tRNA-ի ֆունկցիոնալ շրջաններ. tRNA-ի կառուցվածքը և գործառույթները, ամինաթթուների ակտիվացման առանձնահատկությունները
70-90Ն | երկրորդական էջ - երեքնուկ | CCA 3» կոնստ բոլոր tRNA |
թիմինի, պսևդուրիդին-փսի, դիգիուրիդին DGU-ի առկայությունը D հանգույցում - պաշտպանություն ռիբոնուկլեազներից: երկարակյաց | tRNA-ի մի շարք առաջնային կառուցվածքներ - 61 + 1 - կոդոնների քանակով + ֆորմիլմեթիոնին tRNA-ն, կատվի հակակոդոնը նույնն է, ինչ մեթիոնինի tRNA-ն: Երրորդական կառուցվածքների բազմազանություն - 20 (ըստ ամինաթթուների քանակի) | ճանաչում - կովալենտային կապի ձևավորում m-y tRNA և ակտ | aminoacyl-tRNA սինթետազները կցում են ակտերը tRNA-ին
tRNA-ի գործառույթն է ցիտոպլազմից ամինաթթուների տեղափոխումը ռիբոսոմներ, որոնցում տեղի է ունենում սպիտակուցի սինթեզ:
tRNA-ները, որոնք կապում են մեկ ամինաթթու, կոչվում են isoacceptor:
Ընդհանուր առմամբ, բջջում միաժամանակ գոյություն ունի 64 տարբեր tRNA:
Յուրաքանչյուր tRNA զուգակցվում է միայն իր կոդոնի հետ:
Յուրաքանչյուր tRNA ճանաչում է իր սեփական կոդոնը առանց ամինաթթվի ներգրավման: tRNA-ի հետ կապված ամինաթթուները քիմիապես փոփոխվել են, որից հետո վերլուծվել է ստացված պոլիպեպտիդը, որը պարունակում է փոփոխված ամինաթթու: Ցիստեյնիլ-tRNACys (R=CH2-SH) վերածվել է ալանիլ-tRNACys (R=CH3):
tRNA-ների մեծ մասը, անկախ նուկլեոտիդային հաջորդականությունից, ունեն երեքնուկի տերևի երկրորդական կառուցվածք՝ դրա մեջ երեք մազակալների առկայության պատճառով։
tRNA-ի կառուցվածքային առանձնահատկությունները
Մոլեկուլի 3 «վերջում» միշտ կան չորս չզույգված նուկլեոտիդներ, և դրանցից երեքը պարտադիր կերպով CCA են: ՌՆԹ-ի շղթայի 5» և 3» ծայրերը կազմում են ընդունող ցողուն: յոթ նուկլեոտիդ 5" - ավարտվում է յոթ նուկլեոտիդով, որոնք գտնվում են 3" ծայրի մոտ: 2. Բոլոր մոլեկուլներն ունեն T?C մազակալ, որն այդպես է նշանակված, քանի որ այն պարունակում է երկու անսովոր մնացորդներ՝ ռիբոթիմիդին (T) և պսևդուրիդին (? Մազակալը բաղկացած է կրկնակի մազից: - հինգ զույգ հիմքերից բաղկացած ցողուն, ներառյալ G-C զույգը և երկարությամբ յոթ նուկլեոտիդներից բաղկացած հանգույց:
օղակի նույն կետում: 3. Հակակոդոնային մազակալում ցողունը միշտ ներկայացված է զույգերի ընտանիքով
հիմքերը. Կապակցված կոդոնին լրացնող եռյակը՝ հակակոդոնը, գտնվում է օղակում:
le, բաղկացած յոթ նուկլեոտիդներից։ Անփոփոխ ուրա-
ցիլը և փոփոխված ցիտոսինը, իսկ ձևափոխված պուրինը, որպես կանոն, հարում է դրա 3 «վերջին»
ադենին. 4. Մեկ այլ վարսահարդարիչ բաղկացած է երեքից չորս զույգ նուկլեոտիդների երկարությամբ ցողունից և փոփոխական օղակից:
չափը, որը հաճախ պարունակում է ուրացիլ կրճատված ձևով `դիհիդրոուրասիլ (DU): Ցողունների նուկլեոտիդային հաջորդականությունները, նուկլեոտիդների թիվը հակակոդոնային ցողունի և T?C ցողունի միջև (փոփոխական հանգույց), ինչպես նաև օղակի չափը և դիհիդրոուրացիլային մնացորդների տեղայնացումը DU հանգույցում ամենից խիստ տարբերվում են:
[Երգչուհի, 1998]։
tRNA-ի երրորդային կառուցվածքը
L-ձևավորված կառուցվածք:
Ամինաթթուների կցումը tRNA-ին
Որպեսզի ամինաթթուն ձևավորի պոլիպեպտիդային շղթա, այն պետք է միացվի tRNA-ին aminoacyl-tRNA սինթետազ ֆերմենտի միջոցով: Այս ֆերմենտը կովալենտային կապ է ստեղծում ամինաթթուների կարբոքսիլ խմբի և ռիբոզային հիդրոքսիլ խմբի միջև tRNA-ի 3' ծայրում՝ ATP-ի մասնակցությամբ։ Ամինոացիլ-tRNA սինթետազը ճանաչում է հատուկ կոդոն ոչ թե tRNA-ում հակակոդոնի առկայության պատճառով, այլ tRNA-ի վրա հատուկ ճանաչման վայրի առկայության պատճառով:
Ընդհանուր առմամբ, բջջում կա 21 տարբեր aminoacyl-tRNA սինթետազներ:
Միացումը տեղի է ունենում երկու փուլով.
1. Ամինաթթվի կարբոքսիլ խումբը կցվում է ATP ա-ֆոսֆատին։ Ստացված անկայուն ամինասիլ ադենիլատը կայունանում է ֆերմենտին միանալու միջոցով:
2. Ամինացիլ ադենիլատի ամինացիլ խմբի տեղափոխումը tRNA-ի վերջնական ռիբոզի 2' կամ 3'-OH խմբին.
Որոշ aminoacyl-tRNA սինթետազներ բաղկացած են մեկ պոլիպեպտիդային շղթայից, իսկ մյուսները բաղկացած են երկու կամ չորս նույնական շղթաներից, որոնցից յուրաքանչյուրը մոլեկուլային քաշով 35-ից 115 կԴա է: Որոշ դիմերային և տետրամերային ֆերմենտներ կազմված են երկու տեսակի ենթամիավորներից։ Չկա հստակ հարաբերակցություն ֆերմենտի մոլեկուլի չափի կամ նրա ենթամիավորի կառուցվածքի և առանձնահատկությունների միջև:
Ֆերմենտի առանձնահատկությունը որոշվում է նրա ուժեղ կապով tRNA-ի ընդունող ծայրին, DU շրջանին և փոփոխական օղակին: Որոշ ֆերմենտներ, կարծես, չեն ճանաչում հակակոդոնային եռյակը և կատալիզացնում են ամինացետիլացման ռեակցիան նույնիսկ այն դեպքում, երբ հակակոդոնը փոփոխվում է: Այնուամենայնիվ, որոշ ֆերմենտներ ցույց են տալիս նվազեցված ակտիվություն նման ձևափոխված tRNA-ների նկատմամբ և հակակոդոնը փոխարինելիս ավելացնում են սխալ ամինաթթու:
70-90ն | երկրորդական էջ - երեքնուկ | CCA 3» կոնստ բոլոր tRNA |
թիմինի, պսևդուրիդին-փսի, դիգիուրիդին DGU-ի առկայությունը D հանգույցում - պաշտպանություն ռիբոնուկլեազներից: երկարակյաց | tRNA-ի մի շարք առաջնային կառուցվածքներ - 61 + 1 - կոդոնների քանակով + ֆորմիլմեթիոնին tRNA-ն, կատվի հակակոդոնը նույնն է, ինչ մեթիոնինի tRNA-ն: Երրորդական կառուցվածքների բազմազանություն - 20 (ըստ ամինաթթուների քանակի)
Կան երկու տեսակի tRNA կապող մեթիոնին tRNAFMet և tRNAMMet պրոկարիոտներում և tRNAIMet և tRNAMMet էուկարիոտներում: Մեթիոնինը ավելացվում է յուրաքանչյուր tRNA-ին՝ օգտագործելով համապատասխան aminoacyl-tRNA սինթեզը: tRNAFMet-ին և tRNAIMet-ին կցված մեթիոնինը ձևավորվում է մեթիոնիլ-tRNA-տրանսֆորմիլազա ֆերմենտից մինչև Fmet-tRNAFMet: tRNA-ները, որոնք բեռնված են ֆորմիլմեթիոնինով, ճանաչում են մեկնարկային կոդոնը AUG:
Գրականություն:
Ցավոք, մատենագիտություն չկա։
Դասագիրք. Չնայած այն հանգամանքին, որ tRNA-ն շատ ավելի փոքր է, դրա կառուցվածքի, առանձնահատկությունների և գործունեության մասին պատմությունը արժանի է առանձին գլխի:
Այսպիսով, tRNA-ն «ադապտեր» է, որը ճանաչում է գենետիկ կոդի եռատառ հաջորդականությունը մի ծայրում՝ այն համապատասխանեցնելով tRNA-ի մյուս ծայրում ամրագրված միակ համապատասխան ամինաթթվի հետ: Տրանսֆերային ՌՆԹ-ի վերջում, որը դիպչում է սուրհանդակ ՌՆԹ-ին, ձևավորվում են 3 նուկլեոտիդներ. հակակոդոն. Միայն այն դեպքում, եթե հակակոդոնը լրացնում է mRNA տարածաշրջանին, կարող է փոխանցվող ՌՆԹ-ն միանալ դրան: Բայց նույնիսկ այս դեպքում tRNA-ն չի կարող ինքնուրույն միանալ mRNA-ին, նա կարիք ունի ռիբոսոմի օգնությանը, որը նրանց փոխազդեցության վայրն է, ինչպես նաև թարգմանության ակտիվ մասնակցի։ Օրինակ՝ հենց ռիբոսոմն է կապեր ստեղծում tRNA-ի կողմից բերված ամինաթթուների միջև՝ ձևավորելով սպիտակուցային շղթա։
tRNA-ի կառուցվածքային առանձնահատկությունները որոշվում են գենետիկ կոդով, այսինքն՝ սպիտակուցի կառուցման կանոններով՝ ըստ գենի, որը կարդում է փոխանցման ՌՆԹ-ն։ Այս ծածկագիրը գործում է երկրագնդի յուրաքանչյուր կենդանի արարածի մեջ. վիրուսի ստեղծումը գրված է նույն երեք տառանոց կոդոններով, որոնք օգտագործվում են դելֆինի «հավաքման հրահանգները» գրելու համար: Փորձնականորեն հաստատվել է, որ մեկ կենդանի արարածի գեները, որոնք տեղադրված են մյուսի բջիջում, կատարելապես պատճենվում և վերածվում են սպիտակուցների, որոնք չեն տարբերվում տիրոջ բջիջներում վերարտադրվող գեներից: Գենետիկ կոդի միատեսակությունը հիմք է հանդիսանում ինսուլինի և շատ այլ մարդկային ֆերմենտների գաղութների կողմից ձևափոխված E. coli-ի արտադրության համար, որոնք օգտագործվում են որպես դեղամիջոց այն մարդկանց համար, ում մարմինն ի վիճակի չէ կամ անբավարար է արտադրում դրանք: Չնայած մարդկանց և E. coli-ի միջև ակնհայտ տարբերությանը, մարդկային սպիտակուցները հեշտությամբ ստեղծվում են մարդու նախագծերից՝ օգտագործելով E. coli պատճենահանող սարք: Զարմանալի չէ, որ տարբեր արարածների փոխանցման ՌՆԹ-ները շատ քիչ են տարբերվում:

Յուրաքանչյուր կոդոն այս ցուցակից, բացառությամբ երեքի դադարեցնել կոդոնները, որն ազդարարում է թարգմանության ավարտը, պետք է ճանաչվի փոխանցման ՌՆԹ-ի կողմից: Ճանաչումն իրականացվում է սուրհանդակային ՌՆԹ-ին հակակոդոն կցելով, որը կարող է կապվել ցանկից միայն մեկ կոդոնի հետ, ուստի tRNA-ն կարող է ճանաչել միայն մեկ կոդոն: Սա նշանակում է, որ բջիջում կա այդ մոլեկուլների առնվազն 61 տեսակ։ Իրականում, դրանք նույնիսկ ավելի շատ են, քանի որ որոշ իրավիճակներում սուրհանդակային ՌՆԹ-ն կարդալու համար բավարար չէ միայն ճիշտ հակակոդոն ունենալը. այլ պայմաններ են պահանջվում, որոնց համաձայն ստեղծվում է հատուկ, փոփոխված tRNA:
Առաջին հայացքից tRNA-ների նման բազմազանությունը պետք է զգալիորեն բարդացնի թարգմանության գործընթացը. ի վերջո, այս մոլեկուլներից յուրաքանչյուրը կստուգի մատրիցային ՌՆԹ-ի կոդոնը, որը փոխարինում է իրեն ռիբոսոմով, որպեսզի համապատասխանի իր հակակոդոնին. թվում է, թե այդքան անիմաստ մեխանիկական աշխատանք է կատարվում: , այնքան ժամանակ ու էներգիա է վատնել։ Բայց էվոլյուցիայի արդյունքում ձևավորվել են նաև բջջային մեխանիզմներ, որոնք կանխում են այս խնդիրը։ Օրինակ՝ բջջում յուրաքանչյուր տեսակի tRNA-ի քանակը համապատասխանում է նրան, թե որքան հաճախ է այդ տեսակի կրող ամինաթթուն հայտնաբերվում կառուցվող սպիտակուցներում։ Կան ամինաթթուներ, որոնք հազվադեպ են օգտագործվում բջիջի կողմից, և կան այնպիսիք, որոնք հաճախ օգտագործվում են, և եթե դրանք կրող tRNA-ների թիվը նույնը լիներ, դա մեծապես կբարդացներ սպիտակուցների հավաքումը: Հետևաբար, բջջում քիչ են «հազվագյուտ» ամինաթթուները և դրանց համապատասխան tRNA-ները, մինչդեռ հաճախ հանդիպող ամինաթթուները արտադրվում են մեծ քանակությամբ:

TRNA մոլեկուլների նման բազմազանությամբ նրանք բոլորը շատ նման են, հետևաբար, հաշվի առնելով դրանց կառուցվածքը և գործառույթները, մենք հիմնականում կուսումնասիրենք բոլոր տեսակների համար ընդհանուր հատկանիշները: Երբ նայում եք tRNA-ի 3D դասավորությանը, այն կարծես ատոմների խիտ կույտ լինի: Անհավանական է թվում, որ այս խճճված ոլորված մոլեկուլը նուկլեոտիդների երկար շղթայի ծալման արդյունք է, բայց դա այդպես է ձևավորվում:
Հնարավոր է հետևել դրա ձևավորման փուլերին՝ սկսած հենց առաջինից՝ ՌՆԹ պոլիմերազի կողմից նուկլեոտիդային հաջորդականության կազմում՝ համաձայն այս տրանսֆերային ՌՆԹ-ի մասին տեղեկատվություն պարունակող գենի։ Այս նուկլեոտիդների հաջորդականությունը և դրանց թիվը կոչվում է tRNA-ի առաջնային կառուցվածքը. Պարզվում է, որ դա tRNA-ի առաջնային կառուցվածքն է, որը կոդավորված է ՌՆԹ պոլիմերազով կարդացվող գենում։ Ընդհանուր առմամբ, առաջնային կառուցվածքը նույն տեսակի համեմատաբար պարզ մոլեկուլների հաջորդականությունն է, որոնցից կազմված է ավելի բարդ, ծալված պոլիմերային մոլեկուլ։ Օրինակ, սպիտակուցի մոլեկուլի առաջնային կառուցվածքը նրա բաղկացուցիչ ամինաթթուների պարզ հաջորդականությունն է։

Նուկլեոտիդների ցանկացած շղթա բջջում չի կարող լինել բացված վիճակում, պարզապես ձգված է գծով: Նուկլեոտիդների եզրերին կան չափից շատ դրական և բացասական լիցքավորված մասեր, որոնք հեշտությամբ ջրածնային կապեր են ստեղծում միմյանց հետ։ Ինչպես են նույն կապերը ձևավորվում ԴՆԹ-ի երկու մոլեկուլների նուկլեոտիդների միջև՝ դրանք միացնելով կրկնակի պարուրակի մեջ, նկարագրված է, և դուք կարող եք մանրամասնել ջրածնային կապերի մասին: Ջրածնային կապերն ավելի քիչ ամուր են, քան մոլեկուլներում ատոմների միջև կապերը, բայց դրանք բավական են tRNA թելը բարդ ոլորելու և այդ դիրքում պահելու համար: Սկզբում այդ կապերը ձևավորվում են միայն որոշ նուկլեոտիդների միջև՝ tRNA-ն ծալելով երեքնուկի տերևի տեսքով։ Այս սկզբնական ծալման արդյունքը կոչվում է երկրորդական կառուցվածքը tRNA. Ձախ կողմի դիագրամը ցույց է տալիս, որ միայն որոշ նուկլեոտիդներ են կապված ջրածնային կապերով, իսկ մյուսները մնում են չզույգված՝ կազմելով օղակներ և օղակներ։ Տարբեր տեսակի tRNA-ների երկրորդական կառուցվածքի տարբերությունները պայմանավորված են դրանց առաջնային կառուցվածքի տարբերություններով: Սա դրսևորվում է տարբեր երկարությունների «երեքնուկի տերևներով» կամ «ցողունով»՝ նուկլեոտիդների սկզբնական շղթայի տարբեր երկարությունների պատճառով։
Տարբեր tRNA-ների առաջնային կառուցվածքի մեկ այլ տարբերություն այն է, որ միայն որոշ դիրքերում նրանք ունեն նույն նուկլեոտիդները (վերևի գծապատկերում դրանք նշված են իրենց անվան առաջին տառերով), մինչդեռ տարբեր tRNA-ների նուկլեոտիդների մեծ մասը տարբերվում է միմյանցից: Վերոնշյալ սխեման ընդհանուր է բոլոր tRNA-ների համար, ուստի տարբեր նուկլեոտիդները նշվում են թվերով:
tRNA-ի հիմնական ֆունկցիոնալ մասերն են.
=) հակակոդոն, այսինքն՝ նուկլեոտիդային հաջորդականությունը, որը լրացնում է սուրհանդակ ՌՆԹ-ի մեկ կոդոնին, որը գտնվում է հակակոդոնային մազակալ
=) ընդունող վերջորին կարող է կցվել ամինաթթու: Այն գտնվում է հակակոդոնային մազակալի հակառակ կողմում։
Իրականում ոչ մի tRNA-ն նման չէ երկրորդական կառուցվածքի դիագրամում, քանի որ միայն որոշ նուկլեոտիդներ են միացել իրար՝ ձևավորելով այն, իսկ մնացածը մնացել է չզույգված: Երեքնուկի տերևի տարբեր մասերից նուկլեոտիդների միջև ջրածնային կապերի ձևավորման շնորհիվ այն ավելի է ծալվում և դառնում է ավելի բարդ: երրորդական կառուցվածքը L-ի տեսքով: Դուք կարող եք ճշգրիտ հասկանալ, թե ինչպես են երկրորդական կառուցվածքի տարբեր մասերը կորացած՝ ձևավորելով երրորդական կառուցվածքը՝ համապատասխանեցնելով ստորև ներկայացված դրանց գծապատկերների գույները: Հակակոդոնային սանրվածքը, որը նշված է կապույտ և մոխրագույնով, մնում է ներքևում (արժե հիշել, որ այս «ներքևը» պայմանական է. սպիտակուցների թարգմանության սխեմաներում հարմար է պատկերել tRNA-ն այս տարածական կողմնորոշմամբ), իսկ ընդունող ծայրը (դեղին) թեքված է կողքի վրա.

Ահա թե ինչպիսի տեսք ունի tRNA-ն, երբ այն պատրաստ է կցել ամինաթթուն: tRNA-ն ի վիճակի չէ ինքնուրույն միանալ ամինաթթվի հետ, դրա համար անհրաժեշտ է հատուկ ֆերմենտի մասնակցություն. aminoacyl-tRNA սինթետազներ. Բջջում սինթետազների տեսակների թիվը համընկնում է tRNA տեսակների թվի հետ։
Բոլոր տեսակի tRNA-ների ձևի միատեսակությունն անհրաժեշտ է, որպեսզի ռիբոսոմը կարողանա ճանաչել դրանցից որևէ մեկը, հեշտացնել դրանց միացումը mRNA-ի հետ և շարժվել իր ներսում մի տեղից մյուսը: Եթե tRNA-ի տարբեր տեսակներ էականորեն տարբերվեին միմյանցից, դա կդժվարացներ ռիբոսոմի աշխատանքը՝ քննադատորեն նվազեցնելով սպիտակուցի սինթեզի արագությունը: Այսպիսով, բնական ընտրությունը նպատակ ունի tRNA-ները նմանեցնել միմյանց: Բայց միևնույն ժամանակ կա ևս մեկ գործոն, որը պահանջում է նկատելի տարբերությունների առկայություն tRNA-ի տարբեր տեսակների միջև. ի վերջո, անհրաժեշտ է ճանաչել յուրաքանչյուր տեսակ և դրան կցել միակ համապատասխան ամինաթթուն։ Ակնհայտ է, որ այդ տարբերությունները պետք է նկատելի լինեն, բայց ոչ շատ էական, որպեսզի tRNA տեսակների ճանաչման աշխատանքը վերածվի ոսկերչական գործընթացի։ Եվ հենց դա է իրականացվում ամինոացիլ-tRNA սինթետազների միջոցով. նրանցից յուրաքանչյուրը կարող է կապվել 20 ամինաթթուներից միայն մեկին և այն ճշգրիտ կցել tRNA-ի այն տեսակներին, որոնք համապատասխանում են այս ամինաթթունին: Գենետիկ կոդով աղյուսակից երևում է, որ յուրաքանչյուր ամինաթթու կոդավորված է մի քանի նուկլեոտիդային հաջորդականությամբ, հետևաբար, օրինակ, բոլոր չորս tRNA-ները հակակոդոններով CGA, CGG, CGU և CGC կճանաչվեն նույն սինթետազով, որը կցում է ալանինը: նրանց. Մեկ սինթետազով մշակված այդպիսի tRNA-ները կոչվում են կապված.
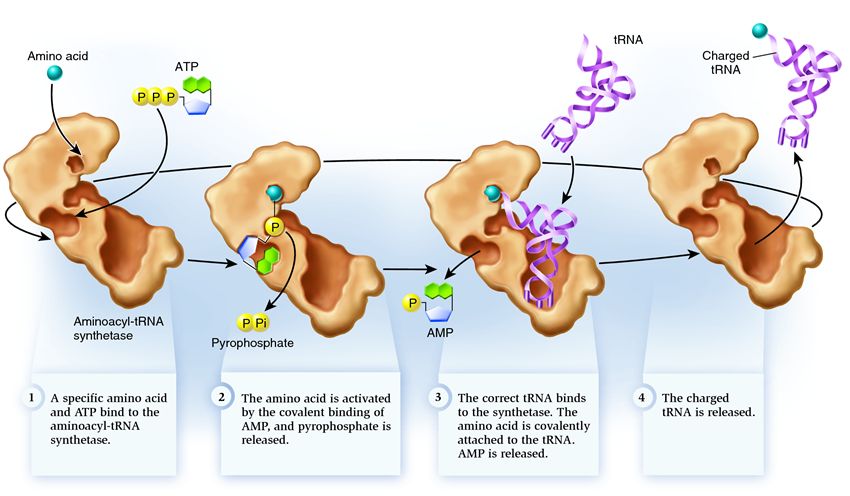
Սինթետազը պատկանում է մի խումբ ֆերմենտների, որոնց գործառույթն է կապել առանձին գոյություն ունեցող մոլեկուլներին և միավորել դրանք մեկում.
1 . սինթետազը միացնում է ամինաթթուն և ATP մոլեկուլը: Երկու ֆոսֆատ խմբեր անջատվում են ATP-ից՝ ազատելով էներգիան, որն անհրաժեշտ է հետևյալ գործողությունների համար. Քանդված մոլեկուլից մնացած ադենոզին մոնոֆոսֆատը (AMP) կպչում է ամինաթթունին՝ նախապատրաստելով այն ընդունող վարսահարդարիչի հետ միացմանը։
2 . սինթետազն իրեն կցում է այս ամինաթթուն համապատասխանող tRNA-ներից մեկը:

Այս փուլում ստուգվում է փոխանցման ՌՆԹ-ի համապատասխանությունը սինթետազին։ Կան ճանաչման մի քանի եղանակներ, և յուրաքանչյուր սինթետազ ունի դրանց եզակի համադրություն: Առնվազն մեկ հակակոդոն նուկլեոտիդ ներգրավված է սինթետազի և tRNA-ի փոխազդեցության մեջ: Ստուգման կարիք ունի նաև ընդունող մազակալը. որոշվում է դրա վրա հատուկ նուկլեոտիդների առկայությունը, որոնք բնորոշ են համապատասխան tRNA-ներին, որոնք համապատասխանում են ցանկալի ամինաթթուն: tRNA-ի այլ մասերի նուկլեոտիդները նույնպես կարող են մասնակցել համընկնումին՝ կապված որոշակի սինթետազային տեղամասերի հետ: Սխալ tRNA-ն կարող է ինչ-որ կերպ համընկնել ցանկալիին, սակայն թերի համապատասխանության պատճառով այն դանդաղ և թույլ կմիանա սինթետազին՝ հեշտությամբ ընկնելով: Իսկ ճիշտ tRNA-ն արագ և ամուր կկպչի սինթետազին, ինչի արդյունքում փոխվում է սինթետազի կառուցվածքը՝ սկսելով գործընթացը. ամինոացիլացիա , այսինքն՝ ամինաթթվի կցումը tRNA-ին։

3 . aminoacylation բաղկացած է փոխարինել AMP մոլեկուլը կցված է amino թթու է tRNA մոլեկուլ. Այս փոխարինումից հետո AMP-ը թողնում է սինթետազը և tRNA-ն պահվում է վերջին ամինաթթուների ստուգման համար: Եթե կցված ամինաթթուն սխալ ճանաչվի, այն կկտրվի tRNA-ից, սինթետազում ամինաթթվի տեղը դատարկ կլինի, և մեկ այլ մոլեկուլ կարող է միանալ այնտեղ։ Նոր ամինաթթուն կանցնի ATP-ի և tRNA-ի հետ կապի փուլերը, ինչպես նաև կփորձարկվի։ Եթե սխալներ թույլ չեն տրվել, ամինաթթուով լիցքավորված tRNA-ն ազատվում է. այն պատրաստ է իր դերը կատարել սպիտակուցների թարգմանության մեջ: Իսկ սինթետազը պատրաստ է կցել նոր ամինաթթուներ և tRNA-ներ, և ցիկլը նորից կսկսվի։
Շատ բան կախված է aminoacyl-tRNA սինթետազի ճիշտ աշխատանքից. եթե այս փուլում խափանում է տեղի ունենում, ապա tRNA-ին կկցվի սխալ ամինաթթու: Եվ այն կներկառուցվի ռիբոսոմի վրա աճող սպիտակուցի մեջ, քանի որ tRNA-ն և ռիբոսոմը չունեն կոդոնի և ամինաթթվի համապատասխանությունը ստուգելու գործառույթ։ Սխալի հետևանքները կարող են լինել աննշան կամ աղետալի, և բնական ընտրության միջոցով ֆերմենտներով արարածները, որոնք նման ստուգումների գործառույթ չունեն, փոխարինվել են ավելի հարմարվողներով՝ ամինաթթուների և tRNA-ի միջև համապատասխանեցնելու տարբեր տարբերակներով: Հետևաբար, ժամանակակից բջիջներում սինթետազը 50 հազարից միջինում մեկ դեպքում միավորվում է սխալ ամինաթթվի հետ, իսկ սխալ tRNA-ի հետ՝ 100 հազար կցորդներից միայն մեկ անգամ։

Որոշ ամինաթթուներ միմյանցից տարբերվում են ընդամենը մի քանի ատոմով։ Եթե նայեք դրանց սխեմաներին, ապա ակնհայտ է դառնում, որ արգինինը ալանինի հետ շփոթելու հավանականությունը շատ ավելի քիչ է, քան իզոլեյցինը լեյցինի կամ վալինի հետ շփոթելու հավանականությունը: Հետևաբար, յուրաքանչյուր սինթետազ, որը կապվում է միմյանց նման ամինաթթուներից մեկին, ունի լրացուցիչ ստուգման մեխանիզմներ: Ահա այսպիսի հարմարվողականության օրինակ isoleucine-tRNA synthetase-ում.

Յուրաքանչյուր սինթազ ունի սինթետիկ կենտրոնորի մեջ ամինաթթուն կցված է tRNA-ին: Սինթետազի կողմից գրավված tRNA-ի ընդունիչ մազակալը գնում է այնտեղ, ինչպես նաև ամինաթթուն, որը պատրաստ է արձագանքել դրա հետ: Որոշ սինթետազների աշխատանքն ավարտվում է ամինաթթվի և tRNA-ի միացումից անմիջապես հետո։ Բայց Ile-tRNA սինթետազը սխալվելու մեծ հավանականություն ունի՝ այլ իզոլեյցինանման ամինաթթուների առկայության պատճառով: Հետեւաբար, նա նույնպես ունի ուղղիչ կենտրոնանունից պարզ է դառնում, թե ինչ դեր է խաղում tRNA-ի և ամինաթթուների միացման գործընթացում։ Աջ նկարը ցույց է տալիս, որ tRNA ընդունիչ սանրվածքի վերջի դիրքը Ile-tRNA սինթետազի սինթետիկ կենտրոնում այս մազակալին տալիս է անբնական թեքում: Այնուամենայնիվ, սինթետազը tRNA-ն պահում է այս դիրքում, մինչև ամինաթթուն կցվի դրան: Այս միացումից հետո սինթետիկ կենտրոնում ակցեպտորի մազակալը գտնելու անհրաժեշտությունը սպառվում է, և tRNA-ն ուղղվում է՝ իր ծայրը դրան կցված ամինաթթուով հասցնելով ուղղման կենտրոն:

Իհարկե, սինթետիկ կենտրոնը նույնպես դեր է խաղում սինթետազի համար ոչ պիտանի ամինաթթուների զննման գործում: Դրա մեջ մտնելու համար մոլեկուլը պետք է համապատասխանի մի շարք պայմանների, այդ թվում՝ ունենալ համապատասխան չափ: Չնայած այն հանգամանքին, որ լեյցինը և իզոլեյցինը պարունակում են նույն թվով ատոմներ, տարածական կառուցվածքի տարբերությունների պատճառով լեյցինը ավելի մեծ է: Հետեւաբար, այն չի կարող ներթափանցել սինթետիկ կենտրոն, որի չափը համապատասխանում է ավելի կոմպակտ իզոլեյցինին, և պարզապես ցատկում է Ile-tRNA սինթետազից:
Բայց վալինը, որն այս երեք մոլեկուլներից ամենափոքրն է, որն ունի նմանատիպ ատոմային կառուցվածք, հեշտությամբ սինթետիկ կենտրոնում զբաղեցնում է իզոլեյցինի տեղը, իսկ սինթետազը այն միացնում է tRNA-ին։ Հենց այս դեպքում է, որ գործում է սինթետազի ուղղիչ կենտրոնը։ Եթե ուղղիչ ընդունիչ սանրվածքը ճիշտ լիցքավորված է և կրում է իզոլեյցին, ապա այն չի կարող սեղմվել ուղղիչ կենտրոնի ներսում. այն պարզապես շատ փոքր է այս մոլեկուլի համար: Այսպիսով, ուղղված tRNA-ն այլևս ոչնչով չի պահվում, և այն անջատվում է սինթետազից։ Բայց եթե վալինը կցվում է tRNA-ին, այն սահում է ուղղիչ կենտրոն, դրանով իսկ tRNA-ն միացված է մնում սինթետազում: Ներսում tRNA-ի չափազանց երկար մնալը սխալ ազդանշան է սինթետազի համար՝ փոխելով դրա տարածական կոնֆիգուրացիան: Որպես արդյունք:
=) վալինը անջատվում է tRNA-ից և հեռացվում սինթետազից
=) ընդունող սանրվածքը վերադառնում է սինթետիկ տեղ՝ սպասելով ամինաթթուին միանալուն
=) սինթետազը կապվում է նոր ամինաթթվի հետ, դրանով «լիցքավորում է» tRNA և նորից ստուգում, թե արդյոք դրա համար օգտագործվել է իզոլեյցին:
Նմանատիպ կրկնակի ճանաչման մեխանիզմ օգտագործվում է այլ սինթետազների կողմից:
ԴՆԹ-ի ֆիզիկական և քիմիական հատկությունները
Ջրածնային կապերը խզող տարբեր գործոններ (ջերմաստիճանի բարձրացում 80 C-ից բարձր, pH-ի և իոնային ուժի փոփոխություն, միզանյութի ազդեցություն և այլն) առաջացնում են ԴՆԹ-ի դենատուրացիա, այսինքն. ԴՆԹ-ի շղթաների տարածական դասավորության փոփոխություն՝ առանց կովալենտային կապերի խզման: ԴՆԹ-ի կրկնակի պարույրը դենատուրացիայի ժամանակ ամբողջությամբ կամ մասամբ բաժանվում է իր բաղադրիչ շղթաների։ ԴՆԹ-ի դենատուրացիան ուղեկցվում է պուրինային և պիրիմիդինային հիմքերի ուլտրամանուշակագույն ճառագայթման շրջանում օպտիկական կլանման ավելացմամբ: Այս երեւույթը կոչվում է հիպերքրոմային ազդեցություն . Դենատուրացիան նաև նվազեցնում է բնական ԴՆԹ լուծույթներին բնորոշ բարձր մածուցիկությունը: Երբ վերականգնվում է սկզբնական երկշղթա ԴՆԹ-ի կառուցվածքը, վերածնման արդյունքում 260 նմ-ում ազոտային հիմքերի կողմից կլանումը նվազում է դրանց «պաշտպանվածության» պատճառով։ Այս երեւույթը կոչվում է հիպոքրոմային ազդեցություն .
Յուրաքանչյուր ԴՆԹ-ի «փաթաթումը» իր բաղադրիչ շղթաների մեջ իրականացվում է որոշակի ջերմաստիճանի միջակայքում։ Այս միջակայքի միջնակետը կոչվում է հալման կետ: ԴՆԹ-ի հալման ջերմաստիճանը ստանդարտ պայմաններում (որոշակի pH և իոնային ուժ) կախված է ազոտային հիմքերի հարաբերակցությունից։ Երեք ջրածնային կապ պարունակող G-C զույգերը ավելի ամուր են, հետևաբար, որքան բարձր է G-C զույգերի պարունակությունը ԴՆԹ-ում, այնքան բարձր է հալման կետը:
ԴՆԹ-ի գործառույթները. ԴՆԹ-ի մոլեկուլներում նուկլեոտիդների հաջորդականության մեջ գենետիկական տեղեկատվությունը կոդավորված է: ԴՆԹ-ի հիմնական գործառույթներն են, առաջին հերթին, ապահովել իր վերարտադրությունը մի շարք բջիջների սերունդների և օրգանիզմների սերունդների մեջ, և երկրորդը, ապահովել սպիտակուցների սինթեզը: Այս գործառույթները պայմանավորված են նրանով, որ ԴՆԹ-ի մոլեկուլները ծառայում են որպես մատրիցա առաջին դեպքում վերարտադրության համար, այսինքն. տեղեկատվության պատճենումը դուստր ԴՆԹ-ի մոլեկուլներում, երկրորդում՝ տառադարձման համար, այսինքն. տեղեկատվությունը ՌՆԹ-ի կառուցվածքում վերակոդավորելու համար:
Բրինձ. 5 Հալման կոր (ԴՆԹ-ի դենատուրացիա)
Դենատուրացիայի ժամանակ առանձնացված ԴՆԹ-ի կոմպլեմենտար շղթաները, որոշակի պայմաններում, կարող են նորից միանալ կրկնակի պարույրի: Այս գործընթացը կոչվում է RENATURATION: Եթե denaturation-ը ամբողջությամբ տեղի չի ունեցել, և առնվազն մի քանի հիմքեր չեն կորցրել ջրածնային կապերի փոխազդեցությունը, ապա վերածումը տեղի է ունենում շատ արագ:
Բջիջների ցիտոպլազմը պարունակում է ՌՆԹ-ի երեք հիմնական ֆունկցիոնալ տեսակ. Սրանք սուրհանդակային ՌՆԹ-ներ են՝ mRNA-ներ, որոնք գործում են որպես սպիտակուցների սինթեզի ձևանմուշներ, ռիբոսոմային ՌՆԹ-ներ՝ rRNAs, որոնք գործում են որպես ռիբոսոմների կառուցվածքային բաղադրիչներ, և փոխանցում են ՌՆԹ-ներ՝ tRNA-ներ, որոնք ներգրավված են mRNA տեղեկատվության թարգմանության (թարգմանման) մեջ սպիտակուցի ամինաթթուների հաջորդականության մեջ:
Աղյուսակ 2-ը ցույց է տալիս ԴՆԹ-ի և ՌՆԹ-ի տարբերությունները կառուցվածքի, բջջում տեղայնացման և գործառույթների առումով:
Աղյուսակ 2 ԴՆԹ-ի և ՌՆԹ-ի տարբերությունները
Տրանսֆերային ՌՆԹ, tRNA-ռիբոնուկլեինաթթու, որի գործառույթը AA-ն սպիտակուցի սինթեզի վայր տեղափոխելն է: Այն ունի 73-ից 93 նուկլեոտիդների տիպիկ երկարություն և մոտ 5 նմ չափ: tRNA-ները նույնպես անմիջականորեն մասնակցում են պոլիպեպտիդային շղթայի աճին, միանալով ամինաթթվի հետ բարդույթի մեջ՝ միանալով mRNA կոդոնին և ապահովելով նոր պեպտիդային կապի ձևավորման համար անհրաժեշտ համալիրի կոնֆորմացիան: Յուրաքանչյուր ամինաթթու ունի իր սեփական tRNA: tRNA-ն միաշղթա ՌՆԹ է, բայց իր ֆունկցիոնալ տեսքով ունի երեքնուկային կոնֆորմացիա։ AA-ն կովալենտորեն կցվում է մոլեկուլի 3" ծայրին, օգտագործելով aminoacyl-tRNA սինթետազ ֆերմենտը, որը հատուկ է tRNA-ի յուրաքանչյուր տեսակի համար: C տեղում կա AA-te-ին համապատասխան հակակոդոն: tRNA-ները սինթեզվում են սովորական ՌՆԹ պոլիմերազի դեպքում: պրոկարիոտների և ՌՆԹ պոլիմերազ III-ի կողմից էուկարիոտների դեպքում. tRNA գեների տրանսկրիպտները ենթարկվում են բազմաստիճան վերամշակման, ինչը հանգեցնում է tRNA-ին բնորոշ տարածական կառուցվածքի ձևավորմանը:
tRNA մշակումը ներառում է 5 հիմնական քայլ.
5" առաջատար նուկլեոտիդային հաջորդականության հեռացում;
3'-տերմինալ հաջորդականության հեռացում;
ավելացնելով CCA հաջորդականություն 3 դյույմանոց վերջում;
ինտրոնների հեռացում (էուկարիոտներում և արխեայում);
առանձին նուկլեոտիդների փոփոխություններ.
tRNA-ի փոխադրումն իրականացվում է Ran-ից կախված ճանապարհով՝ արտահանման t-ի տրանսպորտային գործոնի մասնակցությամբ, որը ճանաչում է հասուն tRNA-ի բնորոշ երկրորդական և երրորդական str-ru-ն՝ կարճ երկշղթա հատվածներ և ճիշտ մշակված 5 «- և 3»: ավարտվում է. Այս մեխանիզմը ապահովում է, որ միջուկից արտահանվեն միայն հասուն tRNA-ները:
62. Թարգմանություն - mRNA կոդոնի ճանաչում
Թարգմանությունը սպիտակուցի սինթեզ է, որն իրականացվում է ամինաթթուների ռիբոսոմների կողմից mRNA (կամ և RNA) ձևանմուշի վրա: Թարգմանության գործընթացի բաղկացուցիչ տարրերը՝ ամինաթթուներ, tRNA, ռիբոսոմներ, mRNA, ֆերմենտներ tRNA-ի ամինոացիլացման համար, սպիտակուցի թարգմանության գործոններ (սկսման, երկարացման, ավարտի սպիտակուցային գործոններ՝ թարգմանչական գործընթացների համար անհրաժեշտ հատուկ արտառիբոսոմային սպիտակուցներ), ATP և GTP էներգիայի աղբյուրներ։ , մագնեզիումի իոններ (կայունացնում են ռիբոսոմի կառուցվածքը)։ Սպիտակուցների սինթեզում ներգրավված են 20 ամինաթթուներ: Որպեսզի ամինաթթուն «ճանաչի» իր տեղը ապագա պոլիպեպտիդային շղթայում, այն պետք է կապվի փոխանցող ՌՆԹ-ին (tRNA), որը կատարում է ադապտերային ֆունկցիա: tRNA-ն, որը միանում է ամինաթթուն, այնուհետև ճանաչում է mRNA-ի համապատասխան կոդոնը: mRNA կոդոնի ճանաչում.
Կոդոն-հակակոդոն փոխազդեցությունը հիմնված է փոխլրացման և հակազուգահեռացման սկզբունքների վրա.
3'----C - G-A*------5' tRNA հակակոդոն
5'-----G-C-Y*------3' mRNA կոդոն
Տատանումների վարկածն առաջարկվել է Ֆ. Քրիքի կողմից.
mRNA կոդոնի 3' բազան ունի ոչ խիստ զուգավորում tRNA հակակոդոնի 5' հիմքի հետ. օրինակ, Y (mRNA) կարող է փոխազդել A և G (tRNA) հետ:
Որոշ tRNA-ներ կարող են զուգակցվել մեկից ավելի կոդոնների հետ:
63. Թարգմանության գործընթացի բաղկացուցիչ տարրերի բնութագրերը.Թարգմանությունը (translatio-translation) ամինաթթուներից սպիտակուցի սինթեզի գործընթացն է տեղեկատվական (մատրիցային) ՌՆԹ-ի (mRNA, mRNA) մատրիցով, որն իրականացվում է ռիբոսոմի կողմից:
Սպիտակուցների սինթեզը բջիջների կյանքի հիմքն է: Այս գործընթացն իրականացնելու համար բոլոր օրգանիզմների բջիջներում կան հատուկ օրգանելներ. ռիբոսոմներ- ռիբոնուկլեոպրոտեինային համալիրներ, որոնք կառուցված են 2 ենթամիավորներից՝ մեծ և փոքր: Ռիբոսոմների ֆունկցիան եռատառ (երեք նուկլեոտիդ) ճանաչելն է։ կոդոններ mRNA, համեմատելով դրանք համապատասխան tRNA հակակոդոնների հետ ամինաթթուներ, և այս ամինաթթուների ավելացումը աճող սպիտակուցային շղթային: Շարժվելով mRNA մոլեկուլով՝ ռիբոսոմը սինթեզում է սպիտակուց՝ mRNA մոլեկուլում պարունակվող տեղեկատվությանը համապատասխան։
Բջջում AK-t-ի ճանաչման համար կան հատուկ «ադապտերներ», փոխանցել ՌՆԹ մոլեկուլները(tRNA): Երեքնուկի տերևի նման մոլեկուլները ունեն տեղ (հակիկոդոն), որը լրացնում է mRNA կոդոնին, ինչպես նաև մեկ այլ տեղ, որին կցված է այդ կոդոնին համապատասխան ամինաթթուն։ Ամինաթթուների կցումը tRNA-ին իրականացվում է էներգիայից կախված ռեակցիայի միջոցով aminoacyl-tRNA սինթետազների ֆերմենտների միջոցով, և արդյունքում առաջացող մոլեկուլը կոչվում է aminoacyl-tRNA: Այսպիսով, թարգմանության առանձնահատկությունը որոշվում է mRNA կոդոնի և tRNA հակակոդոնի փոխազդեցությամբ, ինչպես նաև ամինացիլ-tRNA սինթետազների յուրահատկությամբ, որոնք ամինաթթուները խստորեն կցում են իրենց համապատասխան tRNA-ներին (օրինակ, GGU կոդոնը կհամապատասխանի tRNA, որը պարունակում է CCA հակակոդոն և միայն AK գլիցին):
պրոկարիոտիկ ռիբոսոմ
5S և 23S rRNA 16S rRNA
34 squirrels 21 squirrels
Պրոկարիոտների ռիբոսոմներն ունեն 70S նստվածքային հաստատուն, ինչի պատճառով էլ կոչվում են 70S մասնիկներ։ Դրանք կառուցված են երկու տարբեր ստորաբաժանումներից՝ 30S և 50S ստորաբաժանումներից: Յուրաքանչյուր ենթամիավոր իրենից ներկայացնում է rRNA և ռիբոսոմային սպիտակուցների համալիր:
30S մասնիկը պարունակում է մեկ 16S rRNA մոլեկուլ և շատ դեպքերում մեկ սպիտակուցի մոլեկուլ՝ ավելի քան 20 տեսակներից (21): 50S ենթամիավորը բաղկացած է երկու rRNA մոլեկուլներից (23S և 5S): Այն բաղկացած է ավելի քան 30 տարբեր սպիտակուցներից (34), որոնք նույնպես, որպես կանոն, ներկայացված են մեկ օրինակով։ Ռիբոսոմային սպիտակուցների մեծ մասը կատարում է կառուցվածքային ֆունկցիա։
էուկարիոտիկ ռիբոսոմ
5S; 5,8S և 28S rRNA 18S rRNA
առնվազն 50 սպիտակուց առնվազն 33 սպիտակուց
Ռիբոսոմը բաղկացած է մեծ և փոքր ենթամիավորներից։ Յուրաքանչյուր ենթամիավորի կառուցվածքի հիմքը բարդ ծալված rRNA-ն է։ Ռիբոսոմի սպիտակուցները կցվել են rRNA փայտամածին:
Ամբողջական էուկարիոտիկ ռիբոսոմի նստվածքային գործակիցը կազմում է մոտ 80 Svedberg միավոր (80S), իսկ նրա ենթամասնիկների նստվածքային գործակիցը՝ 40S և 60S։
Ավելի փոքր 40S ենթամիավորը բաղկացած է մեկ 18S rRNA մոլեկուլից և 30-40 սպիտակուցի մոլեկուլից։ 60S մեծ ենթաբաժինը պարունակում է երեք տեսակի rRNA՝ 5S, 5.8S և 28S նստվածքային գործակիցներով և 40-50 սպիտակուցներ (օրինակ՝ առնետի հեպատոցիտների ռիբոսոմները ներառում են 49 սպիտակուցներ)։
Ռիբոսոմների ֆունկցիոնալ շրջաններ
P - պեպտիդիլային կայք պեպտիդիլ tRNA-ի համար
A - aminoacyl կայքը aminoacyl tRNA
E - կայք ռիբոսոմից tRNA-ի ազատման համար
Ռիբոսոմը պարունակում է tRNA-ի հետ փոխազդեցության 2 ֆունկցիոնալ տեղամաս՝ ամինացիլ (ընդունող) և պեպտիդիլ (դոնոր)։ Ամինոացիլ-tRNA-ն մտնում է ռիբոսոմի ընդունող տեղ և փոխազդում է կոդոնի և հակակոդոնային եռյակների միջև ջրածնային կապեր ձևավորելու համար: Ջրածնային կապերի ձևավորումից հետո համակարգը առաջ է մղում 1 կոդոն և հայտնվում դոնոր տեղում: Միաժամանակ ազատված ընդունող տեղամասում հայտնվում է նոր կոդոն, որի վրա կցվում է համապատասխան ամինոացիլ-t-ՌՆԹ-ն։
Ռիբոսոմներ՝ կառուցվածք, ֆունկցիա
Ռիբոսոմները սպիտակուցների կենսասինթեզի ցիտոպլազմային կենտրոններն են։ Կազմված են խոշոր և փոքր ստորաբաժանումներից, որոնք տարբերվում են նստվածքի գործակիցներով (ցենտրիֆուգման ժամանակ նստվածքի արագությունը), արտահայտված Svedberg - Ս. միավորներով։
Ռիբոսոմները առկա են ինչպես էուկարիոտ, այնպես էլ պրոկարիոտ բջիջներում, քանի որ դրանք կատարում են կարևոր գործառույթ սպիտակուցի կենսասինթեզ.Յուրաքանչյուր բջիջ պարունակում է տասնյակ, հարյուր հազարավոր (մինչև մի քանի միլիոն) այս փոքր կլորացված օրգանելներ։ Այն կլորացված ռիբոնուկլեոպրոտեինային մասնիկ է։ Դրա տրամագիծը 20-30 նմ է։ Ռիբոսոմը բաղկացած է մեծ և փոքր ենթամիավորներից, որոնք տարբերվում են նստվածքի գործակիցներով (ցենտրիֆուգման ժամանակ նստվածքի արագությունը), արտահայտված Svedberg միավորներով - S։ Ռիբոսոմների խմբի համալիրը, որը միավորված է մեկ mRNA մոլեկուլով, ինչպես ուլունքների շարանը կոչվում է. պոլիսոմ. Այս կառույցները կա՛մ ազատորեն տեղակայված են ցիտոպլազմայում, կա՛մ կցվում են հատիկավոր ԷՀ-ի թաղանթներին (երկու դեպքում էլ դրանց վրա ակտիվորեն ընթանում է սպիտակուցի սինթեզը)։
Հատիկավոր ER-ի պոլիսոմները ձևավորում են սպիտակուցներ, որոնք արտազատվում են բջջից և օգտագործվում են ամբողջ օրգանիզմի կարիքների համար (օրինակ՝ մարսողական ֆերմենտներ, մարդու կրծքի կաթի սպիտակուցներ): Բացի այդ, ռիբոսոմները առկա են միտոքոնդրիալ թաղանթների ներքին մակերեսին, որտեղ նրանք նույնպես ակտիվ մասնակցություն են ունենում սպիտակուցի մոլեկուլների սինթեզում։
Տրանսֆերային ՌՆԹ-ն (tRNA) կարևոր դեր է խաղում բջջի կողմից ժառանգական տեղեկատվության օգտագործման գործընթացում: Հաղորդելով անհրաժեշտ ամինաթթուները պեպտիդային շղթաների հավաքման վայր՝ tRNA-ն հանդես է գալիս որպես թարգմանական միջնորդ։
tRNA մոլեկուլները պոլինուկլեոտիդային շղթաներ են, որոնք սինթեզվում են հատուկ ԴՆԹ հաջորդականությունների վրա: Դրանք կազմված են համեմատաբար փոքր թվով նուկլեոտիդներից -75-95։ Հիմքերի կոմպլեմենտար միացման արդյունքում, որոնք տեղակայված են tRNA պոլինուկլեոտիդային շղթայի տարբեր մասերում, այն ձեռք է բերում տեսքով երեքնուկի տերևի նմանվող կառուցվածք (նկ. 3.26):
Բրինձ. 3.26. Տիպիկ tRNA մոլեկուլի կառուցվածքը.
Այն ունի չորս հիմնական մասեր, որոնք կատարում են տարբեր գործառույթներ: ընդունող«Ցողունը» ձևավորվում է tRNA-ի երկու փոխլրացնող տերմինալ մասերով։ Այն բաղկացած է յոթ բազային զույգերից։ Այս ցողունի 3' ծայրը որոշ չափով ավելի երկար է և կազմում է միաշղթա շրջան, որն ավարտվում է CCA հաջորդականությամբ՝ ազատ OH խմբով: Այս ծայրին կցվում է տեղափոխվող ամինաթթու: Մնացած երեք ճյուղերը փոխլրացնող զույգ նուկլեոտիդային հաջորդականություններ են, որոնք ավարտվում են չզույգված հատվածներով, որոնք կազմում են օղակներ։ Այս ճյուղերի միջին մասը՝ հակակոդոնը, բաղկացած է հինգ զույգ նուկլեոտիդներից և պարունակում է հակակոդոն իր օղակի կենտրոնում։ Հակոդոնը երեք նուկլեոտիդ է, որոնք լրացնում են mRNA կոդոնին, որը կոդավորում է այս tRNA-ի կողմից պեպտիդների սինթեզի վայր տեղափոխվող ամինաթթուն:
Ընդունիչի և հակակոդոնային ճյուղերի միջև կան երկու կողային ճյուղեր: Իրենց օղակներում դրանք պարունակում են փոփոխված հիմքեր՝ դիհիդրոուրիդին (D-loop) և TψC եռյակը, որտեղ \y-ը կեղծավոր է (T^C-loop):
Aiticodone և T^C ճյուղերի միջև կա լրացուցիչ հանգույց, որը ներառում է 3-5-ից մինչև 13-21 նուկլեոտիդներ:
Ընդհանուր առմամբ, tRNA-ի տարբեր տեսակները բնութագրվում են նուկլեոտիդային հաջորդականության որոշակի կայունությամբ, որն առավել հաճախ բաղկացած է 76 նուկլեոտիդից։ Դրանց թվի փոփոխությունը հիմնականում պայմանավորված է լրացուցիչ օղակում նուկլեոտիդների քանակի փոփոխությամբ։ Կոմպլեմենտար շրջանները, որոնք աջակցում են tRNA կառուցվածքին, սովորաբար պահպանվում են: tRNA-ի առաջնային կառուցվածքը, որը որոշվում է նուկլեոտիդների հաջորդականությամբ, կազմում է tRNA-ի երկրորդական կառուցվածքը, որն ունի երեքնուկի տերեւի տեսք։ Իր հերթին, երկրորդական կառուցվածքը առաջացնում է եռաչափ երրորդական կառուցվածք, որը բնութագրվում է երկու ուղղահայաց կրկնակի խխունջների ձևավորմամբ (նկ. 3.27): Դրանցից մեկը ձևավորվում է ակցեպտորով և TψC ճյուղերով, մյուսը՝ հակակոդոններով և D ճյուղերով։
Կրկնակի պարույրներից մեկի վերջում տեղափոխվող ամինաթթուն է, մյուսի վերջում՝ հակակոդոնը։ Այս տարածքները միմյանցից ամենահեռավորն են։ tRNA-ի երրորդային կառուցվածքի կայունությունը պահպանվում է պոլինուկլեոտիդային շղթայի տարբեր մասերում տեղակայված, բայց երրորդական կառուցվածքում տարածականորեն փակ պոլինուկլեոտիդային շղթայի հիմքերի միջև լրացուցիչ ջրածնային կապերի առաջացման պատճառով։
tRNA-ների տարբեր տեսակներ ունեն նմանատիպ երրորդական կառուցվածք, թեև որոշ տատանումներով:
Բրինձ. 3.27. tRNA-ի տարածական կազմակերպում.
I - tRNA-ի երկրորդական կառուցվածքը «երեքնուկի տերևի» տեսքով, որը որոշվում է նրա առաջնային կառուցվածքով (շղթայում նուկլեոտիդների հաջորդականությունը);
II - tRNA-ի երրորդական կառուցվածքի երկչափ պրոյեկցիա;
III - tRNA մոլեկուլի դասավորությունը տարածության մեջ
ՀԱՎԵԼՎԱԾ (եթե ինչ-որ մեկը դա չի հասկանում)
Կայծակ ատամներ - նուկլեոտիդներ (Ադենին-Թիմին / Ուրասիլ /, Գուանին-Ցիտազին): Ամբողջ կայծակը ԴՆԹ է:

ԴՆԹ-ից տեղեկատվություն փոխանցելու համար անհրաժեշտ է կոտրել 2 թել: A-T-ի և G-C-ի միջև կապը ջրածին է, հետևաբար այն հեշտությամբ կոտրվում է Helicase ֆերմենտի կողմից.

Հանգույցների ձևավորումը կանխելու համար (Որպես օրինակ, ես պտտեցի սրբիչը).



Տոպոիզոմերազը կրկնօրինակման սկզբում կտրում է ԴՆԹ-ի մեկ շղթան, որպեսզի շղթան չոլորվի:

Երբ մի թելը ազատ է, երկրորդը կարող է հեշտությամբ պտտվել իր առանցքի շուրջ՝ դրանով իսկ ազատելով լարվածությունը «թափվելու» ժամանակ։ Հանգույցները չեն հայտնվում, էներգիան խնայվում է:
Այնուհետև անհրաժեշտ է ՌՆԹ այբբենարան՝ ՌՆԹ-ի հավաքումը սկսելու համար: Սպիտակուցը, որը հավաքում է mRNA-ն, չի կարող պարզապես հավաքել առաջին նուկլեոտիդը, դրա մեկնարկի համար անհրաժեշտ է ՌՆԹ-ի մի կտոր (այնտեղ մանրամասն գրված է, հետո կգրեմ): Այս կտորը կոչվում է ՌՆԹ այբբենարան: Եվ այս սպիտակուցն արդեն կցում է առաջին նուկլեոտիդը։


Նուկլեինաթթուների կառուցվածքը նկարագրելիս հաշվի են առնվում մակրոմոլեկուլների կազմակերպման տարբեր մակարդակներ՝ առաջնային և երկրորդային կառուցվածք։
Նուկլեինաթթուների առաջնային կառուցվածքը նուկլեոտիդային կազմն է և պոլիմերային շղթայում նուկլեոտիդային միավորների որոշակի հաջորդականություն։
ՌՆԹ-ի երկրորդական կառուցվածքը. Ռիբոնուկլեինաթթվի մոլեկուլը կառուցված է մեկ պոլինուկլեոտիդային շղթայից։
ՌՆԹ-ի երկրորդական կառուցվածքը
ՌՆԹ-ի շղթայի առանձին հատվածները կազմում են պարուրաձև օղակներ՝ «մազակալներ», որոնք պայմանավորված են A-U և G-C կոմպլեմենտար ազոտային հիմքերի միջև ջրածնային կապերով: Նման պտուտակավոր կառուցվածքներում ՌՆԹ-ի շղթայի հատվածները հակազուգահեռ են, բայց ոչ միշտ՝ ամբողջությամբ փոխլրացնող, դրանք պարունակում են չզույգված նուկլեոտիդային մնացորդներ կամ նույնիսկ միաշղթա օղակներ, որոնք չեն տեղավորվում կրկնակի պարույրի մեջ: Պարուրաձև շրջանների առկայությունը բնորոշ է ՌՆԹ-ի բոլոր տեսակներին:
ՌՆԹ-ի հիմնական դերը սպիտակուցի կենսասինթեզի անմիջական մասնակցությունն է։
Հայտնի են բջջային ՌՆԹ-ի երեք տեսակ, որոնք տարբերվում են բջջում իրենց տեղակայմամբ, կազմով, չափսերով և հատկություններով, որոնք որոշում են նրանց հատուկ դերը սպիտակուցի մակրոմոլեկուլների ձևավորման գործում.
- տեղեկատվական (մատրիցային) ՌՆԹ-ները սպիտակուցի կառուցվածքի մասին ԴՆԹ-ում կոդավորված տեղեկատվություն են փոխանցում բջջի միջուկից դեպի ռիբոսոմներ, որտեղ իրականացվում է սպիտակուցի սինթեզ. Բոլոր mRNA-ների առաջնային կառուցվածքը, անկախ դրանց կոդավորման հաջորդականության եզակիությունից, ունի 5'- և 3'- ծայրերի նույն կառուցվածքը:
Այսպիսով, 5'-վերջում կա մոդիֆիկացված նուկլեոտիդ 7-մեթիլգուանոզին-5'-տրիֆոսֆատ (գլխարկ): Մի քանի տասնյակ նուկլեոտիդներ առանձնացնում են գլխարկը մեկնարկային կոդոնից, սովորաբար -AUG- եռյակը: Կոդավորման շրջանին հաջորդում է վերջնակետային կոդոններից մեկը՝ UGA-, -UUA-, -UAG-: mRNA-ների մեծ մասի 3' վերջում կա 100-200 ադենոզին մոնոֆոսֆատի մնացորդներից բաղկացած նուկլեոտիդային հաջորդականություն:
- փոխանցող ՌՆԹ-ները հավաքում են ամինաթթուներ բջջի ցիտոպլազմայում և դրանք տեղափոխում ռիբոսոմ; ՌՆԹ-ի այս տիպի մոլեկուլները «սովորում են» սուրհանդակ ՌՆԹ-ի շղթայի համապատասխան հատվածներից, թե որ ամինաթթուները պետք է մասնակցեն սպիտակուցի սինթեզին։
Ցանկացած tRNA-ի տարածական կառուցվածքը, անկախ նուկլեոտիդային հաջորդականության տարբերություններից, նկարագրված է ունիվերսալ երեքնուկային մոդելով։ Յուրաքանչյուր tRNA մոլեկուլ ունի շղթայական հատվածներ, որոնք ներգրավված չեն նուկլեոտիդային մնացորդների միջև ջրածնային կապերի ձևավորման մեջ:
Դրանք ներառում են, մասնավորապես, տեղանքը, որը պատասխանատու է մոլեկուլի 3'-վերջում գտնվող ամինաթթվի հետ կապվելու համար և հակակոդոնը, նուկլեոտիդների հատուկ եռյակ, որը փոխազդում է mRNA կոդոնի հետ կոմպլեմենտար:
- Ռիբոսոմային ՌՆԹ-ն ապահովում է որոշակի կառուցվածքի սպիտակուցի սինթեզ՝ տեղեկատվություն կարդալով տեղեկատվության (մատրիցի) ՌՆԹ-ից: rRNA-ները բարդություններ են կազմում սպիտակուցների հետ, որոնք կոչվում են ռիբոսոմներ:
Յուրաքանչյուր ռիբոսոմ բաղկացած է երկու ենթամիավորներից՝ փոքր (40S) և մեծ (60S): Ռիբոսոմի ենթամիավորները տարբերվում են ոչ միայն rRNA-ների բազմությամբ, այլև սպիտակուցների քանակով և կառուցվածքով։
Հրապարակման ամսաթիվ՝ 2015-02-03; Կարդացեք՝ 2729 | Էջի հեղինակային իրավունքի խախտում

ՌՆԹ-ն պոլիմեր է, որի մոնոմերները ռիբոնուկլեոտիդներ են։
Ի տարբերություն ԴՆԹ-ի՝ ՌՆԹ-ն ձևավորվում է ոչ թե երկու, այլ մեկ պոլինուկլեոտիդային շղթայով (բացառություն՝ որոշ ՌՆԹ պարունակող վիրուսներ ունեն երկշղթա ՌՆԹ)։ ՌՆԹ նուկլեոտիդներն ունակ են միմյանց հետ ջրածնային կապեր ստեղծել։ ՌՆԹ շղթաները շատ ավելի կարճ են, քան ԴՆԹ-ի շղթաները:
ՌՆԹ մոնոմերը՝ նուկլեոտիդ (ռիբոնուկլեոտիդ) - բաղկացած է երեք նյութերի մնացորդներից՝ 1) ազոտային հիմքից, 2) հինգածխածնային մոնոսաքարիդից (պենտոզա) և 3) ֆոսֆորաթթվից։ ՌՆԹ-ի ազոտային հիմքերը նույնպես պատկանում են պիրիմիդինների և պուրինների դասերին։
ՌՆԹ-ի պիրիմիդինային հիմքերը՝ ուրացիլ, ցիտոզին, պուրինային հիմքեր՝ ադենին և գուանին:
31. ՌՆԹ-ի տեսակները և կառուցվածքի առանձնահատկությունները
ՌՆԹ նուկլեոտիդ մոնոսաքարիդը ներկայացված է ռիբոզով:
Գոյություն ունի ՌՆԹ-ի երեք տեսակ՝ 1) տեղեկատվական (մատրիցային) ՌՆԹ – mRNA (mRNA), 2) փոխանցումային ՌՆԹ – tRNA, 3) ռիբոսոմային ՌՆԹ – rRNA:
ՌՆԹ-ի բոլոր տեսակները չճյուղավորված պոլինուկլեոտիդներ են, ունեն հատուկ տարածական կոնֆորմացիա և մասնակցում են սպիտակուցների սինթեզի գործընթացներին։
ՌՆԹ-ի բոլոր տեսակների կառուցվածքի մասին տեղեկատվությունը պահվում է ԴՆԹ-ում: ԴՆԹ կաղապարի վրա ՌՆԹ-ի սինթեզի գործընթացը սովորաբար կոչվում է տրանսկրիպցիա:

Տրանսֆերային ՌՆԹ-ները սովորաբար պարունակում են 76 (75-ից 95) նուկլեոտիդներ; մոլեկուլային քաշը՝ 25000–30000։
tRNA-ն կազմում է բջջի ընդհանուր ՌՆԹ-ի 10%-ը: tRNA-ի գործառույթները. 1) ամինաթթուների տեղափոխում սպիտակուցի սինթեզի վայր, ռիբոսոմներ, 2) թարգմանական միջնորդ. Բջջում հայտնաբերվել է tRNA-ի մոտ 40 տեսակ, որոնցից յուրաքանչյուրն ունի միայն իրեն բնորոշ նուկլեոտիդային հաջորդականություն։ Միևնույն ժամանակ, բոլոր tRNA-ներն ունեն մի քանի ներմոլեկուլային կոմպլեմենտար շրջաններ, որոնց շնորհիվ tRNA-ները ձեռք են բերում ձևով երեքնուկի տերևի նմանվող կոնֆորմացիա։
Ցանկացած tRNA-ն ունի ռիբոսոմի հետ շփման օղակ (1), հակակոդոնային հանգույց (2), ֆերմենտի հետ շփման հանգույց (3), ընդունող ցողուն (4) և հակակոդոն (5): Ամինաթթուն կցվում է ընդունող ցողունի 3' ծայրին: Anticodon - երեք նուկլեոտիդներ, որոնք «ճանաչում են» mRNA կոդոնը:
Պետք է ընդգծել, որ որոշակի tRNA-ն կարող է փոխադրել խիստ սահմանված ամինաթթու, որը համապատասխանում է իր հակակոդոնին: Ամինաթթուների և tRNA-ի միացման առանձնահատկությունը ձեռք է բերվում ամինոացիլ-tRNA սինթետազ ֆերմենտի հատկությունների շնորհիվ:
Ռիբոսոմային ՌՆԹ-ները պարունակում են 3000–5000 նուկլեոտիդներ; մոլեկուլային քաշը՝ 1000000–1500000։
rRNA-ն կազմում է բջջի ընդհանուր ՌՆԹ-ի 80-85%-ը: Ռիբոսոմային սպիտակուցների հետ բարդույթում՝ rRNA-ն ձևավորում է ռիբոսոմներ՝ օրգանելներ, որոնք իրականացնում են սպիտակուցի սինթեզ։ Էուկարիոտիկ բջիջներում rRNA սինթեզը տեղի է ունենում միջուկում: ՌՌՆԹ-ի գործառույթները. 2) ռիբոսոմի և tRNA փոխազդեցության ապահովում. 3) ռիբոսոմի և mRNA նախաձեռնող կոդոնի սկզբնական կապը և ընթերցման շրջանակի որոշումը, 4) ռիբոսոմի ակտիվ կենտրոնի ձևավորումը.
Սուրհանդակային ՌՆԹ-ները բազմազան են նուկլեոտիդների պարունակությամբ և մոլեկուլային քաշով (50000-ից մինչև 4000000):
mRNA-ի մասնաբաժինը կազմում է բջջում ՌՆԹ-ի ընդհանուր պարունակության մինչև 5%-ը: mRNA-ի գործառույթները՝ 1) գենետիկական տեղեկատվության փոխանցում ԴՆԹ-ից ռիբոսոմներ, 2) սպիտակուցի մոլեկուլի սինթեզի մատրիցա, 3) սպիտակուցի մոլեկուլի առաջնային կառուցվածքի ամինաթթուների հաջորդականության որոշում։
Կարդացեք նաև
ՌՆԹ-ն պոլիմեր է, որի մոնոմերները նուկլեոտիդներ են։
Երեք ազոտային հիմքերը նույնն են, ինչ ԴՆԹ-ում (ադենին, գուանին, ցիտոզին); չորրորդը՝ ուրացիլը, առկա է ՌՆԹ-ի մոլեկուլում՝ թիմինի փոխարեն: ՌՆԹ նուկլեոտիդները պարունակում են ռիբոզ՝ դեզօքսիրիբոզի փոխարեն։ ՌՆԹ շղթայում...
ՌՆԹ-ի երեք հիմնական տեսակ. տեղեկատվական(mRNA), կամ մատրիցա(mRNA), ռիբոսոմային(rRNA), և տրանսպորտ(tRNA): Նրանք տարբերվում են մոլեկուլային չափերով և գործառույթներով: ՌՆԹ-ի բոլոր տեսակները սինթեզվում են ԴՆԹ-ի վրա՝ ֆերմենտների՝ ՌՆԹ պոլիմերազների մասնակցությամբ։ Սուրհանդակային ՌՆԹ-ն կազմում է ամբողջ բջջային ՌՆԹ-ի 2-3%-ը, ռիբոսոմայինը՝ 80-85, տրանսպորտը՝ մոտ 15%-ը։
mRNA.
այն կարդում է ժառանգական տեղեկատվությունը ԴՆԹ-ի հատվածից և ազոտային հիմքերի պատճենված հաջորդականության տեսքով այն փոխանցում է ռիբոսոմներին, որտեղ սինթեզվում է որոշակի սպիտակուց։ mRNA մոլեկուլներից յուրաքանչյուրը նուկլեոտիդների կարգով և չափերով համապատասխանում է ԴՆԹ-ի գենին, որից այն արտագրվել է: Միջինում mRNA-ն պարունակում է 1500 նուկլեոտիդ (75-3000)։ mRNA-ի յուրաքանչյուր եռյակ (երեք նուկլեոտիդ) կոչվում է կոդոն:Կոդոնից է կախված, թե սպիտակուցի սինթեզի ժամանակ որ ամինաթթուն կհայտնվի տվյալ տեղում։
(tRNA)ունի համեմատաբար ցածր մոլեկուլային զանգված՝ մոտ 24-29 հազ.
D և պարունակում է 75-ից 90 նուկլեոտիդ մոլեկուլում: Բոլոր tRNA նուկլեոտիդների մինչև 10% -ը փոքր հիմքեր են, որոնք, ըստ երևույթին, պաշտպանում են այն հիդրոլիտիկ ֆերմենտների ազդեցությունից։ Յուրաքանչյուր ամինաթթու կցվում է հատուկ tRNA-ին: Մի շարք ամինաթթուներ ունեն մեկից ավելի tRNA: Մինչ օրս հայտնաբերվել են ավելի քան 60 tRNA-ներ, որոնք տարբերվում են իրենց առաջնային կառուցվածքով (հիմնական հաջորդականությամբ):
Բոլոր tRNA-ների երկրորդական կառուցվածքը ներկայացված է երեքնուկի տերևի տեսքով՝ երկշղթա ցողունով և երեք միաշղթաներով): Շղթաներից մեկի վերջում կա ընդունող տեղամաս՝ CCA եռյակը, որի ադենինին կցված է կոնկրետ ամինաթթու։
(rRNA). Դրանք պարունակում են 120-3100 նուկլեոտիդ։ Ռիբոսոմային ՌՆԹ-ն կուտակվում է միջուկում՝ միջուկներում։
Ռիբոսոմային սպիտակուցները ցիտոպլազմից տեղափոխվում են միջուկներ, և այնտեղ տեղի է ունենում ռիբոսոմային ենթամասնիկների ինքնաբուխ ձևավորում՝ սպիտակուցները համապատասխան rRNA-ի հետ համատեղելով։ Ռիբոսոմի ենթամասնիկները միջուկային մեմբրանի ծակոտիներով միասին կամ առանձին տեղափոխվում են ցիտոպլազմա։ Ռիբոսոմներ 20-30 նմ չափի օրգանելներ են։
Դրանք կառուցված են տարբեր չափերի և ձևերի երկու ենթամասերից։ Բջջում սպիտակուցի սինթեզի որոշակի փուլերում ռիբոսոմները բաժանվում են ենթամասնիկների։
Ռիբոսոմային ՌՆԹ-ն ծառայում է որպես ռիբոսոմների շրջանակ և հեշտացնում է մՌՆԹ-ի սկզբնական կապը ռիբոսոմին սպիտակուցի կենսասինթեզի ընթացքում:
Հարց 6 ԴՆԹ-ի և ՌՆԹ-ի առաջնային և երկրորդային կառուցվածքները կազմող կապերը: ՌՆԹ-ի տեսակները
Գենետիկ կոդը սպիտակուցների ամինաթթուների հաջորդականությունը կոդավորելու միջոց է՝ օգտագործելով նուկլեոտիդների հաջորդականությունը, որը բնորոշ է բոլոր կենդանի օրգանիզմներին։
Հատկություններ՝ 1) գենետիկ կոդը եռյակ(յուրաքանչյուր ամինաթթու կոդավորված է երեք նուկլեոտիդներով); 2) ոչ համընկնող(հարևան եռյակները չունեն ընդհանուր նուկլեոտիդներ); 3) այլասերված(բացառությամբ մեթիոնինի և տրիպտոֆանի, բոլոր ամինաթթուներն ունեն մեկից ավելի կոդոններ); 4) ունիվերսալ(հիմնականում նույնը բոլոր կենդանի օրգանիզմների համար); 5) մեկ ամինաթթվի կոդոններում առաջին երկու նուկլեոտիդները սովորաբար նույնն են, իսկ երրորդը տատանվում է. 6) ունի գծային ընթերցման կարգ և բնութագրվում է համաչափություն,տ.
ե. mRNA-ում կոդոնների կարգի համընկնումը սինթեզված պոլիպեպտիդ շղթայում ամինաթթուների կարգի հետ:
Հրապարակման ամսաթիվ՝ 2014-12-08; Կարդացեք՝ 11268 | Էջի հեղինակային իրավունքի խախտում
studopedia.org - Studopedia.Org - 2014-2018. (0.001 վ) ...
Բջիջների ցիտոպլազմը պարունակում է ՌՆԹ-ի երեք հիմնական ֆունկցիոնալ տեսակ.
- սուրհանդակ ՌՆԹ (mRNA), որը գործում է որպես սպիտակուցների սինթեզի ձևանմուշներ.
- ռիբոսոմային ՌՆԹ (rRNA), որը գործում է որպես ռիբոսոմների կառուցվածքային բաղադրիչներ;
- փոխանցել ՌՆԹ-ները (tRNAs), որոնք ներգրավված են mRNA տեղեկատվության թարգմանության (թարգմանության) մեջ սպիտակուցի մոլեկուլի ամինաթթուների հաջորդականության մեջ:
Բջիջների միջուկում հայտնաբերվում է միջուկային ՌՆԹ, որը կազմում է ընդհանուր բջջային ՌՆԹ-ի 4-ից 10%-ը:
Միջուկային ՌՆԹ-ի հիմնական մասը ներկայացված է ռիբոսոմային և տրանսֆերային ՌՆԹ-ի բարձր մոլեկուլային պրեկուրսորներով: Բարձր մոլեկուլային քաշի rRNA-ների պրեկուրսորները (28 S, 18 S և 5 S RNA) հիմնականում տեղայնացված են միջուկում։
ՌՆԹ-ն կենդանական և բուսական որոշ վիրուսների (գենոմային ՌՆԹ) հիմնական գենետիկ նյութն է։ ՌՆԹ վիրուսների մեծամասնությանը բնորոշ է իրենց ՌՆԹ գենոմի հակադարձ տրանսկրիպցիան՝ ուղղված հակադարձ տրանսկրիպտազին։
Բոլոր ռիբոնուկլեինաթթուները ռիբոնուկլեոտիդների պոլիմերներ են, որոնք կապված են, ինչպես ԴՆԹ-ի մոլեկուլում, 3',5'-ֆոսֆորոդիեսթեր կապերով:
Ի տարբերություն ԴՆԹ-ի, որն ունի երկշղթա կառուցվածք, ՌՆԹ-ն միաշղթա գծային պոլիմերային մոլեկուլ է։
mRNA կառուցվածքը. mRNA-ն ՌՆԹ-ի առավել տարասեռ դասն է չափի և կայունության տեսանկյունից:
tRNA կառուցվածքը.
Տրանսֆերային ՌՆԹ-ները մՌՆԹ թարգմանության ժամանակ հանդես են գալիս որպես միջնորդներ (ադապտերներ): Նրանք կազմում են ընդհանուր բջջային ՌՆԹ-ի մոտավորապես 15%-ը: 20 պրոտեինոգեն ամինաթթուներից յուրաքանչյուրն ունի իր սեփական tRNA: Որոշ ամինաթթուների համար, որոնք կոդավորված են երկու կամ ավելի կոդոններով, կան մի քանի tRNA:
tRNA-ները համեմատաբար փոքր միաշղթա մոլեկուլներ են, որոնք բաղկացած են 70-93 նուկլեոտիդներից։ Նրանց մոլեկուլային զանգվածը (2,4-3,1) ,104 կԴա է։
tRNA-ի երկրորդական կառուցվածքը ձևավորվում է ազոտային հիմքերի ներմոլեկուլային կոմպլեմենտար զույգերի միջև առավելագույն թվով ջրածնային կապերի ձևավորման շնորհիվ։
Այս կապերի ձևավորման արդյունքում tRNA պոլինուկլեոտիդային շղթան պտտվում է պարուրաձև ճյուղերի ձևավորմամբ, որոնք ավարտվում են չզույգված նուկլեոտիդների օղակներով։ Բոլոր tRNA-ների երկրորդական կառուցվածքների տարածական պատկերն ունի երեքնուկի տերևի տեսք։
«Երեքնուկի տերեւում» առանձնանում են չորս պարտադիր ճյուղեր, ավելի երկար tRNA-ները, բացի այդ, պարունակում են կարճ հինգերորդ (լրացուցիչ) ճյուղ։
tRNA-ի ադապտերային ֆունկցիան ապահովվում է ընդունող ճյուղով, որի 3' ծայրին ամինաթթուների մնացորդը կցվում է եթերային կապով, և հակակոդոնային ճյուղը հակառակ ակցեպտորի ճյուղին, որի վերևում կա մի օղակ, որը պարունակում է հակակոդոն:
Հակոդոնը նուկլեոտիդների հատուկ եռյակ է, որը հակազուգահեռ ուղղությամբ լրացնում է համապատասխան ամինաթթուն կոդավորող mRNA կոդոնին:
Պսեւդուրիդինային հանգույցը կրող T-ճյուղը (TyC-loop) ապահովում է tRNA-ի փոխազդեցությունը ռիբոսոմների հետ:
D-ճյուղը, որը կրում է դեհիդրոուրիդինի հանգույցը, ապահովում է tRNA-ի փոխազդեցությունը համապատասխան aminoacyl-tRNA սինթետազի հետ:

tRNA-ի երկրորդական կառուցվածքը
Հինգերորդ լրացուցիչ ճյուղի գործառույթները դեռ վատ են հասկացված, ամենայն հավանականությամբ, այն հավասարեցնում է տարբեր tRNA մոլեկուլների երկարությունը:
tRNA-ի երրորդական կառուցվածքը շատ կոմպակտ է և ձևավորվում է ջրածնային լրացուցիչ կապերի շնորհիվ երեքնուկի տերևի առանձին ճյուղերի համախմբմամբ՝ ձևավորելով L-աձև «արմունկի թեքում» կառուցվածք:
Տրանսպորտային ՌՆԹ, կառուցվածք և ֆունկցիոնալ մեխանիզմ:
Այս դեպքում ամինաթթուն կապող ակցեպտորը գտնվում է մոլեկուլի մի ծայրում, իսկ հակակոդոնը՝ մյուսում։

tRNA-ի երրորդային կառուցվածքը (ըստ A.S. Spirin-ի)
rRNA-ի և ռիբոսոմների կառուցվածքը. Ռիբոսոմային ՌՆԹ-ները կազմում են այն ողնաշարը, որի հետ կապվում են հատուկ սպիտակուցներ՝ առաջացնելով ռիբոսոմներ: Ռիբոսոմները նուկլեոպրոտեինային օրգանելներ են, որոնք ապահովում են սպիտակուցի սինթեզ mRNA-ից:
Բջջում ռիբոսոմների թիվը շատ մեծ է՝ 104-ից պրոկարիոտներում մինչև 106 էուկարիոտներում: Ռիբոսոմները տեղայնացված են հիմնականում ցիտոպլազմայում, էուկարիոտներում, բացի այդ, միջուկում, միտոքոնդրիումային մատրիցայում և քլորոպլաստների ստրոմայում։ Ռիբոսոմները բաղկացած են երկու ենթամասերից՝ մեծ և փոքր։ Ըստ չափի և մոլեկուլային քաշի՝ բոլոր ուսումնասիրված ռիբոսոմները բաժանվում են 3 խմբի՝ պրոկարիոտների 70S ռիբոսոմներ (S-նստվածքի գործակից), որոնք բաղկացած են փոքր 30S և մեծ 50S ենթամասնիկներից; 80S էուկարիոտիկ ռիբոսոմներ՝ կազմված 40S փոքր և 60S մեծ ենթամիավորներից։
80S ռիբոսոմների փոքր ենթամիավորը ձևավորվում է մեկ rRNA մոլեկուլով (18S) և տարբեր սպիտակուցների 33 մոլեկուլներով։
Մեծ ենթամիավորը ձևավորվում է երեք rRNA մոլեկուլներով (5S, 5.8S և 28S) և մոտավորապես 50 սպիտակուցներով։
rRNA-ի երկրորդական կառուցվածքը ձևավորվում է մոլեկուլի կարճ երկշղթա հատվածների շնորհիվ՝ մազակալներ (rRNA-ի մոտ 2/3), 1/3-ը ներկայացված է պուրինային նուկլեոտիդներով հարուստ միաշղթա հատվածներով։
Սոցիալական կոճակներ Joomla-ի համար
Սպիտակուցները կազմում են կյանքի հիմքը։ Նրանց գործառույթները խցում շատ բազմազան են: Այնուամենայնիվ, սպիտակուցները «չեն կարող» վերարտադրվել։ Իսկ սպիտակուցների կառուցվածքի մասին ամբողջ տեղեկատվությունը պարունակվում է գեներում (ԴՆԹ):
Բարձրագույն օրգանիզմներում սպիտակուցները սինթեզվում են բջջի ցիտոպլազմայում, իսկ ԴՆԹ-ն թաքնված է միջուկի թաղանթի հետևում։ Հետևաբար, ԴՆԹ-ն չի կարող ուղղակիորեն ծառայել որպես սպիտակուցի սինթեզի ձևանմուշ։ Այս դերը կատարում է մեկ այլ նուկլեինաթթու՝ ՌՆԹ:
ՌՆԹ-ի մոլեկուլը երրորդական կառուցվածքով չճյուղավորված պոլինուկլեոտիդ է։
Այն ձևավորվում է մեկ պոլինուկլեոտիդային շղթայով, և չնայած դրանում ընդգրկված կոմպլեմենտար նուկլեոտիդները կարող են նաև ջրածնային կապեր ստեղծել միմյանց միջև, այդ կապերը տեղի են ունենում մեկ շղթայի նուկլեոտիդների միջև։ ՌՆԹ շղթաները շատ ավելի կարճ են, քան ԴՆԹ-ի շղթաները: Եթե բջիջում ԴՆԹ-ի պարունակությունը համեմատաբար հաստատուն է, ապա ՌՆԹ-ի պարունակությունը մեծ տատանվում է։ Բջիջներում ՌՆԹ-ի ամենամեծ քանակությունը նկատվում է սպիտակուցի սինթեզի ժամանակ։
ՌՆԹ-ն մեծ դեր է խաղում ժառանգական տեղեկատվության փոխանցման և իրականացման գործում:
Գործառույթին և կառուցվածքային առանձնահատկություններին համապատասխան առանձնանում են բջջային ՌՆԹ-ի մի քանի դասեր։
Բջջային ՌՆԹ-ի երեք հիմնական դաս կա.
- Տեղեկատվական (mRNA) կամ մատրիցա (mRNA): Նրա մոլեկուլներն ամենատարբերն են չափերով, մոլեկուլային քաշով (0,05x106-ից մինչև 4x106) և կայունությամբ։
Դրանք կազմում են բջջի ՌՆԹ-ի ընդհանուր քանակի մոտ 2%-ը։ Բոլոր mRNA-ները գենետիկ տեղեկատվության կրողներ են միջուկից մինչև ցիտոպլազմա՝ մինչև սպիտակուցի սինթեզի վայր։ Նրանք ծառայում են որպես մատրիցա (աշխատանքային գծագիր) սպիտակուցի մոլեկուլի սինթեզի համար, քանի որ որոշում են սպիտակուցի մոլեկուլի ամինաթթուների հաջորդականությունը (առաջնային կառուցվածքը)։

- Ռիբոսոմային ՌՆԹ (rRNA):
Նրանք կազմում են բջջի ընդհանուր ՌՆԹ պարունակության 80–85%-ը։
31. ՌՆԹ-ի կառուցվածքը. ՌՆԹ-ի տեսակները, կառուցվածքային առանձնահատկությունները և գործառույթները: tRNA-ի երկրորդական կառուցվածքը
Ռիբոսոմային ՌՆԹ-ն բաղկացած է 3–5 հազար նուկլեոտիդներից։ Այն սինթեզվում է միջուկի միջուկներում։ Ռիբոսոմային սպիտակուցների հետ բարդույթում՝ rRNA-ն ձևավորում է ռիբոսոմներ՝ օրգանելներ, որոնց վրա հավաքվում են սպիտակուցի մոլեկուլները։ RRNA-ի հիմնական նշանակությունն այն է, որ այն ապահովում է mRNA-ի և ռիբոսոմի նախնական կապը և կազմում է ռիբոսոմի ակտիվ կենտրոնը, որում պոլիպեպտիդային շղթայի սինթեզի ընթացքում ամինաթթուների միջև ձևավորվում են պեպտիդային կապեր։
- Տրանսֆերային ՌՆԹ (tRNAs):
tRNA մոլեկուլները սովորաբար պարունակում են 75-86 նուկլեոտիդ: tRNA մոլեկուլների մոլեկուլային քաշը կազմում է մոտ 25 հազար: tRNA մոլեկուլները միջնորդների դեր են խաղում սպիտակուցների կենսասինթեզի մեջ. նրանք ամինաթթուները առաքում են սպիտակուցի սինթեզի վայր, այսինքն ՝ ռիբոսոմներ: Բջիջը պարունակում է ավելի քան 30 տեսակի tRNA: tRNA-ի յուրաքանչյուր տեսակ ունի իր յուրահատուկ նուկլեոտիդային հաջորդականությունը:
Այնուամենայնիվ, բոլոր մոլեկուլներն ունեն մի քանի ներմոլեկուլային կոմպլեմենտար շրջաններ, որոնց առկայության շնորհիվ բոլոր tRNA-ները ունեն երրորդական կառուցվածք, որն իր ձևով նման է երեքնուկի տերևին:
ՌՆԹ-ի երկրորդական կառուցվածքը- tRNA-ին բնորոշ՝ միաշղթա, «երեքնուկի տերևի» ձևով։
Ներառում է.
- համեմատաբար կարճ կրկնակի պարուրակներ՝ ցողուններ,
- միաշղթա հատվածներ - հանգույցներ:
Կան 4 ցողուններ (ընդունիչ, հակակոդոն, դիհիդրոուրիդիլ, պսևդուրիդիլ) և 3 օղակ։

«Ցողունային հանգույց» - ՌՆԹ-ի երկրորդական կառուցվածքի տարր, սխեմատիկորեն

«Pseudoknot» - ՌՆԹ-ի երկրորդական կառուցվածքի տարր, սխեմատիկորեն

Ընդունող ցողունը պարունակում է պոլինուկլեոտիդային շղթայի 3'- և 5' ծայրերը, 5' ծայրը վերջանում է գուանիլաթթվի մնացորդով, 3' ծայրը CCA եռյակ է և ծառայում է AA-ի հետ էսթերային կապ ստեղծելուն:
Հակոդոնային ցողունը ճանաչում է իր կոդոնը mRNA-ի վրա ռիբոսոմներում կոմպլեմենտարության սկզբունքով։
Պսեւդուրիդիլ ցողունը ծառայում է ռիբոսոմին միանալու համար։
Դիհիդրոուրիդիլ ցողունը ծառայում է aminoacyl-tRNA սինթետազին կապելու համար:
Սոցիալական կոճակներ Joomla-ի համար
Մենք նաև խորհուրդ ենք տալիս
 Ի՞նչ SIM է անհրաժեշտ iPhone-ի համար:
Ի՞նչ SIM է անհրաժեշտ iPhone-ի համար:
 Ինչպես լրացնել ձեր Webmoney հաշիվը ձեր հեռախոսից
Ինչպես լրացնել ձեր Webmoney հաշիվը ձեր հեռախոսից
 iPhone-ում զանգի ձայնը սահմանելու բոլոր եղանակները
iPhone-ում զանգի ձայնը սահմանելու բոլոր եղանակները
 Հասցեներ և հեռախոսահամարներ Ճապոնիայում Ինչպես զանգահարել Ճապոնիայից Ռուսաստան
Հասցեներ և հեռախոսահամարներ Ճապոնիայում Ինչպես զանգահարել Ճապոնիայից Ռուսաստան
 Հեռակառավարում հեռուստացույցի համար բջջային հեռախոսում. ինչպե՞ս կառավարել հեռուստացույցը Android-ի միջոցով:
Հեռակառավարում հեռուստացույցի համար բջջային հեռախոսում. ինչպե՞ս կառավարել հեռուստացույցը Android-ի միջոցով:
 Ածխածնի մանրաթելից վինիլային ֆիլմեր
Ածխածնի մանրաթելից վինիլային ֆիլմեր