Kāds ir transkripcijas process? RNS saņem iedzimtu informāciju
Transkripcijas uzsākšana
Transkripcijas pagarinājums
Brīdis, kurā RNS polimerāze pāriet no transkripcijas ierosināšanas uz pagarinājumu, nav precīzi noteikts. Trīs galvenie bioķīmiski notikumi raksturo šo pāreju Escherichia coli RNS polimerāzes gadījumā: sigmas faktora izdalīšanās, pirmā fermenta molekulas pārvietošana gar šablonu un spēcīga transkripcijas kompleksa stabilizācija, kas papildus RNS. polimerāze ietver augošo RNS ķēdi un transkribēto DNS. Tādas pašas parādības ir raksturīgas arī eikariotu RNS polimerāzēm. Pāreju no iniciācijas uz pagarināšanu pavada saišu pārrāvums starp enzīmu, promotoru, transkripcijas iniciācijas faktoriem un dažos gadījumos RNS polimerāzes pāreju uz pagarināšanas kompetences stāvokli (piemēram, CTD domēna fosforilēšanās RNS polimerāze II). Pagarinājuma fāze beidzas pēc augošā transkripta atbrīvošanas un fermenta atdalīšanās no veidnes (pārtraukšana).
Pagarināšana tiek veikta ar pagarinājuma pamatfaktoru palīdzību, kas nepieciešami, lai process priekšlaicīgi neapstātos.
Nesen ir parādījušies pierādījumi, ka regulējošie faktori var arī regulēt pagarinājumu. Pagarināšanas procesa laikā RNS polimerāze apstājas noteiktās gēna daļās. Tas ir īpaši skaidri redzams zemās substrātu koncentrācijās. Dažos matricas apgabalos ir ilgstoša RNS polimerāzes progresēšanas kavēšanās, tā sauktā. pauzes tiek novērotas pat pie optimālām substrāta koncentrācijām. Šo paužu ilgumu var kontrolēt ar pagarinājuma faktoriem.
Izbeigšana
Baktērijām ir divi transkripcijas pārtraukšanas mehānismi:
- no rho atkarīgs mehānisms, kurā Rho (rho) proteīns destabilizē ūdeņraža saites starp DNS šablonu un mRNS, atbrīvojot RNS molekulu.
- rho-neatkarīgs, kurā transkripcija apstājas, kad tikko sintezētā RNS molekula veido cilmes cilpu, kam seko vairāki uracili (...UUUU), kas noved pie RNS molekulas atdalīšanās no DNS veidnes.
Transkripcijas pārtraukšana eikariotos ir mazāk pētīta. Tas beidzas ar RNS pārgriešanu, pēc tam ferments savam 3" galam pievieno vairākus adenīnus (...AAAA), kuru skaits nosaka dotā transkripta stabilitāti.
Transkripcijas rūpnīcas
Ir vairāki eksperimentāli dati, kas liecina, ka transkripcija notiek tā sauktajās transkripcijas rūpnīcās: milzīgi, pēc dažām aplēsēm, līdz 10 Da kompleksi, kas satur apmēram 8 RNS polimerāzes II un komponentus turpmākai apstrādei un savienošanai, kā arī korekcijai. no tikko sintezētā stenogramma. Šūnas kodolā notiek pastāvīga apmaiņa starp šķīstošās un aktivētās RNS polimerāzes kopumiem. Šādā kompleksā ir iesaistīta aktīvā RNS polimerāze, kas savukārt ir strukturāla vienība, kas organizē hromatīna blīvēšanu. Jaunākie dati liecina, ka transkripcijas rūpnīcas pastāv pat tad, ja nav transkripcijas, tās ir fiksētas šūnā (vēl nav skaidrs, vai tās mijiedarbojas ar šūnas kodolmatricu vai nē) un ir neatkarīgs kodola apakšnodalījums. Transkripcijas rūpnīcas komplekss, kas satur RNS polimerāzi I, II vai III, tika analizēts ar masas spektrometriju.
Reversā transkripcija
Reversās transkripcijas shēmaDažiem vīrusiem (piemēram, HIV, kas izraisa AIDS) ir iespēja pārrakstīt RNS DNS. HIV ir RNS genoms, kas ir integrēts DNS. Tā rezultātā vīrusa DNS var apvienot ar saimniekšūnas genomu. Galvenais enzīms, kas atbild par DNS sintezēšanu no RNS, tiek saukts par reverso. Viena no reversetāzes funkcijām ir radīt komplementāru DNS (cDNS) no vīrusa genoma. Saistītais enzīms ribonukleāze H šķeļ RNS, un reverseāze sintezē cDNS no DNS dubultās spirāles. cDNS ir integrēta saimniekšūnas genomā ar integrāzes palīdzību. Rezultātā saimniekšūna sintezē vīrusu proteīnus, kas veido jaunus vīrusus. HIV gadījumā tiek ieprogrammēta arī T-limfocītu apoptoze (šūnu nāve). Citos gadījumos šūna var palikt vīrusu izplatītāja.
Dažas eikariotu šūnas satur enzīmu telomerāzi, kas arī uzrāda reversās transkripcijas aktivitāti. Ar tās palīdzību tiek sintezētas atkārtotas DNS sekvences. Telomerāze bieži tiek aktivizēta vēža šūnās, lai neierobežoti dublētu genomu, nezaudējot proteīnu kodējošo DNS secību.
Piezīmes
Wikimedia fonds. 2010. gads.
Skatiet, kas ir “Transkripcija (bioloģija)” citās vārdnīcās:
- (no latīņu transcriptio, lit. pārrakstīšana), RNS molekulu biosintēze, resp. DNS sekcijas; ģenētiskās ieviešanas pirmais posms. informācija dzīvās šūnās. To veic no DNS atkarīgā enzīma RNS polimerāze līdz visvairāk pētīto paradīzei... ... Bioloģiskā enciklopēdiskā vārdnīca
bioloģija- BIOLOĢIJA (no grieķu vārda bio life un logos, doktrīna) zinātņu kopums par dzīvību visās tās formu, īpašību, savienojumu un attiecību izpausmju daudzveidībā uz Zemes. Šis termins pirmo reizi tika ierosināts vienlaicīgi un neatkarīgi 1802. gadā... ... Epistemoloģijas un zinātnes filozofijas enciklopēdija
Dzīves zinātne, kas ietver visas zināšanas par dzīvo būtņu dabu, struktūru, funkcijām un uzvedību. Bioloģija nodarbojas ne tikai ar dažādu organismu formu lielo dažādību, bet arī ar to evolūciju, attīstību un tām attiecībām, kas... ... Koljēra enciklopēdija
BIOLOĢIJA- zinātņu kopums par dzīvi visās tās formu, īpašību, savienojumu un attiecību izpausmju daudzveidībā uz Zemes. Pirmo reizi šo terminu vienlaikus un neatkarīgi viens no otra 1802. gadā ierosināja izcilais franču zinātnieks J.B. Lamarks un vācietis...... Zinātnes filozofija: pamatterminu vārdnīca
I Transkripcija (no latīņu valodas transcriptio rewriting) vārdu un tekstu rakstiska reproducēšana, ņemot vērā to izrunu, izmantojot noteiktu grafisko sistēmu. T. var būt zinātnisks un praktisks. Zinātniskā T. lieto lingvistiskajā...
- (no latīņu transcriptio, burtu pārrakstīšana), RNS biosintēze uz DNS matricas; ģenētiskās ieviešanas pirmais posms. informācija, griešanas gaitā DNS nukleotīdu secība tiek nolasīta RNS nukleotīdu secības formā (skat. Ģenētiskais kods) ... Ķīmiskā enciklopēdija
Iepriekšēja mRNS ar cilmes cilpu. Slāpekļa atomi bāzēs ir izcelti zilā krāsā, skābekļa atomi molekulas fosfāta mugurkaulā sarkanā krāsā Ribonukleīnskābes (RNS) ir nukleīnskābes, nukleotīdu polimēri, kas satur ortofosforskābes atlikumu ... Wikipedia
Zinātne, kuras mērķis ir izprast dzīvības parādību būtību, pētot bioloģiskos objektus un sistēmas līmenī, kas tuvojas molekulārajam līmenim un dažos gadījumos sasniedz šo robežu. Galīgais mērķis ir...... Lielā padomju enciklopēdija
Reversā transkripcija ir divpavedienu DNS iegūšanas process no vienpavedienu RNS veidnes. Šo procesu sauc par reverso transkripciju, jo ģenētiskās informācijas pārsūtīšana notiek "reversā", nosacīti ... ... Wikipedia
Pieprasījums "Vīruss" tiek novirzīts uz šejieni. Skat arī citas nozīmes. ? Vīrusi Rotavīruss Zinātniskā klasifikācija Overkingdom ... Wikipedia
Pēc ģenētiskā koda atšifrēšanas radās jautājums: kā informācija tiek pārnesta no DNS uz proteīnu? Bioķīmiskie pētījumi ir atklājuši, ka lielākā daļa DNS šūnā ir lokalizēta kodolā, bet olbaltumvielu sintēze notiek citoplazmā. Šī DNS un olbaltumvielu sintēzes teritoriālā atdalīšana noveda pie starpnieka meklējumiem. Tā kā olbaltumvielu sintēze notika, piedaloties ribosomām, RNS tika izvirzīta starpnieka lomai. Tika izveidota diagramma, kas ilustrē ģenētiskās informācijas plūsmas virzienu šūnā:
DNS → RNS → proteīns
To sauc par molekulārās bioloģijas centrālo dogmu. F. Kriks postulēja, ka makromolekulu sintēze saskaņā ar šo shēmu tiek veikta pēc matricas principa. Bija vajadzīgi daudzi gadi, lai pierādītu šī postulāta pareizību.
Sākumā tika pieņemts, ka ribosomu RNS (“viens gēns - viena ribosoma - viens proteīns”) spēlēja starpnieka lomu. Tomēr drīz vien kļuva skaidrs, ka šis pieņēmums nav attaisnojams. Ir pierādīts, ka proteīnu sintēzes laikā ribosomu skaits nemainās, t.i. jauna RNS netiek sintezēta un līdz ar to netiek saņemta jauna informācija. Drīz vien ribosomu sastāvā tika atklāta nestabilas RNS daļa, kuras molekulas ar Mg katjonu palīdzību brīvi noturas uz ribosomas. Izmantojot molekulāro hibridizāciju, tika parādīts, ka šīs RNS molekulas ir noteiktu DNS sekciju kopijas. Viņa ieguva vārdu matrica, vai kurjers RNS. Iepriekš to sauca arī par kurjera RNS un ziņojuma RNS. Šo molekulu komplementaritāte ar noteiktām DNS sekcijām liecināja, ka tās tika sintezētas saskaņā ar DNS šablona tipu.
Pamazām tika noskaidrots viss informācijas pārnešanas ceļš no DNS uz proteīnu. Tas sastāv no diviem posmiem: transkripcijas Un raidījumi. Transkripcijas stadijā ģenētiskā informācija tiek nolasīta un pārnesta no DNS uz mRNS. Transkripcijas process notiek trīs posmos: iniciācija, pagarinājums Un izbeigšanu. Informācija tiek nolasīta tikai no vienas DNS ķēdes (+ ķēde), jo, pamatojoties uz ģenētiskā koda īpašībām, komplementārās DNS sekcijas nevar kodēt viena un tā paša proteīna struktūru, jo trūkst koda komplementāras deģenerācijas. Transkripciju veic enzīms RNS polimerāze, kas sastāv no četrām apakšvienībām (ααββ") un kurai nav specifiskuma attiecībā uz DNS avotu. Transkripcijas sākotnējā stadijā - iniciācijā - piektā apakšvienība, tā sauktais s-faktors , ir pievienots fermentam, kas atpazīst noteiktu DNS sekciju, promotoru.Promoteri netiek transkribēti Tos atpazīst pēc s-faktora pēc noteiktas nukleotīdu sekvences klātbūtnes tajos Baktēriju promotoros to sauc par Pribnovu. bloks un ir forma TATAAT (ar nelielām variācijām).Pie promotora ir piesaistīts enzīms RNS polimerāze.mRNS ķēdes augšana notiek vienā virzienā,transkripcijas ātrums ≈ 45-50 nukleotīdi sekundē.Iniciācijas stadijā tiek sintezēta tikai īsa 8 nukleotīdu ķēde, pēc kuras s-faktors tiek atdalīts no RNS polimerāzes un sākas pagarinājuma stadija MRNS ķēdes pagarināšanu veic tetramēra proteīns Sadaļa , no kuras tiek nolasīta informācija, ir ko sauc par transkriptonu.Tas beidzas ar terminatoru - noteiktu nukleotīdu secību, kas spēlē apturēšanas signāla lomu. Sasniedzot terminatoru, RNS polimerāzes enzīms pārstāj darboties un ar proteīna terminācijas faktoru palīdzību tiek atdalīts no matricas.

Baktēriju šūnās iegūtās mRNS molekulas uzreiz var kalpot par proteīnu sintēzes šabloniem, t.i. pārraide. Tie savienojas ar ribosomām, kurām RNS (tRNS) transportēšanas molekulas vienlaikus piegādā aminoskābes. Pārneses RNS ķēdes sastāv no aptuveni 70 nukleotīdiem. Vienpavedienu tRNS molekulai ir komplementāras savienošanās vietas, kurās ir aktīvi centri: vieta tRNS atpazīšanai ar enzīmu tRNS sintetāzi, kas piesaista tRNS atbilstošo aktivēto aminoskābi; akceptors - vieta, kurai pievienota aminoskābe, un antikodona cilpa.

Antikodons ir triplets, kas komplementārs ar atbilstošo kodonu mRNS molekulā. Kodona un antikodona mijiedarbība notiek pēc komplementārā savienojuma veida, kura laikā augošajai proteīna ķēdei tiek pievienota aminoskābe. Sākuma kodons dažādās mRNS ir AUG kodons, kas atbilst aminoskābes metionīnam. Tāpēc tRNS ar UAC antikodonu, kas savienots ar aktivēto aminoskābi metionīnu, ir pirmā, kas tuvojas matricai. Fermentus, kas aktivizē aminoskābes un savieno tās ar tRNS, sauc par aminoacil-tRNS sintetāzēm. Visus proteīnu biosintēzes posmus (iniciāciju, pagarinājumu, izbeigšanu) apkalpo proteīnu translācijas faktori. Prokariotiem katrā posmā ir trīs no tiem. MRNS veidnes beigās ir bezjēdzīgi kodoni, kas netiek nolasīti un iezīmē tulkošanas beigas.
Daudzu organismu genomā, sākot no baktērijām līdz cilvēkiem, ir atklāti gēni un atbilstošas tRNS, kas veic nestandarta kodonu nolasīšanu. Šo fenomenu sauc pārraides neskaidrības.

Tas ļauj izvairīties no kļūdu negatīvajām sekām, kas rodas mRNS molekulu struktūrā transkripcijas laikā. Tādējādi, kad mRNS molekulas iekšpusē parādās absurdi kodoni, kas spēj priekšlaicīgi apturēt transkripcijas procesu, tiek aktivizēts nomākšanas mehānisms. Tas sastāv no tā, ka šūnā parādās neparasta tRNS forma ar antikodonu, kas ir komplementārs nonsense kodonam, kuram parasti nevajadzētu pastāvēt. Tās parādīšanās ir gēna darbības rezultāts, kas aizstāj bāzi tRNS antikodonā, kas pēc sastāva ir līdzīgs bezjēdzīgajam kodonam. Šīs aizstāšanas rezultātā bezjēdzīgais kodons tiek lasīts kā regulārs nozīmīgais kodons. Šādas mutācijas sauc par supresoru mutācijām, jo tie nomāc sākotnējo mutāciju, kas noveda pie nonsense kodona.
Dzīve oglekļa formā pastāv olbaltumvielu molekulu klātbūtnes dēļ. Un proteīnu biosintēze šūnā ir vienīgā gēnu ekspresijas iespēja. Bet, lai īstenotu šo procesu, ir jāuzsāk vairāki procesi, kas saistīti ar ģenētiskās informācijas “izpakošanu”, vajadzīgā gēna meklēšanu, lasīšanu un reproducēšanu. Termins “transkripcija” bioloģijā īpaši attiecas uz informācijas pārsūtīšanas procesu no gēna uz ziņojuma RNS. Tas ir biosintēzes sākums, tas ir, tieša ģenētiskās informācijas ieviešana.
Ģenētiskās informācijas uzglabāšana
Dzīvu organismu šūnās ģenētiskā informācija ir lokalizēta kodolā, mitohondrijās, hloroplastos un plazmīdos. Mitohondriji un hloroplasti satur nelielu daudzumu dzīvnieku un augu DNS, savukārt baktēriju plazmīdas ir gēnu uzglabāšanas vieta, kas atbild par ātru pielāgošanos vides apstākļiem.
Vīrusu ķermeņos iedzimtā informācija tiek glabāta arī RNS vai DNS polimēru veidā. Bet tā ieviešanas process ir saistīts arī ar transkripcijas nepieciešamību. Bioloģijā šim procesam ir ārkārtīgi liela nozīme, jo tas noved pie iedzimtas informācijas ieviešanas, izraisot olbaltumvielu biosintēzi.

Dzīvnieku šūnās iedzimto informāciju attēlo DNS polimērs, kas ir kompakti iesaiņots kodola iekšpusē. Tāpēc pirms proteīnu sintēzes vai jebkura gēna nolasīšanas ir jāiziet noteikti posmi: kondensētā hromatīna attīšana un vēlamā gēna “atbrīvošana”, tā atpazīšana ar enzīmu molekulām, transkripcija.
Bioloģijā un bioloģiskajā ķīmijā šie posmi jau ir pētīti. Tie noved pie proteīna sintēzes, kuras primārā struktūra tika kodēta vienā gēnā.
Transkripcijas modelis eikariotu šūnās
Lai gan transkripcija bioloģijā nav pietiekami pētīta, tās secība tradicionāli tiek parādīta diagrammas veidā. Tas sastāv no iniciācijas, pagarināšanas un izbeigšanas. Tas nozīmē, ka viss process ir sadalīts trīs komponentu parādībās.

Iniciācija ir bioloģisko un bioķīmisko procesu kopums, kas noved pie transkripcijas sākuma. Pagarinājuma būtība ir nepārtraukta molekulārās ķēdes augšana. Izbeigšana ir procesu kopums, kas noved pie RNS sintēzes pārtraukšanas. Starp citu, proteīnu biosintēzes kontekstā transkripcijas process bioloģijā parasti tiek identificēts ar ziņneša RNS sintēzi. Pamatojoties uz to, vēlāk tiks sintezēta polipeptīdu ķēde.

Iniciācija
Iniciācija ir vismazāk saprotamais transkripcijas mehānisms bioloģijā. Kas tas ir no bioķīmiskā viedokļa, nav zināms. Tas nozīmē, ka īpašie fermenti, kas ir atbildīgi par transkripcijas izraisīšanu, vispār netiek atpazīti. Tāpat nav zināmi intracelulārie signāli un to pārraides metodes, kas norāda uz nepieciešamību pēc jauna proteīna sintēzes. Tas ir citoloģijas un bioķīmijas pamatuzdevums.
Pagarinājums
Pagaidām nav iespējams nodalīt iniciācijas un pagarinājuma procesu laikā, jo nav iespējams veikt laboratorijas pētījumus, kas paredzēti, lai apstiprinātu konkrētu enzīmu un sprūda faktoru klātbūtni. Tāpēc šī robeža ir ļoti nosacīta. Pagarināšanas procesa būtība ir saistīta ar augšanas ķēdes pagarināšanu, kas sintezēta, pamatojoties uz DNS veidnes sadaļu.
Tiek uzskatīts, ka pagarinājums sākas pēc pirmās RNS polimerāzes translokācijas un pirmā kadona pievienošanās sākuma RNS sākuma vietai. Pagarināšanas laikā kadonus nolasa 3"-5" virknes virzienā uz despiralizētas DNS daļas, kas sadalīta divās daļās. Tajā pašā laikā augošā RNS ķēde tiek pievienota ar jauniem nukleotīdiem, kas papildina šablona DNS reģionu. Šajā gadījumā DNS tiek “paplašināta” līdz 12 nukleotīdu platumam, tas ir, 4 kadoniem.

Enzīms RNS polimerāze pārvietojas pa augšanas ķēdi, un “aiz tās” DNS ir apgriezti “savienota” divpavedienu struktūrā, atjaunojot ūdeņraža saites starp nukleotīdiem. Tas daļēji atbild uz jautājumu par to, kādu procesu bioloģijā sauc par transkripciju. Tieši pagarinājums ir galvenā transkripcijas fāze, jo tās gaitā tiek samontēts tā sauktais starpnieks starp gēnu un proteīna sintēzi.
Izbeigšana
Transkripcijas pārtraukšanas process eikariotu šūnās ir slikti izprotams. Līdz šim zinātnieki ir samazinājuši tā būtību, pārtraucot DNS nolasīšanu 5 collu galā un pievienojot adenīna bāzu grupu RNS 3 collu galam. Pēdējais process ļauj stabilizēt iegūtās RNS ķīmisko struktūru. Baktēriju šūnās ir divu veidu izbeigšanās. Tas ir no Rho atkarīgs un no Ro neatkarīgs process.

Pirmais notiek Rho proteīna klātbūtnē un tiek samazināts līdz vienkāršai ūdeņraža saišu pārtraukšanai starp DNS šablona reģionu un sintezēto RNS. Otrais, no Rho neatkarīgais, rodas pēc cilmes cilpas parādīšanās, ja aiz tās atrodas uracila bāzes komplekts. Šī kombinācija izraisa RNS atdalīšanu no DNS veidnes. Ir skaidrs, ka transkripcijas pārtraukšana ir fermentatīvs process, bet specifiski biokatalizatori tam vēl nav atrasti.
Vīrusu transkripcija
Vīrusu ķermeņiem nav savas proteīnu biosintēzes sistēmas, un tāpēc tie nevar vairoties, neizmantojot šūnas. Bet vīrusiem ir savs ģenētiskais materiāls, kas ir jārealizē un arī jāintegrē inficēto šūnu gēnos. Lai to izdarītu, viņiem ir vairāki enzīmi (vai izmanto šūnu enzīmu sistēmas), kas transkribē to nukleīnskābi. Tas ir, šis enzīms, pamatojoties uz vīrusa ģenētisko informāciju, sintezē ziņojuma RNS analogu. Bet tā nemaz nav RNS, bet gan DNS polimērs, kas papildina, piemēram, cilvēka gēnus.
Tas pilnībā pārkāpj tradicionālos transkripcijas principus bioloģijā, kā to var redzēt HIV vīrusa piemērā. Tās reversais enzīms no vīrusa RNS spēj sintezēt cilvēka nukleīnskābei komplementāru DNS. Papildu DNS sintezēšanas procesu no RNS sauc par reverso transkripciju. Tā ir bioloģijas definīcija procesam, kas ir atbildīgs par vīrusa iedzimtās informācijas integrāciju cilvēka genomā.
Vannu restaurācija Kolpino vk.com/restavraciya_vann_kolpino.
Transkripcija. Sākums - transkripcijas sākums, beigas - transkripcijas beigas, DNS - DNS.
Transkripcija ir RNS sintēzes process, izmantojot DNS kā veidni, un tas notiek visās dzīvajās šūnās. Citiem vārdiem sakot, tā ir ģenētiskās informācijas pārnešana no DNS uz RNS.
Transkripciju katalizē enzīma DNS atkarīgā RNS polimerāze. RNS sintēzes process notiek virzienā no 5" līdz 3" galam, tas ir, gar DNS šablona virkni, RNS polimerāze virzās virzienā 3"->5"
Transkripcija sastāv no iniciācijas, pagarināšanas un beigu posmiem.
Transkripcijas uzsākšana
Transkripcijas ierosināšana ir sarežģīts process, kas ir atkarīgs no DNS sekvences transkribētās sekvences tuvumā un no dažādu proteīna faktoru esamības vai neesamības.
Transkripcijas pagarinājums
Brīdis, kurā RNS polimerāze pāriet no transkripcijas ierosināšanas uz pagarinājumu, nav precīzi noteikts. Trīs galvenie bioķīmiski notikumi raksturo šo pāreju Escherichia coli RNS polimerāzes gadījumā: sigmas faktora izdalīšanās, pirmā fermenta molekulas pārvietošana gar šablonu un spēcīga transkripcijas kompleksa stabilizācija, kas papildus RNS. polimerāze ietver augošo RNS ķēdi un transkribēto DNS. Tādas pašas parādības ir raksturīgas arī eikariotu RNS polimerāzēm. Pāreju no iniciācijas uz pagarinājumu pavada saišu pārrāvums starp fermentu, promotoru, transkripcijas ierosināšanas faktoriem un dažos gadījumos RNS polimerāzes pāreja uz pagarinājuma kompetences stāvokli. Pagarinājuma fāze beidzas pēc augošā transkripta atbrīvošanas un fermenta atdalīšanās no veidnes.
Pagarinājuma stadijā DNS tiek savīti aptuveni 18 nukleotīdu pāri. Apmēram 12 DNS šablona virknes nukleotīdi veido hibrīda spirāli ar RNS virknes augošo galu. Kad RNS polimerāze pārvietojas pa veidni, DNS dubultspirāles attīšana notiek pirms tās, un DNS dubultspirāles atjaunošana notiek aiz tās. Tajā pašā laikā nākamā augošās RNS ķēdes saite tiek atbrīvota no kompleksa ar šablonu un RNS polimerāzi. Šīs kustības jāpavada RNS polimerāzes un DNS relatīvai rotācijai. Ir grūti iedomāties, kā tas varētu notikt šūnā, īpaši hromatīna transkripcijas laikā. Tāpēc ir iespējams, ka, lai novērstu šādu rotāciju, RNS polimerāzi, kas pārvietojas gar DNS, pavada topoizomerāzes.
Pagarināšana tiek veikta ar pagarinājuma pamatfaktoru palīdzību, kas nepieciešami, lai process priekšlaicīgi neapstātos.
Nesen ir parādījušies pierādījumi, ka regulējošie faktori var arī regulēt pagarinājumu. Pagarināšanas procesa laikā RNS polimerāze apstājas noteiktās gēna daļās. Tas ir īpaši skaidri redzams zemās substrātu koncentrācijās. Dažos matricas apgabalos ir ilgstoša RNS polimerāzes progresēšanas kavēšanās, tā sauktā. pauzes tiek novērotas pat pie optimālām substrāta koncentrācijām. Šo paužu ilgumu var kontrolēt ar pagarinājuma faktoriem.
| Triptofāna operons |
Transkripcija ir sintēzes processmolekulasRNS ieslēgtsapgabalāDNS, ko izmanto kā matricu. Transkripcijas nozīme ir ģenētiskās informācijas pārnešana no DNS uz RNS.
DNS molekula sastāv no divām komplementārām virknēm, savukārt RNS sastāv tikai no viena. Transkripcijas laikā tikai viena no DNS virknēm kalpo par RNS sintēzes veidni. Viņi viņu sauc semantiskā ķēde. Izņēmums ir mitohondriju DNS, kurā abas virknes ir jūtīgas un satur dažādus gēnus. Daži gēni ir ne tikai kodola DNS izņēmums, bet arī var būt lokalizēti nejēdzīgā virknē.
Transkripcijas laikā RNS molekula tiek sintezēta virzienā no 5" līdz 3" galam (kas ir dabiski visu nukleīnskābju sintēzei), savukārt pa DNS ķēdi sintēze notiek pretējā virzienā: 3"→5 ".
Eikariotos katrs gēns tiek transkribēts atsevišķi. Izņēmums atkal ir mitohondriju DNS, kas tiek pārrakstīta kopējā daudzgēnu transkriptā, kas pēc tam tiek sagriezts. Tā kā prokariotos gēni veido grupas, veidojot vienu operonu, šādi gēni tiek pārrakstīti kopā. Vienalga transkriptons sauc par DNS sadaļu, kas sastāv no promotora, transkribēta reģiona un terminatora.
Transkripcijā ir 3 posmi: iniciācija, pagarināšana, izbeigšana.
Iniciācija transkripcija ļauj sākt RNS molekulas sintēzi. Iniciācija ietver enzīmu kompleksa pievienošanu promotoram. Galvenā ir RNS polimerāze (šajā gadījumā atkarīga no DNS), kas, savukārt, sastāv no vairākām apakšvienības olbaltumvielām un spēlē procesa katalizatora lomu. Eikariotos transkripcijas uzsākšanu ietekmē īpašas DNS sadaļas: pastiprinātāji (stiprina) un klusinātāji (nomākti), kas parasti atrodas zināmā attālumā no paša gēna. Ir dažādi olbaltumvielu faktori, kas ietekmē transkripcijas ierosināšanas iespēju.
Prokariotiem ir tikai viena veida RNS polimerāze, savukārt eikariotiem ir trīs. RNS polimerāze 1 tiek izmantota trīs veidu ribosomu RNS sintezēšanai (pavisam ir 4 rRNS veidi). RNS polimerāzi 2 izmanto, lai sintezētu pre-mRNS (prekursoru messenger RNS). RNS polimerāze-3 sintezē vienu no ribosomu RNS veidiem, transporta un mazo kodolu.
RNS polimerāze spēj atpazīt specifiskas nukleotīdu sekvences un pievienoties tām. Šīs secības ir īsas un universālas visām dzīvajām būtnēm.
Pēc tam, kad RNS polimerāze saistās ar promotoru, DNS dubultās spirāles daļa atritinās un nukleotīdu saites starp šīs sadaļas pavedieniem tiek pārtrauktas. Tiek atšķetināti aptuveni 18 nukleotīdu pāri.
Uz skatuves pagarinājums secīga pievienošana notiek saskaņā ar brīvo nukleotīdu komplementaritātes principu atbrīvotajai DNS sadaļai. RNS polimerāze apvieno nukleotīdus poliribonukleotīdu ķēdē.
RNS sintēzes laikā aptuveni 12 tās nukleotīdi ir komplementāri un īslaicīgi saistīti ar DNS nukleotīdiem. Kad RNS polimerāze virzās tai priekšā, DNS ķēdes atdalās, un aiz tās ar enzīmu palīdzību tiek “sašūtas”. RNS ķēde pakāpeniski aug un iziet no RNS polimerāzes kompleksa.
Ir pagarinājuma faktori, kas novērš priekšlaicīgu transkripcijas pārtraukšanu.
Izbeigšana Transkripcijas process notiek terminatora reģionā, ko atpazīst RNS polimerāze, pateicoties īpašiem proteīna terminācijas faktoriem.
Daudzi adenīna nukleotīdi (poli-A) ir piesaistīti sintezētās RNS molekulas 3" galam, lai novērstu tās enzīmu sadalīšanos. Vēl agrāk, kad tika sintezēts 5" gals, t.s. vāciņš.
Vairumā gadījumu transkripcija nerada gatavu RNS. Neapstrādātai RNS joprojām ir jāiziet process apstrāde, kurā notiek tā modifikācijas izmaiņas un tas kļūst funkcionāli aktīvs. Katrs RNS veids eikariotos tiek pakļauts savām modifikācijām. Poli-A un vāciņa veidošanos bieži sauc arī par apstrādi.
Mēs arī iesakām
 Skaitliskā simbolika Jāņa Atklāsmes grāmatā
Skaitliskā simbolika Jāņa Atklāsmes grāmatā
 Apmācības nodarbība "Konfliktoloģija": Arābu-Izraēlas konflikts
Apmācības nodarbība "Konfliktoloģija": Arābu-Izraēlas konflikts
 Brīnums ar čūsku, kas notika ar svēto lielo mocekli Džordžu
Brīnums ar čūsku, kas notika ar svēto lielo mocekli Džordžu
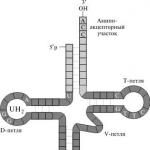 RNS saņem iedzimtu informāciju
RNS saņem iedzimtu informāciju
 Scaevola, kara ar etruskiem Gaja Muciusa varoņa goda vārds
Scaevola, kara ar etruskiem Gaja Muciusa varoņa goda vārds
 Kur ir predikāts? Kas ir "predikāts"? Predikātu veidi. Vai tā ir taisnība, ka vienkāršs verbāls predikāts ir predikāts, kura leksisko un gramatisko nozīmi izsaka viens darbības vārds
Kur ir predikāts? Kas ir "predikāts"? Predikātu veidi. Vai tā ir taisnība, ka vienkāršs verbāls predikāts ir predikāts, kura leksisko un gramatisko nozīmi izsaka viens darbības vārds