič, zagotoviti sintezo beljakovin. Te funkcije so posledica dejstva, da molekule DNK v prvem primeru služijo kot matrica za replikacijo, t.j. kopiranje informacij v hčerinske molekule DNK, v drugem - za transkripcijo, t.j. za prekodiranje informacij v strukturo RNA.
riž. 5 Krivulja taljenja (denaturacija DNK)
Komplementarne verige DNK, ločene med denaturacijo, se lahko pod določenimi pogoji ponovno povežejo v dvojno vijačnico. Ta proces se imenuje RENATURATION. Če do denaturacije ni prišlo v celoti in vsaj nekaj baz ni izgubilo interakcije z vodikovimi vezmi, renaturacija poteka zelo hitro.
Citoplazma celic vsebuje tri glavne funkcionalne vrste RNA. To so sporočilne RNA - mRNA, ki delujejo kot predloge za sintezo beljakovin, ribosomske RNA - rRNA, ki delujejo kot strukturne komponente ribosomov, in prenosne RNA - tRNA, ki sodelujejo pri prevajanju (prevajanju) informacij o mRNA v zaporedje aminokislin v proteinu.
Tabela 2 prikazuje razlike med DNK in RNA glede na strukturo, lokalizacijo v celici in funkcije.
Tabela 2 Razlike med DNK in RNA
Prenos RNA, tRNA-ribonukleinska kislina, katere funkcija je transport AA do mesta sinteze beljakovin. Ima tipično dolžino od 73 do 93 nukleotidov in velikost približno 5 nm. tRNA so tudi neposredno vključene v rast polipeptidne verige, saj se združujejo – v kompleksu z aminokislino – na kodon mRNA in zagotavljajo konformacijo kompleksa, ki je potrebna za tvorbo nove peptidne vezi. Vsaka aminokislina ima svojo tRNA. tRNA je enoverižna RNA, vendar ima v svoji funkcionalni obliki konformacijo listov deteljice. AA je kovalentno vezan na 3" konec molekule z uporabo encima aminoacil-tRNA sintetaze, specifičnega za vsako vrsto tRNA. Na mestu C je antikodon, ki ustreza AA-te. tRNA sintetizira navadna RNA polimeraza v v primeru prokariotov in z RNA polimerazo III v primeru evkariontov Transkripti genov tRNA so podvrženi večstopenjski obdelavi, kar vodi do tvorbe prostorske strukture, značilne za tRNA.
Obdelava tRNA vključuje 5 ključnih korakov:
odstranitev 5" vodilnega nukleotidnega zaporedja;
odstranitev 3'-terminalne sekvence;
dodajanje zaporedja CCA na 3" koncu;
izrezovanje intronov (pri evkariontih in arhejah);
modifikacije posameznih nukleotidov.
Transport tRNA poteka po Ran-odvisni poti s sodelovanjem transportnega faktorja exportin t, ki prepozna značilno sekundarno in terciarno str-ru zrele tRNA: kratke dvoverižne odseke in pravilno obdelane 5 "- in 3" konča. Ta mehanizem zagotavlja, da se iz jedra izvozijo samo zrele tRNA.
62. Prevajanje - prepoznavanje kodonov mRNA
Translacija je sinteza beljakovin, ki jo izvajajo ribosomi iz aminokislin na predlogi mRNA (ali in RNA). Sestavni elementi translacijskega procesa: aminokisline, tRNA, ribosomi, mRNA, encimi za aminoacilacijo tRNA, faktorji prevajanja beljakovin (beljakovinski faktorji iniciacije, elongacije, terminacije - specifični ekstraribosomski proteini, potrebni za procese prevajanja), viri energije ATP in GTP , magnezijevi ioni (stabilizirajo strukturo ribosomov). Pri sintezi beljakovin sodeluje 20 aminokislin. Da bi aminokislina »prepoznala« svoje mesto v prihodnji polipeptidni verigi, se mora vezati na prenosno RNA (tRNA), ki opravlja funkcijo adapterja. tRNA, ki se veže na aminokislino, nato prepozna ustrezen kodon na mRNA. prepoznavanje kodona mRNA:
Interakcija kodon-antikodon temelji na načelih komplementarnosti in antiparalelnosti:
3'----C - G-A*------5' tRNA antikodon
5'-----G-C-Y*------3' mRNA kodon
Hipotezo nihanja je predlagal F. Crick:
3'-baza kodona mRNA ni strogo povezana s 5'-bazo antikodona tRNA: na primer, Y (mRNA) lahko medsebojno deluje z A in G (tRNA)
Nekatere tRNA se lahko združijo z več kot enim kodonom.
63. Značilnosti sestavnih elementov prevajalskega postopka. Translacija (translatio-translation) je proces sinteze beljakovin iz aminokislin na matriksu informacijske (matriksne) RNA (mRNA, mRNA), ki ga izvaja ribosom.
Sinteza beljakovin je osnova življenja celic. Za izvajanje tega procesa v celicah vseh organizmov obstajajo posebne organele - ribosomi- ribonukleoproteinski kompleksi, zgrajeni iz 2 podenot: velike in majhne. Funkcija ribosomov je prepoznati tri črke (tri nukleotide) kodonov mRNA, ki jih primerja z ustreznimi nosilci antikodonov tRNA amino kisline in dodajanje teh aminokislin rastoči beljakovinski verigi. Ko se premika vzdolž molekule mRNA, ribosom sintetizira protein v skladu z informacijami, ki jih vsebuje molekula mRNA.
Za prepoznavanje AK-t v celici obstajajo posebni "adapterji", prenos molekul RNA(tRNA). Te molekule v obliki deteljnega lista imajo mesto (antikodon), komplementarno kodonu mRNA, pa tudi drugo mesto, na katerega je vezana aminokislina, ki ustreza temu kodonu. Vezava aminokislin na tRNA poteka v energijsko odvisni reakciji s pomočjo encimov aminoacil-tRNA sintetaze, nastala molekula pa se imenuje aminoacil-tRNA. Tako je specifičnost prevajanja določena z interakcijo med kodonom mRNA in antikodonom tRNA, pa tudi s specifičnostjo sintetaz aminoacil-tRNA, ki vežejo aminokisline strogo na svoje ustrezne tRNA (na primer, kodon GGU bo ustrezal tRNA, ki vsebuje antikodon CCA in samo AK glicin).
prokariontski ribosom
5S in 23S rRNA 16S rRNA
34 veveric 21 veveric
Prokariontski ribosomi imajo konstanto sedimentacije 70S, zato jih imenujemo delci 70S. Zgrajene so iz dveh različnih podenot: podenot 30S in 50S. Vsaka podenota je kompleks rRNA in ribosomskih proteinov.
Delec 30S vsebuje eno molekulo 16S rRNA in v večini primerov eno proteinsko molekulo iz več kot 20 vrst (21). Podenota 50S je sestavljena iz dveh molekul rRNA (23S in 5S). Sestavljen je iz več kot 30 različnih beljakovin (34), ki jih praviloma predstavlja en izvod. Večina ribosomskih beljakovin opravlja strukturno funkcijo.
evkariontskega ribosoma
5S; 5,8S in 28S rRNA 18S rRNA
najmanj 50 beljakovin najmanj 33 beljakovin
Ribosom je sestavljen iz velikih in majhnih podenot. Osnova strukture vsake podenote je kompleksno zvita rRNA. Proteini ribosoma so bili pritrjeni na oder rRNA.
Sedimentacijski koeficient popolnega evkariontskega ribosoma je približno 80 Svedbergovih enot (80S), sedimentacijski koeficient njegovih poddelcev pa 40S in 60S.
Manjša podenota 40S je sestavljena iz ene molekule 18S rRNA in 30-40 proteinskih molekul. Velika podenota 60S vsebuje tri vrste rRNA s sedimentacijskimi koeficienti beljakovin 5S, 5,8S in 28S ter 40-50 (na primer ribosomi hepatocitov podgan vključujejo 49 beljakovin).
Funkcionalne regije ribosomov
P - peptidilno mesto za peptidil tRNA
A - aminoacilno mesto za aminoacil tRNA
E – mesto za sproščanje tRNA iz ribosoma
Ribosom vsebuje 2 funkcionalni mesti za interakcijo s tRNA: aminoacil (akceptor) in peptidil (donor). Aminoacil-tRNA vstopi v akceptorsko mesto ribosoma in v interakciji tvori vodikove vezi med kodonom in antikodonskim trojčkom. Po tvorbi vodikovih vezi sistem napreduje za 1 kodon in konča na mestu darovalca. Hkrati se na izpraznjenem akceptorskem mestu pojavi nov kodon, na katerega se pritrdi ustrezna aminoacil-t-RNA.
Ribosomi: zgradba, funkcija
Ribosomi so citoplazemski centri biosinteze beljakovin. Sestavljeni so iz velikih in majhnih podenot, ki se razlikujejo po sedimentacijskih koeficientih (hitrost sedimentacije med centrifugiranjem), izraženih v enotah Svedberg - S.
Ribosomi so prisotni tako v evkariontskih kot v prokariontskih celicah, saj opravljajo pomembno funkcijo v biosinteza beljakovin. Vsaka celica vsebuje desetine, stotine tisoč (do več milijonov) teh majhnih zaobljenih organelov. Je zaobljen ribonukleoproteinski delec. Njegov premer je 20-30 nm. Ribosom je sestavljen iz velikih in majhnih podenot, ki se razlikujejo po sedimentacijskih koeficientih (hitrost sedimentacije med centrifugiranjem), izraženih v Svedbergovih enotah – S. Te podenote se združijo v prisotnosti verige m-RNA (matrična ali informacijska RNA). Kompleks skupine ribosomov, ki jih združuje ena sama molekula mRNA kot niz kroglic, se imenuje polisom. Te strukture so bodisi prosto nameščene v citoplazmi bodisi pritrjene na membrane zrnatega ER (v obeh primerih na njih aktivno poteka sinteza beljakovin).
Polisomi granularnega ER tvorijo beljakovine, ki se izločajo iz celice in uporabljajo za potrebe celotnega organizma (na primer prebavni encimi, beljakovine materinega mleka). Poleg tega so ribosomi prisotni na notranji površini mitohondrijske membrane, kjer tudi aktivno sodelujejo pri sintezi beljakovinskih molekul.
Prenosna RNA (tRNA) ima pomembno vlogo v procesu uporabe dednih informacij v celici. Za dostavo potrebnih aminokislin na mesto sestavljanja peptidnih verig tRNA deluje kot translacijski posrednik.
Molekule tRNA so polinukleotidne verige, sintetizirane na specifičnih zaporedjih DNK. Sestavljeni so iz relativno majhnega števila nukleotidov -75-95. Zaradi komplementarne povezave baz, ki se nahajajo v različnih delih polinukleotidne verige tRNA, pridobi strukturo, ki po obliki spominja na list detelje (slika 3.26).
riž. 3.26. Struktura tipične molekule tRNA.
Ima štiri glavne dele, ki opravljajo različne funkcije. sprejemnik"Steblo" tvorita dva komplementarno povezana terminalna dela tRNA. Sestavljen je iz sedmih baznih parov. 3'-konec tega stebla je nekoliko daljši in tvori enoverižno regijo, ki se konča s sekvenco CCA s prosto OH skupino. Na ta konec je pritrjena transportna aminokislina. Preostale tri veje so komplementarne parne nukleotidne sekvence, ki se končajo v neparnih odsekih, ki tvorijo zanke. Sredina teh vej - antikodon - je sestavljena iz petih parov nukleotidov in vsebuje antikodon v središču svoje zanke. Antikodon so trije nukleotidi, komplementarni kodonu mRNA, ki kodira aminokislino, ki jo ta tRNA prenaša na mesto sinteze peptidov.
Med akceptorsko in antikodonsko vejo sta dve stranski veji. V svojih zankah vsebujejo modificirane baze - dihidrouridin (D-zanka) in triplet TψC, kjer je \y psevdouriain (T^C-zanka).
Med vejami aitikodona in T^C je dodatna zanka, ki vključuje od 3-5 do 13-21 nukleotidov.
Na splošno je za različne vrste tRNA značilna določena konstantnost nukleotidnega zaporedja, ki ga najpogosteje sestavlja 76 nukleotidov. Sprememba njihovega števila je predvsem posledica spremembe števila nukleotidov v dodatni zanki. Komplementarne regije, ki podpirajo strukturo tRNA, so običajno ohranjene. Primarna struktura tRNA, določena z zaporedjem nukleotidov, tvori sekundarno strukturo tRNA, ki ima obliko lista detelje. Po drugi strani pa sekundarna struktura povzroči tridimenzionalno terciarno strukturo, za katero je značilna tvorba dveh pravokotnih dvojnih vijačnic (slika 3.27). Eno od njih tvorita akceptorska in TψC veja, drugo pa antikodon in D veje.
Na koncu ene od dvojnih vijačnic je transportirana aminokislina, na koncu druge pa antikodon. Ta območja so med seboj najbolj oddaljena. Stabilnost terciarne strukture tRNA se ohranja zaradi pojava dodatnih vodikovih vezi med bazami polinukleotidne verige, ki se nahajajo na različnih delih le-te, a prostorsko blizu v terciarni strukturi.
Različne vrste tRNA imajo podobno terciarno strukturo, čeprav z nekaterimi različicami.
riž. 3.27. Prostorska organizacija tRNA:
I - sekundarna struktura tRNA v obliki "deteljnega lista", ki jo določa njena primarna struktura (zaporedje nukleotidov v verigi);
II - dvodimenzionalna projekcija terciarne strukture tRNA;
III - razporeditev molekule tRNA v vesolju
PRILOGA (če kdo tega ne razume)
Svetlobni zobje - nukleotidi (adenin-timin / uracil /, gvanin-citazin). Vse strele so DNK.

Za prenos informacij iz DNK morate prekiniti 2 verigi. Vez med A-T in G-C je vodik, zato jo encim Helicase zlahka prekine:

Da preprečim nastanek vozlov (kot primer sem zvil brisačo):



Topoizomeraza prereže eno verigo DNK v izvoru replikacije, tako da se veriga ne zvije.

Ko je ena nit prosta, se lahko druga zlahka vrti okoli svoje osi in s tem razbremeni napetost med "odvijanjem". Vozlišča se ne prikažejo, energija je prihranjena.
Nato je za začetek zbiranja RNA potreben RNA primer. Protein, ki sestavlja mRNA, ne more samo sestaviti prvega nukleotida, za začetek potrebuje kos RNA (tam je podrobno napisano, bom napisal kasneje). Ta del se imenuje RNA primer. In ta protein nanj že veže prvi nukleotid.


Pri opisu zgradbe nukleinskih kislin upoštevamo različne ravni organiziranosti makromolekul: primarno in sekundarno strukturo.
Primarna struktura nukleinskih kislin je nukleotidna sestava in določeno zaporedje nukleotidnih enot v polimerni verigi.
Sekundarna struktura RNA. Molekula ribonukleinske kisline je zgrajena iz ene same polinukleotidne verige.
Sekundarna struktura RNA
Ločeni odseki verige RNA tvorijo spiralizirane zanke - "lasnice", zaradi vodikovih vezi med komplementarnima dušikovima bazama A-U in G-C. Odseki verige RNA v takšnih spiralnih strukturah so antiparalelni, vendar ne vedno popolnoma komplementarni; vsebujejo neparne nukleotidne ostanke ali celo enoverižne zanke, ki se ne prilegajo dvojni vijačnici. Prisotnost spiraliziranih regij je značilna za vse vrste RNA.
Glavna vloga RNA je neposredno sodelovanje pri biosintezi beljakovin.
Poznane so tri vrste celične RNA, ki se razlikujejo po lokaciji v celici, sestavi, velikosti in lastnostih, ki določajo njihovo specifično vlogo pri tvorbi beljakovinskih makromolekul:
- informacijske (matrične) RNA prenašajo v DNK kodirane informacije o strukturi proteina iz celičnega jedra do ribosomov, kjer poteka sinteza beljakovin; primarna struktura vseh mRNA, ne glede na edinstvenost njihovega kodirnega zaporedja, ima enako strukturo 5'- in 3'-koncev.
Torej, na 5'-koncu je modificiran nukleotid 7-metilgvanozin-5'-trifosfat (cap). Več deset nukleotidov loči pokrovček od iniciacijskega kodona, običajno triplet -AUG-. Kodirnemu območju sledi eden od končnih kodonov -UGA-, -UUA-, -UAG-. Na 3' koncu večine mRNA je nukleotidno zaporedje 100-200 ostankov adenozin monofosfata.
- prenosne RNA zbirajo aminokisline v citoplazmi celice in jih prenašajo v ribosom; Tovrstne molekule RNA se iz ustreznih odsekov verige sporočilne RNA "učijo", katere aminokisline naj sodelujejo pri sintezi beljakovin.
Prostorsko strukturo katere koli tRNA, ne glede na razlike v nukleotidnem zaporedju, opisuje univerzalni model deteljice. Vsaka molekula tRNA ima verižne odseke, ki ne sodelujejo pri tvorbi vodikovih vezi med nukleotidnimi ostanki.
Ti vključujejo zlasti mesto, ki je odgovorno za vezavo na aminokislino na 3'-koncu molekule in antikodon, specifičen triplet nukleotidov, ki komplementarno deluje s kodonom mRNA.
- ribosomska RNA zagotavlja sintezo beljakovin določene strukture, branje informacij iz informacijske (matriksne) RNA. rRNA tvorijo komplekse z beljakovinami, imenovanimi ribosomi.
Vsak ribosom je sestavljen iz dveh podenot - majhne (40S) in velike (60S). Podenote ribosoma se ne razlikujejo le po naboru rRNA, temveč tudi po številu in strukturi beljakovin.
Datum objave: 2015-02-03; Prebrano: 2729 | Kršitev avtorskih pravic strani

RNA je polimer, katerega monomeri so ribonukleotidi.
Za razliko od DNK RNA ne tvorita dve, ampak ena polinukleotidna veriga (izjema - nekateri virusi, ki vsebujejo RNA, imajo dvoverižno RNA). Nukleotidi RNA so sposobni tvoriti vodikove vezi med seboj. Verige RNA so veliko krajše od verig DNK.
Monomer RNA - nukleotid (ribonukleotid) - je sestavljen iz ostankov treh snovi: 1) dušikove baze, 2) petoogljičnega monosaharida (pentoze) in 3) fosforne kisline. Dušikove baze RNA sodijo tudi v razrede pirimidinov in purinov.
Pirimidinske baze RNA - uracil, citozin, purinske baze - adenin in gvanin.
31. Vrste RNA in značilnosti njene strukture
RNA nukleotidni monosaharid je predstavljen z ribozo.
Obstajajo tri vrste RNA: 1) informacijska (matrična) RNA - mRNA (mRNA), 2) prenosna RNA - tRNA, 3) ribosomska RNA - rRNA.
Vse vrste RNA so nerazvejani polinukleotidi, imajo specifično prostorsko konformacijo in sodelujejo v procesih sinteze beljakovin.
Informacije o strukturi vseh vrst RNA so shranjene v DNK. Proces sinteze RNA na predlogi DNK se običajno imenuje transkripcija.

Prenosne RNA običajno vsebujejo 76 (od 75 do 95) nukleotidov; molekulska masa - 25.000–30.000.
tRNA predstavlja približno 10 % celotne vsebnosti RNA v celici. Funkcije tRNA: 1) transport aminokislin na mesto sinteze beljakovin, do ribosomov, 2) translacijski mediator. V celici najdemo približno 40 vrst tRNA, vsaka od njih ima samo zanjo značilno nukleotidno zaporedje. Hkrati imajo vse tRNA več intramolekularnih komplementarnih regij, zaradi katerih tRNA pridobijo konformacijo, ki po obliki spominja na list detelje.
Vsaka tRNA ima zanko za stik z ribosomom (1), antikodonsko zanko (2), zanko za stik z encimom (3), akceptorsko steblo (4) in antikodon (5). Aminokislina je pritrjena na 3' konec akceptorskega stebla. Antikodon - trije nukleotidi, ki "prepoznajo" kodon mRNA.
Poudariti je treba, da lahko določena tRNA prenaša strogo določeno aminokislino, ki ustreza njenemu antikodonu. Specifičnost povezave aminokislin in tRNA je dosežena zaradi lastnosti encima aminoacil-tRNA sintetaze.
Ribosomske RNA vsebujejo 3000–5000 nukleotidov; molekulska masa - 1.000.000–1.500.000.
rRNA predstavlja 80–85 % celotne vsebnosti RNA v celici. V kompleksu z ribosomskimi beljakovinami rRNA tvori ribosome - organele, ki izvajajo sintezo beljakovin. V evkariontskih celicah pride do sinteze rRNA v nukleolu. Funkcije rRNA: 1) nujna strukturna komponenta ribosomov in s tem zagotavlja delovanje ribosomov; 2) zagotavljanje interakcije ribosoma in tRNA; 3) začetna vezava ribosoma in iniciatorskega kodona mRNA ter določitev bralnega okvirja, 4) nastanek aktivnega centra ribosoma.
Messenger RNA so raznolike po vsebnosti nukleotidov in molekulski masi (od 50.000 do 4.000.000).
Delež mRNA predstavlja do 5 % celotne vsebnosti RNA v celici. Funkcije mRNA: 1) prenos genetskih informacij iz DNK na ribosome, 2) matriks za sintezo proteinske molekule, 3) določitev aminokislinskega zaporedja primarne strukture proteinske molekule.
Preberite tudi
RNA je polimer, katerega monomeri so nukleotidi.
Tri dušikove baze so enake kot v DNK (adenin, gvanin, citozin); četrti - uracil - je prisoten v molekuli RNA namesto timina. Nukleotidi RNA vsebujejo ribozo namesto deoksiriboze. V verigi RNA ...
tri glavne vrste RNA: informativni(mRNA) oz matriko(mRNA), ribosomski(rRNA) in transport(tRNA). Razlikujejo se po velikosti molekul in funkciji. Vse vrste RNA se sintetizirajo na DNK s sodelovanjem encimov - RNA polimeraz. Messenger RNA predstavlja 2-3% vse celične RNA, ribosomska - 80-85, transportna - približno 15%.
mRNA.
bere dedne informacije iz segmenta DNK in jih v obliki kopiranega zaporedja dušikovih baz prenese na ribosome, kjer se sintetizira določen protein. Vsaka od molekul mRNA po vrstnem redu nukleotidov in po velikosti ustreza genu v DNK, iz katerega je bila prepisana. V povprečju mRNA vsebuje 1500 nukleotidov (75-3000). Vsak triplet (trije nukleotidi) na mRNA se imenuje kodon. Od kodona je odvisno, katera aminokislina se bo pojavila na določenem mestu med sintezo beljakovin.
(tRNA) ima relativno nizko molekulsko maso približno 24-29 tisoč.
D in vsebuje od 75 do 90 nukleotidov v molekuli. Do 10% vseh nukleotidov tRNA je manjših baz, ki jo očitno ščitijo pred delovanjem hidrolitičnih encimov.Vloga tRNA je, da prenašajo aminokisline na ribosome in sodelujejo v procesu sinteze beljakovin. Vsaka aminokislina se veže na določeno tRNA. Številne aminokisline imajo več kot eno tRNA. Do danes je bilo odkritih več kot 60 tRNA, ki se razlikujejo po svoji primarni strukturi (bazno zaporedje).
Sekundarna struktura vseh tRNA je predstavljena v obliki deteljnega lista z dvoverižnim steblom in tremi enoverižnimi). Na koncu ene od verig je akceptorsko mesto - triplet CCA, na katerega je adenin vezana specifična aminokislina.
(rRNA). Vsebujejo 120-3100 nukleotidov. Ribosomska RNA se kopiči v jedru, v jedrih.
Ribosomski proteini se transportirajo v jedro iz citoplazme in tam pride do spontane tvorbe ribosomskih poddelcev z združevanjem beljakovin z ustrezno rRNA. Poddelci ribosoma se transportirajo skupaj ali ločeno skozi pore jedrske membrane v citoplazmo. ribosomi so organeli velikosti 20-30 nm.
Sestavljeni so iz dveh poddelcev različnih velikosti in oblik. Na določenih stopnjah sinteze beljakovin v celici se ribosomi razdelijo na poddelce.
Ribosomska RNA služi kot okvir za ribosome in olajša začetno vezavo mRNA na ribosom med biosintezo beljakovin.
6. vprašanje Vezi, ki tvorijo primarne in sekundarne strukture DNK in RNA. Vrste RNA
Genetska koda je način kodiranja aminokislinskega zaporedja beljakovin z uporabo zaporedja nukleotidov, značilnih za vse žive organizme.
Lastnosti: 1) genetska koda trojček(vsaka aminokislina je kodirana s tremi nukleotidi); 2) neprekrivanje(sosednji trojčki nimajo skupnih nukleotidov); 3) degenerirati(z izjemo metionina in triptofana imajo vse aminokisline več kot en kodon); 4) univerzalna(večinoma enako za vse žive organizme); 5) v kodonih za eno aminokislino sta prva dva nukleotida običajno enaka, tretji pa se razlikuje; 6) ima linearni vrstni red branja in je značilen po kolinearnost, t.
e. sovpadanje vrstnega reda kodonov v mRNA z vrstnim redom aminokislin v sintetizirani polipeptidni verigi.
Datum objave: 2014-12-08; Prebrano: 11268 | Kršitev avtorskih pravic strani
studopedia.org - Studopedia.Org - 2014-2018. (0,001 s) ...
Citoplazma celic vsebuje tri glavne funkcionalne vrste RNA:
- sporočilna RNA (mRNA), ki delujejo kot predloge za sintezo beljakovin;
- ribosomska RNA (rRNA), ki deluje kot strukturne komponente ribosomov;
- prenos RNA (tRNA), ki sodelujejo pri prevajanju (prevajanju) informacij o mRNA v aminokislinsko zaporedje proteinske molekule.
V jedru celic najdemo jedrsko RNA, ki predstavlja od 4 do 10 % celotne celične RNA.
Večino jedrske RNA predstavljajo visokomolekularni prekurzorji ribosomske in prenosne RNA. Prekurzorji rRNA z visoko molekulsko maso (28 S, 18 S in 5 S RNA) so v glavnem lokalizirani v jedrcu.
RNA je glavni genetski material v nekaterih živalskih in rastlinskih virusih (genomska RNA). Za večino RNA virusov je značilna reverzna transkripcija njihovega RNA genoma, ki jo usmerja reverzna transkriptaza.
Vse ribonukleinske kisline so polimeri ribonukleotidov, povezanih, kot v molekuli DNK, s 3′,5′-fosforodiesstrskimi vezmi.
Za razliko od DNK, ki ima dvoverižno strukturo, je RNA enoverižna linearna polimerna molekula.
struktura mRNA. mRNA je po velikosti in stabilnosti najbolj heterogen razred RNA.
struktura tRNA.
Prenosne RNA delujejo kot mediatorji (adapterji) med prevajanjem mRNA. Predstavljajo približno 15 % celotne celične RNA. Vsaka od 20 proteinogenih aminokislin ima svojo tRNA. Za nekatere aminokisline, ki jih kodirata dva ali več kodonov, obstaja več tRNA.
tRNA so relativno majhne enoverižne molekule, sestavljene iz 70-93 nukleotidov. Njihova molekulska masa je (2,4-3,1) 0,104 kDa.
Sekundarna struktura tRNA nastane zaradi tvorbe največjega števila vodikovih vezi med intramolekularnimi komplementarnimi pari dušikovih baz.
Zaradi tvorbe teh vezi se polinukleotidna veriga tRNA zasuka s tvorbo spiraliziranih vej, ki se končajo z zankami neparnih nukleotidov. Prostorska slika sekundarnih struktur vseh tRNA ima obliko deteljinega lista.
V "deteljici" ločimo štiri obvezne veje, daljše tRNA poleg tega vsebujejo kratko peto (dodatno) vejo.
Adaptersko funkcijo tRNA zagotavlja akceptorska veja, na 3'-konec katere je z etrsko vezjo pritrjen aminokislinski ostanek, in antikodonska veja nasproti akceptorske veje, na vrhu katere je zanka, ki vsebuje antikodon.
Antikodon je specifičen triplet nukleotidov, ki je komplementaren v antiparalelni smeri kodonu mRNA, ki kodira ustrezno aminokislino.
T-veja, ki nosi psevdouridinsko zanko (TyC-zanka), zagotavlja interakcijo tRNA z ribosomi.
D-veja, ki nosi dehidrouridinsko zanko, zagotavlja interakcijo tRNA z ustrezno aminoacil-tRNA sintetazo.

Sekundarna struktura tRNA
Funkcije pete dodatne veje so še vedno slabo razumljene, najverjetneje izenačuje dolžine različnih molekul tRNA.
Terciarna struktura tRNA je zelo kompaktna in nastane tako, da se posamezne veje deteljnega lista zaradi dodatnih vodikovih vezi tvorijo v L-oblikovno strukturo "komolca".
Transportna RNA, struktura in funkcionalni mehanizem.
V tem primeru se akceptorska roka, ki veže aminokislino, nahaja na enem koncu molekule, antikodon pa na drugem.

Terciarna struktura tRNA (po A.S. Spirinu)
Struktura rRNA in ribosomov. Ribosomske RNA tvorijo hrbtenico, na katero se vežejo specifični proteini in tvorijo ribosome. Ribosomi so nukleoproteinske organele, ki zagotavljajo sintezo beljakovin iz mRNA.
Število ribosomov v celici je zelo veliko: od 104 pri prokariotih do 106 pri evkariontih. Ribosomi so lokalizirani predvsem v citoplazmi, pri evkariontih, poleg tega v nukleolu, v mitohondrijskem matriksu in v stromi kloroplastov. Ribosomi so sestavljeni iz dveh poddelcev: velikega in majhnega. Po velikosti in molekulski masi so vsi preučevani ribosomi razdeljeni v 3 skupine - 70S ribosomi prokariotov (S-sedimentacijski koeficient), sestavljeni iz majhnih 30S in velikih 50S poddelcev; 80S evkariontski ribosomi, sestavljeni iz 40S majhnih in 60S velikih podenot.
Majhno podenoto ribosomov 80S tvorita ena molekula rRNA (18S) in 33 molekul različnih beljakovin.
Veliko podenoto tvorijo tri molekule rRNA (5S, 5,8S in 28S) in približno 50 beljakovin.
Sekundarna struktura rRNA nastane zaradi kratkih dvoverižnih odsekov molekule - lasnic (približno 2/3 rRNA), 1/3 - je predstavljena z enoverižnimi deli, bogatimi s purinskimi nukleotidi.
Socialni gumbi za Joomla
Beljakovine so osnova življenja. Njihove funkcije v celici so zelo raznolike. Vendar se beljakovine "ne morejo" razmnoževati. In vse informacije o strukturi beljakovin so vsebovane v genih (DNK).
Pri višjih organizmih se beljakovine sintetizirajo v citoplazmi celice, DNK pa je skrita za lupino jedra. Zato DNK ne more neposredno služiti kot predloga za sintezo beljakovin. To vlogo opravlja druga nukleinska kislina - RNA.
Molekula RNA je nerazvejen polinukleotid s terciarno strukturo.
Tvori ga ena polinukleotidna veriga in čeprav so komplementarni nukleotidi, ki so vključeni vanjo, sposobni tvoriti vodikove vezi med seboj, se te vezi pojavljajo med nukleotidi ene verige. Verige RNA so veliko krajše od verig DNK. Če je vsebnost DNK v celici razmeroma konstantna, potem vsebnost RNA močno niha. Največjo količino RNK v celicah opazimo med sintezo beljakovin.
RNA ima pomembno vlogo pri prenosu in izvajanju dednih informacij.
V skladu s funkcijo in strukturnimi značilnostmi ločimo več razredov celične RNA.
Obstajajo trije glavni razredi celične RNA.
- Informacijska (mRNA) ali matrična (mRNA). Njegove molekule so najbolj raznolike glede na velikost, molekulsko maso (od 0,05x106 do 4x106) in stabilnost.
Sestavljajo približno 2 % celotne količine RNA v celici. Vse mRNA so nosilci genetskih informacij od jedra do citoplazme, do mesta sinteze beljakovin. Služijo kot matrica (delovna risba) za sintezo beljakovinske molekule, saj določajo zaporedje aminokislin (primarna struktura) proteinske molekule.

- Ribosomska RNA (rRNA).
Predstavljajo 80–85 % celotne vsebnosti RNA v celici.
31. Struktura RNA. Vrste RNA, strukturne značilnosti in funkcije. Sekundarna struktura tRNA
Ribosomska RNA je sestavljena iz 3-5 tisoč nukleotidov. Sintetizira se v nukleolah jedra. V kompleksu z ribosomskimi proteini rRNA tvori ribosome - organele, na katerih so sestavljene beljakovinske molekule. Glavni pomen rRNA je v tem, da zagotavlja začetno vezavo mRNA in ribosoma ter tvori aktivno središče ribosoma, v katerem se med sintezo polipeptidne verige tvorijo peptidne vezi med aminokislinami.
- Prenosne RNA (tRNA).
Molekule tRNA običajno vsebujejo 75-86 nukleotidov. Molekulska teža molekul tRNA je približno 25 tisoč. Molekule tRNA igrajo vlogo posrednikov pri biosintezi beljakovin - dostavljajo aminokisline na mesto sinteze beljakovin, torej do ribosomov. Celica vsebuje več kot 30 vrst tRNA. Vsaka vrsta tRNA ima svoje edinstveno zaporedje nukleotidov.
Vse molekule pa imajo več intramolekularnih komplementarnih regij, zaradi katerih imajo vse tRNA terciarno strukturo, ki po obliki spominja na list detelje.
Sekundarna struktura RNA- značilna za tRNA, enoverižna, oblikovana kot "list detelje".
Vključuje:
- relativno kratke dvojne vijačnice - stebla,
- enoverižni odseki - zanke.
Obstajajo 4 stebla (akceptor, antikodon, dihidrouridil, psevdouridil) in 3 zanke.

"Stem-loop" - element sekundarne strukture RNA, shematično

"Pseudoknot" - element sekundarne strukture RNA, shematično

Akceptorsko steblo vsebuje 3'- in 5'-konca polinukleotidne verige, 5'-konec se konča z ostankom gvanilne kisline, 3'-konec je CCA triplet in služi za tvorbo estrske vezi z AA.
Deblo antikodona prepozna svoj kodon na mRNA v ribosomih po principu komplementarnosti.
Steblo psevdouridila služi za pritrditev na ribosom.
Dihidrouridilno steblo služi za vezavo na aminoacil-tRNA sintetazo.
Socialni gumbi za Joomla
Priporočamo tudi
 Stikalno napajanje: popravilo in izboljšanje
Stikalno napajanje: popravilo in izboljšanje
 Daljinski nadzor svetlobe
Daljinski nadzor svetlobe
 Učne ure plavanja za predšolske otroke
Učne ure plavanja za predšolske otroke
 Opombe za mojstra - domači gospodinjski alarmi
Opombe za mojstra - domači gospodinjski alarmi
 Propeler ure na Atmega8
Propeler ure na Atmega8
 Primeri uporabe naprav in relejev, kako izbrati in pravilno povezati rele Mikrokrmilnik in rele preprosta preklopna vezja
Primeri uporabe naprav in relejev, kako izbrati in pravilno povezati rele Mikrokrmilnik in rele preprosta preklopna vezja