Mājasdarbs. Infusoria čības
Salīdzinot ar citām vienšūņu grupām, ciliātiem ir vissarežģītākā uzbūve, kas saistīta ar to funkciju daudzveidību un sarežģītību.
No kurienes cēlies nosaukums "infuzorijas tupele"? Jūs nebrīnīsieties, ja mikroskopā paskatīsities uz dzīvu skropstiņu vai pat uz tā attēlu (85. att.).
Patiešām, šī ciliāta ķermeņa forma atgādina elegantu dāmu apavu.
Infuzorijas apavi ir nepārtrauktā diezgan ātrā kustībā. Tā ātrums (istabas temperatūrā) ir aptuveni 2,0-2,5 mm/sek. Tas ir liels ātrums tik mazam dzīvniekam! Galu galā tas nozīmē, ka apavi sekundē noskrien distanci, kas 10-15 reizes pārsniedz ķermeņa garumu. Apavu trajektorija ir diezgan sarežģīta. Viņa virzās priekšgalā taisni uz priekšu
INFUSORIUM APAVI (PARAMECIUM CAUDATUM)
Lai iepazītos ar šo interesanto vienšūnu organismu uzbūvi un dzīvesveidu, vispirms pievērsīsimies vienam raksturīgam piemēram. Ņemsim kurpju ciliātus (Paramecium ģints sugas), kas ir plaši izplatītas seklās saldūdens rezervuāros. Šos ciliātus ir ļoti viegli pavairot nelielos akvārijos, ja dīķi piepilda ar parasto pļavas sienu. Šādās tinktūrās attīstās daudz un dažādi vienšūņu veidi, un gandrīz vienmēr attīstās kurpju ciliāti. Ar parastā izglītojošā mikroskopa palīdzību jūs varat redzēt daudz no tā, kas tiks apspriests tālāk.
Starp vienkāršākajiem ciliātu kurpes ir diezgan lieli organismi. Viņu ķermeņa garums ir aptuveni 1/6-1/3 mm. un griežas pa labi pa ķermeņa garenasi.
Šāda aktīva apavu kustība ir atkarīga no liela skaita smalkāko matiņa veida piedēkļu - skropstu, kas aptver visu skropstu ķermeni, darba. Skropstu skaits vienā no apavu ciliātēm ir 10-15 tūkstoši!
Katrs ciliums veic ļoti biežas lāpstiņām līdzīgas kustības – istabas temperatūrā līdz 30 sitieniem sekundē. Trieciena laikā ciliums tiek turēts iztaisnotā stāvoklī. Kad tas atgriežas sākotnējā stāvoklī (virzoties uz leju), tas pārvietojas 3-5 reizes lēnāk un apraksta pusloku.
Kad kurpes peld, tiek summētas daudzu tās ķermeni klājošo skropstu kustības. Atsevišķu skropstu darbības tiek koordinētas, kā rezultātā tiek radītas pareizas visu skropstu viļņveidīgas vibrācijas. Svārstību vilnis sākas ķermeņa priekšējā galā un izplatās atpakaļ. Tajā pašā laikā gar apavu korpusu iziet 2-3 kontrakcijas viļņi. Tādējādi viss skropstu ciliārais aparāts ir it kā vienots funkcionāls fizioloģisks veselums, kura atsevišķu struktūrvienību (ciliāru) darbības ir cieši saistītas (koordinētas) viena ar otru.
Katras atsevišķas apavu cilmes struktūra, kā liecina elektronu mikroskopiskie pētījumi, ir ļoti sarežģīta.
Apavu kustības virziens un ātrums nav nemainīgi un nemainīgi lielumi. Kurpe, tāpat kā visi dzīvie organismi (mēs to jau redzējām ar amēbas piemēru), reaģē uz izmaiņām ārējā vide kustības virziena maiņa.
Vienšūņu kustības virziena maiņu dažādu stimulu ietekmē sauc par taksometriem. Ciliātos ir viegli novērot dažādus taksometrus. Ja kurpes peld pilē, ievietojiet kādu vielu, kas tos negatīvi ietekmē (piemēram, kristālu galda sāls), tad apavi aizpeld (it kā bēgtu) no šī tiem nelabvēlīgā faktora (86. att.).

Pirms mums ir negatīvu taksometru piemērs ķīmiskai iedarbībai (negatīva ķemotaksi). Var novērot apavu un pozitīvu ķemotaksi. Ja, piemēram, ūdens lāse, kurā peld skropstas, tiek pārklāta ar vāka stiklu un zem tā tiek palaists oglekļa dioksīda (CO2) burbulis, tad lielākā daļa skropstu nokļūs šajā burbulī un apmetīsies ap to. gredzens.
Taksometru fenomens ļoti skaidri izpaužas apavos elektriskās strāvas ietekmē. Ja caur šķidrumu, kurā peld apavi, tiek izlaista vāja elektriskā strāva, var novērot šādu attēlu: visi skropstiņi orientē savu garenasi paralēli strāvas līnijai un pēc tam, it kā pēc komandas, virzās uz katodu, kuras apgabalā tie veido blīvu kopu. Skropstu kustību, ko nosaka elektriskās strāvas virziens, sauc par galvanotaksi. Visdažādāko vides faktoru ietekmē var konstatēt dažādus taksometrus ciliātos.
Viss skropstu citoplazmas ķermenis ir skaidri sadalīts 2 slāņos: ārējais ir gaišāks (ektoplazma) un iekšējais ir tumšāks un graudains (endoplazma).Svarīga loma skropstu ķermeņa formas noturības saglabāšanā.
Dzīva apavu korpusa ārējā slānī (ektoplazmā) skaidri redzami daudzi īsi stieņi, kas atrodas perpendikulāri virsmai (85., 7. att.). Šos veidojumus sauc par trihocistām. To funkcija ir ļoti interesanta un saistīta ar vienkāršāko aizsardzību. Ar mehānisku, ķīmisku vai kādu citu spēcīgu kairinājumu trihocistas tiek izmestas ar spēku, pārvēršoties par tieviem gariem pavedieniem, kas ietriecas apaviem uzbrūkošajam plēsējam. Trihocistas ir spēcīga aizsardzība. Tie ir regulāri izvietoti starp skropstiņām, lai trihocistu skaits aptuveni atbilstu skropstu skaitam. Lietoto (“šāvienu”) trihocistu vietā apavu ektoplazmā veidojas jaunas.
Vienā pusē, aptuveni ķermeņa vidū (85., 5. att.), apaviem ir diezgan dziļa ieplaka. Tas ir mutes dobums jeb peristoma. Cilia atrodas gar peristomas sienām, kā arī gar ķermeņa virsmu. Šeit tie ir izstrādāti daudz spēcīgāk nekā uz pārējās ķermeņa virsmas. Šīs cieši izvietotās skropstas ir sakārtotas divās grupās. Šo ļoti diferencēto skropstu funkcija ir saistīta nevis ar kustībām, bet ar uzturu (87. att.).

Kā un ko apavi ēd, kā tie sagremojas?
Apavi pieder pie ciliātiem, kuru galvenā barība ir baktērijas. Kopā ar baktērijām tās var norīt arī citas ūdenī suspendētas daļiņas neatkarīgi no to uzturvērtības. Periorālās skropstas rada nepārtrauktu ūdens plūsmu ar suspendētām daļiņām mutes atveres virzienā, kas atrodas dziļi peristomā. Mazas pārtikas daļiņas (visbiežāk baktērijas) caur muti iekļūst mazā cauruļveida rīklē un uzkrājas tās apakšā, uz robežas ar endoplazmu. Mutes atvere vienmēr ir atvērta. Varbūt nebūs kļūda teikt, ka skropstu kurpes ir viens no rijīgākajiem dzīvniekiem: tas nepārtraukti barojas. Šis process tiek pārtraukts tikai noteiktos dzīves brīžos, kas saistīti ar reprodukciju un dzimumprocesu.
Pēc tam rīkles apakšā uzkrātais barības kamols atraujas no rīkles dibena un kopā ar nelielu daudzumu šķidruma nonāk endoplazmā, veidojot gremošanas vakuolu. Pēdējais nepaliek savā veidošanās vietā, bet, iekrītot endoplazmas straumēs, veido diezgan sarežģītu un regulāru ceļu kurpes korpusā, ko sauc par gremošanas vakuola ciklozi (88. att.). Šī diezgan garā (istabas temperatūrā, apmēram stundu) gremošanas vakuola ceļojuma laikā tajā notiek vairākas izmaiņas, kas saistītas ar tajā esošās pārtikas gremošanu.
Šeit, tāpat kā amēbās un dažos flagellatos, notiek tipiska intracelulāra gremošana. No endoplazmas, kas ieskauj gremošanas vakuolu, tajā nonāk gremošanas enzīmi, kas iedarbojas uz pārtikas daļiņām. Gremošanas produkti tiek absorbēti caur gremošanas vakuolu endoplazmā.
Gremošanas vakuola ciklozes gaitā tajā tiek aizstātas vairākas gremošanas fāzes. Pirmajos brīžos pēc vakuola veidošanās to aizpildošais šķidrums maz atšķiras no apkārtējās vides šķidruma. Drīz no endoplazmas vakuolā sāk iekļūt gremošanas enzīmi, un tajā esošās vides reakcija kļūst krasi skāba. To ir viegli noteikt, ēdienam pievienojot kādu indikatoru, kura krāsa mainās atkarībā no apkārtējās vides reakcijas (skābā, neitrālā vai sārmainā). Šajā skābajā vidē notiek pirmās gremošanas fāzes. Tad attēls mainās, un reakcija gremošanas vakuolu iekšpusē kļūst nedaudz sārmaina. Šādos apstākļos ir nākamie soļi intracelulārā gremošana. Skābā fāze parasti ir īsāka par sārmu fāzi; tas ilgst aptuveni 1/6-1/4 no visa gremošanas vakuola uzturēšanās laika ciliāta ķermenī. Tomēr skābās un sārmainās fāzes attiecība var mainīties diezgan plašās robežās atkarībā no pārtikas veida.
Gremošanas vakuola ceļš endoplazmā beidzas ar to, ka tas tuvojas ķermeņa virsmai un caur pīlingu tiek izmests tās saturs, kas sastāv no šķidrām un nesagremotām pārtikas atliekām - notiek defekācija. Šis process atšķirībā no amēbām, kurās defekācija var notikt jebkur, apavos, tāpat kā citiem ciliātiem, ir stingri ierobežots noteiktā ķermeņa zonā, kas atrodas vēdera pusē (vēderu parasti sauc par dzīvnieka virsmu uz kura ir novietots periorālais padziļinājums), aptuveni pusceļā starp peristomu un ķermeņa aizmugurējo galu.
Tādējādi intracelulārā gremošana ir sarežģīts process, kas sastāv no vairākām fāzēm, kas pēc kārtas aizstāj viena otru.

Aprēķini liecina, ka apmēram 30-45 minūšu laikā no apaviem caur saraušanās vakuoliem izdalās šķidruma tilpums, kas vienāds ar ciliāta ķermeņa tilpumu. Tādējādi kontraktilo vakuolu darbības dēļ caur ciliāta ķermeni tiek veikta nepārtraukta ūdens plūsma, kas ieplūst no ārpuses caur mutes atveri (kopā ar gremošanas vakuoliem), kā arī osmotiski tieši caur pīlingu. Kontrakcijas vakuolām ir svarīga loma ūdens plūsmas regulēšanā, kas iet cauri ciliāta ķermenim, un osmotiskā spiediena regulēšanā. Šis process šeit notiek principā tāpat kā amēbās, tikai kontraktilās vakuolas struktūra ir daudz sarežģītāka.
Daudzus gadus zinātnieku vidū, kas pētīja vienšūņus, pastāvēja strīds par jautājumu, vai citoplazmā ir kādas struktūras, kas saistītas ar saraušanās vakuola parādīšanos, vai arī tā veidojas katru reizi no jauna. Uz dzīva ciliāta nevar novērot nekādas īpašas struktūras, kas būtu pirms tās veidošanās. Pēc vakuolas - sistoles - kontrakcijas citoplazmā bijušās vakuolas vietā nav redzamas absolūti nekādas struktūras. Pēc tam atkal parādās caurspīdīgs burbulis vai adduktora kanāli, kas sāk palielināties. Tomēr nav atrasta saikne starp jaunizveidoto vakuolu un iepriekš esošo vakuolu. Šķiet, ka nav nepārtrauktības starp secīgiem kontraktilās vakuolas cikliem, un citoplazmā no jauna veidojas jauna kontrakta vakuola. Tomēr īpašas pētījumu metodes ir pierādījušas, ka patiesībā tas tā nav. Elektronu mikroskopijas izmantošana, kas dod ļoti lielu palielinājumu (līdz 100 tūkstošiem reižu), pārliecinoši parādīja, ka ciliātam ir īpaši diferencēta citoplazma apgabalā, kur veidojas kontraktilie vakuoli, kas sastāv no plānāko caurulīšu savijas. Tādējādi izrādījās, ka saraušanās vakuola citoplazmā nerodas uz " tukša vieta”, bet uz iepriekšējā speciālā šūnu organoīda bāzes, kura funkcija ir saraušanās vakuola veidošanās.
Tāpat kā visiem vienšūņiem, arī ciliātiem ir šūnu kodols. Tomēr kodolaparatūras uzbūves ziņā ciliāti krasi atšķiras no visām pārējām vienšūņu grupām.
Ciliātu kodolaparātu raksturo tā duālisms. Tas nozīmē, ka ciliātiem ir divu dažādu veidu kodoli – lielie kodoli jeb makrokodoli un mazie kodoli jeb mikrokodoli. Apskatīsim, kāda kodolaparāta uzbūve ir kurpes ciliātos (85. att.).
Ciliāta ķermeņa centrā (peristomas līmenī) atrodas liels masīvs olveida vai pupiņas formas kodols. Tas ir makrokodolis. Tās tiešā tuvumā atrodas otrs daudzkārt mazāks kodols, parasti diezgan cieši blakus makrokodolam. Tas ir mikrokodolis. Atšķirība starp šiem diviem kodoliem ir ne tikai izmērā, tā ir daudz būtiskāka, dziļi ietekmējot to struktūru.
Makrokodolis, salīdzinot ar mikrokodolu, ir daudz bagātāks ar īpašu kodolvielu (hromatīnu vai, precīzāk, dezoksiribonukleīnskābi, saīsināti DNS), kas ir daļa no hromosomām.
Nesenie pētījumi ir parādījuši, ka makrokodolu ir vairākas desmit (un dažos skropstas un simtiem) reižu liels skaits hromosomas nekā mikrokodoli. Makrokodolis ir ļoti savdabīgs daudzhromosomu (poliploīdu) kodolu veids. Tādējādi atšķirība starp mikro- un makrokodoliem ietekmē to hromosomu sastāvu, kas nosaka to kodolvielas - hromatīna - lielāku vai mazāku bagātību.
Vienā no visizplatītākajiem ciliātu veidiem - kurpes(Paramecium caudatum) - ir viens makrokodolu (saīsināti Ma) un viens mikrokodolu (saīsināti Mi). Šāda kodolaparatūras struktūra ir raksturīga daudziem ciliātiem. Citiem var būt vairāki Ma un Mi. Bet visiem ciliātiem raksturīga iezīme ir kodolu diferenciācija divās kvalitatīvi atšķirīgās grupās — Ma un Mi jeb, citiem vārdiem sakot, kodolduālisma fenomens.
Kā vairojas ciliāti? Kā piemēru atkal pievērsīsimies infuzorijas apaviem. Ja jūs iestādīsit vienu apavu eksemplāru nelielā traukā (mikroakvārijā), tad dienā būs divi un bieži četri ciliāti. Kā tas notiek? Pēc noteikta aktīvās peldēšanas un barošanas perioda skropstas ir nedaudz izstieptas garumā. Tad tieši ķermeņa vidū parādās arvien dziļāks šķērseniskais sašaurinājums (90. att.). Beigās skropstas it kā tiek sašņorētas uz pusēm, un no viena indivīda tiek iegūti divi īpatņi, kas sākotnēji ir nedaudz mazāki par vecāku indivīdu. Viss skaldīšanas process istabas temperatūrā aizņem apmēram stundu. Iekšējo procesu izpēte liecina, ka jau pirms šķērseniskās sašaurinājuma parādīšanās sākas kodolaparāta skaldīšanas process. Vispirms tiek koplietots Mi un tikai pēc tam Ma. Mēs šeit neapstāsimies detalizēta izskatīšana kodola dalīšanās procesiem un tikai atzīmējam, ka Mi dalās ar mitozi, savukārt Ma dalīšanās pēc izskata atgādina tiešu kodola skaldīšanu – amitozi. Šis apavu ciliātu aseksuālās vairošanās process, kā mēs redzam, ir līdzīgs amēbu un flagellātu aseksuālai pavairošanai. Turpretim ciliāti aseksuālās vairošanās procesā vienmēr sadalās šķērsām, savukārt kauliņiem dalījuma plakne ir paralēla ķermeņa gareniskajai asij.
Sadalīšanās laikā notiek dziļa iekšēja ciliāta ķermeņa pārstrukturēšana. Tiek veidotas divas jaunas peristomas, divas rīkles un divas mutes atveres. Tajā pašā laikā tiek ieplānots skropstu bazālo kodolu sadalījums, kā rezultātā veidojas jaunas skropstas. Ja vairošanās laikā skropstu skaits nepalielinātos, tad katras dalīšanās rezultātā meitas īpatņi saņemtu apmēram pusi no mātes īpatņa skropstu skaita, kas novestu pie pilnīgas skropstu „plikās”. Patiesībā tas nenotiek.

Laiku pa laikam lielākajai daļai skropstu, arī apaviem, ir īpaša un ārkārtīgi savdabīga dzimumprocesa forma, ko sauc par konjugāciju. Mēs šeit neanalizēsim detalizēti visas sarežģītās kodolizmaiņas, kas pavada šo procesu, bet atzīmēsim tikai vissvarīgākās. Konjugācija notiek šādi (91. att.) Divi skropstiņi tuvojas viens otram, cieši pieliek viens otram ar ventrālajām pusēm, un šādā formā tie diezgan peld. ilgu laiku kopā (pie kurpes apmēram 12 stundas istabas temperatūrā). Pēc tam konjuganti atdalās. Kas notiek ciliātu ķermenī konjugācijas laikā? Šo procesu būtība ir šāda (91. att.). Lielais kodols (makrokodolis) sabrūk un pakāpeniski izšķīst citoplazmā. Mikrokodoli vispirms sadalās, daļa dalīšanās rezultātā izveidojušos kodolu tiek iznīcināti (sk. 91. att.). Katrs no konjugantiem saglabā divus kodolus. Viens no šiem kodoliem paliek vietā indivīdā, kurā tas tika izveidots (stacionārais kodols), bet otrs aktīvi pārvietojas konjugācijas partnerā (migrējošajā kodolā) un saplūst ar savu stacionāro kodolu. Tādējādi katrā no konjugantiem šajā stadijā ir viens kodols, kas veidojas stacionāro un migrējošo kodolu saplūšanas rezultātā. Šo sarežģīto kodolu sauc par sinkarionu. Sinkariona veidošanās nav nekas vairāk kā apaugļošanās process. Un daudzšūnu organismos būtisks apaugļošanas brīdis ir dzimumšūnu kodolu saplūšana. Ciliātos dzimumšūnas neveidojas, ir tikai dzimumkodoli, kas saplūst savā starpā. Tādējādi notiek savstarpēja savstarpēja apaugļošanās.
Neilgi pēc sinkariona veidošanās konjuganti atdalās. Saskaņā ar to kodolaparatūras struktūru šajā posmā tie joprojām ļoti būtiski atšķiras no parastajiem tā sauktajiem neitrālajiem (nekonjugējošiem) ciliātiem, jo tiem katram ir tikai viens kodols. Nākotnē sinkariona dēļ tiek atjaunots normāls kodolaparāts. Sinkarions ir sadalīts (vienu vai vairākas reizes). Daļa no šīs nodaļas produktiem sarežģītu transformāciju rezultātā, kas saistītas ar hromosomu skaita palielināšanos un hromatīna bagātināšanu, pārvēršas makrokodolos. Citi saglabā mikrokodolu raksturīgo struktūru. Tādā veidā tiek atjaunots ciliātiem raksturīgais un tipiskais kodola aparāts, pēc kura skropstiņiem sākas bezdzimuma vairošanās ar skaldīšanu.
Tādējādi konjugācijas process ietver divus būtiskus bioloģiskos momentus: apaugļošanu un jauna makrokodola atjaunošanos sinkariona dēļ.
Kāda ir konjugācijas bioloģiskā nozīme, kāda loma tai ir ciliātu dzīvē? Mēs to nevaram saukt par vairošanos, jo īpatņu skaita pieauguma nav. Iepriekš minētie jautājumi ir kalpojuši par materiālu daudziem eksperimentāliem pētījumiem, kas veikti daudzās valstīs. Šo pētījumu galvenais rezultāts ir šāds. Pirmkārt, konjugācija, tāpat kā jebkurš cits seksuāls process, kurā vienā organismā apvienojas divi iedzimtie principi (tēva un mātes), izraisa iedzimtas mainības, iedzimtas daudzveidības palielināšanos. Iedzimtās mainīguma palielināšanās palielina organisma adaptācijas spējas vides apstākļiem. Otra bioloģiski svarīgā konjugācijas puse ir jauna makrokodola veidošanās sinkariona skaldīšanas produktu dēļ un tajā pašā laikā vecā iznīcināšana. Eksperimentālie dati liecina, ka tieši makrokodolum ir ārkārtīgi liela nozīme skropstu dzīvē. Tas kontrolē visus galvenos dzīvības procesus un nosaka svarīgāko no tiem - proteīna veidošanos (sintēzi), kas veido dzīvas šūnas protoplazmas galveno daļu. Ar ilgstošu aseksuālu vairošanos dalīšanās ceļā notiek savdabīgs makrokodola un vienlaikus visas šūnas “novecošanās” process: vielmaiņas procesa aktivitāte samazinās, sadalīšanās ātrums samazinās. Pēc konjugācijas (kuras laikā, kā redzējām, vecais makrokodols tiek iznīcināts), tiek atjaunots vielmaiņas līmenis un dalīšanās ātrums. Tā kā apaugļošanās process notiek konjugācijas laikā, kas vairumā citu organismu ir saistīts ar vairošanos un jaunas paaudzes parādīšanos, tad ciliātos pēc konjugācijas izveidojušos indivīdu var uzskatīt arī par jaunu dzimumpaaudzi, kas šeit rodas, jo bija, pateicoties veco “atjaunošanai”.
Par piemēru skropstu apaviem mēs tikāmies ar tipisku plašas ciliātu klases pārstāvi. Tomēr šai klasei raksturīga ārkārtēja sugu daudzveidība gan struktūras, gan dzīvesveida ziņā. Sīkāk apskatīsim dažas no raksturīgākajām un interesantākajām formām.
Ciliātiem apavu skropstas vienmērīgi pārklāj visu ķermeņa virsmu. Tā ir raksturīga struktūras iezīme (Holotrich). Daudziem ciliātiem ir raksturīgs atšķirīgs ciliārā apvalka attīstības raksturs. Fakts ir tāds, ka ciliātu skropstas, apvienojot kopā, spēj veidot sarežģītākus kompleksus. Piemēram, bieži tiek novērots, ka skropstas, kas atrodas vienā vai divās rindās tuvu viena otrai, savienojas (salīp kopā), veidojot plāksni, kas, tāpat kā skropstas, spēj pārspēt. Šādus slāņveida saraušanās veidojumus sauc par membrānām (ja tie ir īsi) vai membrānām (ja tie ir gari). Citos gadījumos skropstas ir savienotas kopā, atrodas ciešā saišķī. Šie veidojumi - cirres - atgādina otu, kuras atsevišķie matiņi ir salipuši kopā. Daudziem ciliāriem ir raksturīgi dažādi sarežģīti ciliāru veidojumi. Ļoti bieži ciliārais apvalks neattīstās vienmērīgi, bet tikai atsevišķās ķermeņa daļās.
INFUSORIA trompetists (STENTOR POLYMORPHIC)
Saldūdeņos pieder lielu skaistu ciliātu sugas sava veida trompetisti(Stentors). Šis nosaukums diezgan atbilst šo dzīvnieku ķermeņa formai, kas patiešām atgādina cauruli (92. att.), kas vienā galā plaši atvērta. Pirmajā iepazīšanās reizē ar dzīvajiem trompetistiem var pamanīt vienu apaviem neraksturīgu iezīmi. Pie mazākā kairinājuma, arī mehāniska (piemēram, uzsitot ar zīmuli pa stiklu, kur ir ūdens lāse ar taurētājiem), viņu ķermenis strauji un ļoti ātri (sekundes daļā) saraujas, iegūstot gandrīz regulāru. sfēriska forma. Tad diezgan lēni (laiku mēra sekundēs) trompetists iztaisnojas, iegūstot sev raksturīgo formu. Šī trompetista spēja ātri sarauties ir saistīta ar īpašu muskuļu šķiedru klātbūtni, kas atrodas gar ķermeni un ektoplazmā. Tādējādi muskuļu sistēma var veidoties arī vienšūnu organismā.

Trompetistu ģintī ir sugas, no kurām dažām ir raksturīga diezgan spilgta krāsa. Ļoti bieži sastopams saldūdeņos zilais trompetists(Stentor coeruleus), kas ir spilgti zils. Šāds trompetista krāsojums ir saistīts ar faktu, ka tā ektoplazmā atrodas mazākie zilā pigmenta graudi.
Cita trompetistu suga (Stentor polymorphus) bieži ir zaļā krāsā. Šīs krāsas iemesls ir diezgan atšķirīgs. Zaļā krāsa sakarā ar to, ka skropstas endoplazmā dzīvo un vairojas mazas vienšūnas zaļaļģes, kas trompetista ķermenim piešķir raksturīgu krāsu. Stentor polymorphus ir tipisks abpusēji izdevīgas kopdzīves - simbiozes piemērs. Trompetists un aļģe ir savstarpēji simbiotiskās attiecībās: trompetists aizsargā savā organismā mītošās aļģes un apgādā tās ar elpošanas rezultātā radušos oglekļa dioksīdu; no savas puses aļģes nodrošina trompetistu ar skābekli, kas izdalās fotosintēzes procesā. Acīmredzot daļu aļģu sagremo skropstas, kas ir barība trompetistam.
Trompetisti ūdenī peld lēni ar plato galu uz priekšu. Bet tos var arī īslaicīgi piestiprināt pie pamatnes ar korpusa aizmugurējo šauru galu, uz kura veidojas mazs piesūceknis.
Trompetista korpusā var atšķirt stumbra apvidu, kas izplešas no aizmugures uz priekšu, un plašu periorālo (peristomālo) lauku, kas atrodas gandrīz perpendikulāri tam. Šis lauks atgādina asimetrisku plakanu piltuvi, kuras vienā malā ir padziļinājums - rīkle, kas ved uz ciliāta endoplazmu. Trompetista ķermenis ir klāts ar gareniskām īsu skropstu rindām. Gar peristomālā lauka malu aplī atrodas spēcīgi attīstīta gandrīz orāla (adorāla) membrānas zona (92. att.). Šo zonu veido liels skaits atsevišķu ciliāru plākšņu, no kurām katra, savukārt, sastāv no daudzām viena ar otru salipušām cilijām, kas atrodas divās cieši izvietotās rindās.
Mutes atveres rajonā periorālās membrānas ir ietītas pret rīkli, veidojot kreisās puses spirāli. Ūdens plūsma, ko izraisa periorālās membrānas svārstības, tiek virzīta uz mutes atveri (ķermeņa priekšējā gala veidotās piltuves dziļumā). Kopā ar ūdeni rīklē nonāk arī ūdenī suspendētas pārtikas daļiņas. Trompetista pārtikas priekšmeti ir daudzveidīgāki nekā čībām. Kopā ar baktērijām tas ēd mazus vienšūņus (piemēram, flagellates), vienšūnas aļģes utt.
Trompetistam ir labi attīstīta saraušanās vakuole, kurai ir savdabīga uzbūve. Centrālais rezervuārs atrodas ķermeņa priekšējā trešdaļā, nedaudz zem mutes atveres. No tā atiet divi gari pievienojošie kanāli. Viens no tiem iet no rezervuāra uz ķermeņa aizmugurējo galu, otrs atrodas peristomālā lauka reģionā paralēli membrānas periorālajai zonai.
Infuzorijas trompetists ir iecienīts reģenerācijas eksperimentālo pētījumu objekts. Daudzi eksperimenti ir pierādījuši trompetistu augsto atjaunošanās spēju. Ciliātu ar plānu skalpeli var sagriezt daudzās daļās, un katru no tām cauri īsu laiku(vairākas stundas, reizēm dienā un vairāk) pārvērtīsies par proporcionāli uzbūvētu, bet mazu trompetistu, kas tad enerģētiskās barošanās rezultātā sasniedz šai sugai raksturīgo izmēru. Lai pabeigtu atveseļošanās procesus, reģenerējošajam gabalam jābūt vismaz vienam pērlveida makrokodola segmentam.
Trompetistam, kā mēs redzējām, ir dažādas skropstas: no vienas puses, tās ir īsas, aptverot visu ķermeni, un, no otras puses, ir periorāla membrānas zona. Saskaņā ar šo raksturīgo struktūras iezīmi tika nosaukta ciliātu atdalīšanās, pie kuras pieder trompetists. skropstu ciliāti(Heterotricha).
INFUSORIA BURSARIA (BURSARIA TRUNCATELLA)
Otrs interesantais ciliāru ciliātu pārstāvis bieži sastopams saldūdeņos bursārija(Bursaria truncatella, 93. att.). Tas ir milzis starp ciliātiem: tā izmēri var sasniegt 2 mm, visizplatītākie izmēri ir 0,5-1,0 mm. Bursaria ir skaidri redzama ar neapbruņotu aci. Saskaņā ar nosaukumu bursārijai ir maisiņa forma, kas ir plaši atvērta priekšējā galā (bursa ir latīņu vārds, tulkojumā krievu valodā nozīmē “somiņa”, “soma”) un nedaudz paplašināta aizmugurējā galā. Viss skropstas ķermenis ir klāts ar gareniskām īsu skropstu rindām. Viņu sišana izraisa diezgan lēnu dzīvnieka kustību uz priekšu. Bursārija peld tā, it kā "ripotos" no vienas puses uz otru.

No priekšējā gala ķermeņa dziļumos (apmēram 2/3 no tā garuma) izvirzās periorāla ieplaka - peristoma. Ventrālajā pusē tas sazinās ar ārējo vidi caur šauru spraugu, muguras pusē peristomas dobums nesazinās ar ārējo vidi. Ja paskatās uz bursārijas ķermeņa augšējās trešdaļas šķērsgriezumu (93. att., B), var redzēt, ka peristomas dobums aizņem lielāko daļu ķermeņa, savukārt citoplazma to ieskauj loka formā.
Ķermeņa priekšējā galā pa kreisi ļoti spēcīgi attīstīta periorālās (adorālās) membrānas zona (93., 4. att.) rodas bursārijā. Tas nolaižas peristomas dobuma dziļumā, pagriežoties pa kreisi. Adorālā zona sniedzas līdz peristomas dziļākajai daļai. Citu ciliāru veidojumu peristomālajā dobumā, izņemot periorālās membrānas, nav, izņemot ciliāru joslu, kas iet gar peristomas dobuma ventrālo pusi (93., 10. att.). Iekšpusē aizmugurējā siena peristomālajā dobumā gandrīz visā garumā ir šaurs šķēlums (93., 7. att.), kura malas parasti cieši piekļaujas viena otrai. Šī ir mutes atvēršana. Tās malas atdalās tikai ēšanas laikā.
Bursārijiem nav šauras pārtikas specializācijas, taču tie galvenokārt ir plēsēji. Virzoties uz priekšu, viņi sastopas ar dažādiem maziem dzīvniekiem. Pateicoties periorālās zonas membrānu darbam, upuris ar spēku tiek ievilkts plašajā peristomālajā dobumā, no kurienes tas vairs nevar izpeldēt. Pārtikas priekšmeti tiek nospiesti pret peristomālā dobuma muguras sieniņu un caur paplašināto mutes plaisu iekļūst endoplazmā. Bursārijas ir ļoti rijīgas, tās var norīt diezgan lielus priekšmetus: piemēram, viņu iecienītākais ēdiens ir apavu ciliāti. Bursaria spēj norīt 6-7 apavus pēc kārtas. Tā rezultātā bursārijas endoplazmā veidojas ļoti lieli gremošanas vakuoli.
Bursārijas kodolaparāts ir diezgan sarežģīts. Viņiem ir viens garš desas formas makrokodolis un liels (līdz aptuveni 30) mazu mikrokodolu skaits, kas nejauši izkaisīti ciliāta endoplazmā.
Bursaria ir viena no nedaudzajām saldūdens skropstu sugām, kurām trūkst saraušanās vakuolu. Kā šajā lielajā ciliātā tiek veikta osmoregulācija, joprojām nav pilnībā skaidrs. Zem bursārijas ektoplazmas dažādās ķermeņa daļās var novērot dažādu formu un izmēru šķidruma burbuļus - vakuolus, kas maina to tilpumu. Acīmredzot šīs neregulāra forma vakuolām un pēc savas funkcijas atbilst citu ciliātu saraušanās vakuoliem.

Interesanti ir novērot secīgos Bursaria aseksuālās vairošanās posmus. Sākotnējās dalīšanās stadijās tiek pilnībā samazināts viss peristomas dobums un membrānas gandrīz orālā zona (94. att.). Ir saglabāts tikai ārējais cilijs. Infuzorija ir olas formā. Pēc tam korpuss ir sašņorēts ar šķērsenisku rievu divās daļās. Katrā no iegūtajiem meitas ciliātiem diezgan sarežģītu transformāciju rezultātā veidojas tipiska membrānas peristoma un periorālā zona. Viss bursārijas dalīšanas process norit ātri un aizņem nedaudz vairāk par stundu.
Bursārijā ir ļoti viegli novērot vēl vienu svarīgu dzīvības procesu, kura sākums ir saistīts ar ciliātiem nelabvēlīgiem apstākļiem, cistu veidošanās procesu (encystation). Šī parādība ir raksturīga, piemēram, amēbai. Bet izrādās, ka pat tādi sarežģīti organizēti vienšūņi kā ciliāti spēj pāriet neaktīvā stāvoklī. Ja kultūra, kurā dzīvo bursāri, netiek laikus pabarota vai atdzesēta, tad pēc dažām stundām sāksies masveida enstacija. Šis process notiek šādi. Bursarīdi, kā arī pirms dalīšanās zaudē membrānu peristomu un periorālo zonu. Tad tie kļūst pilnīgi sfēriski, pēc tam tie izšķir raksturīgās formas dubultu apvalku (94. att., D).
Cistu stāvoklī bursārija var būt vairākus mēnešus. Kad rodas labvēlīgi apstākļi, cistas apvalks pārsprāgst, no tā izplūst bursārija, attīstās peristoma un pāriet uz aktīvu dzīvi.
STYLONICHIA MYTILUS
Ļoti sarežģītam un dažādi diferencētam ciliāram aparātam ir ciliāti, kas saistīti ar kuņģa-zarnu trakta atdalīšanās(Hypotricha), kuras daudzas sugas dzīvo gan saldūdenī, gan jūras ūdenī. Viens no visizplatītākajiem, biežāk sastopamajiem pārstāvjiem interesanta grupa var saukt stilonihija(Stylonichia mytilus). Šis ir diezgan liels ciliāts (garums līdz 0,3 mm), kas dzīvo saldūdens rezervuāru dibenā, ūdens veģetācija(95. att.). Atšķirībā no čības, trompetista un bursārijas, stilonihijai trūkst nepārtraukta ciliāra apvalka, un visu ciliāru aparātu pārstāv ierobežots skaits stingri noteiktu ciliāru veidojumu.

Stilonychia ķermenis (tāpat kā lielākajai daļai citu vēdera skropstu ciliātu) ir stipri saplacināts dorso-abdominālā virzienā, un tā dorsālās un vēdera puses, priekšējie un aizmugurējie gali ir skaidri atšķirami. Ķermenis ir nedaudz paplašināts no priekšpuses, sašaurināts aizmugurē. Apskatot dzīvnieku no ventrālās puses, skaidri redzams, ka priekšējā trešdaļā pa kreisi ir sarežģīti izkārtota pinnāta un mutes atvere.
Muguras pusē diezgan reti atrodas skropstas, kuras nav spējīgas pārspēt. Tos drīzāk var saukt par plāniem elastīgiem sariem. Tie ir nekustīgi un tiem nav nekā kopīga ar kustības funkciju. Šīm cilijām parasti tiek piešķirta taustes, jutīga funkcija.
Visi ciliāri veidojumi, kas saistīti ar kustību un barības uztveršanu, koncentrējas dzīvnieka ventrālajā pusē (95. att.). Ir neliels skaits resnu pirkstveida veidojumu, kas izvietoti vairākās grupās. Tās ir vēdera cirras. Katrs no tiem ir sarežģīts ciliārs veidojums, kas ir daudzu desmitu atsevišķu ciliāru ciešas saiknes (salipšanas) rezultāts. Tādējādi cirres ir kā otas, kuru atsevišķie matiņi ir cieši savesti kopā un savienoti viens ar otru.
Ar cirozes palīdzību dzīvnieks diezgan ātri pārvietojas, “skraida” pa substrātu. Papildus "rāpošanai" un "skriešanai" pa pamatni, stilonychia spēj radīt diezgan asus un spēcīgus lēcienus, nekavējoties atraujoties no substrāta. Šīs asās kustības tiek veiktas ar divu spēcīgu astes cirpu palīdzību (95. att.), kas nepiedalās parastajā "rāpošanā".
Divas marginālo (marginālo) cireju rindas atrodas gar ķermeņa malu labajā un kreisajā pusē. No dzīvnieka labās malas tie virzās pa visu ķermeni, savukārt no kreisās malas sasniedz tikai peristomas reģionu. Šie ciliārie veidojumi kalpo dzīvnieka dzīšanai, kad tas ir atdalīts no substrāta un brīvi peld ūdenī.
Tāpēc mēs redzam, ka daudzveidīgais un specializētais stilonihijas ciliārais aparāts ļauj tai veikt ļoti dažādas kustības, atšķirībā no, piemēram, vienkāršas slīdēšanas ūdenī, piemēram, kurpes vai trompetists.
Arī ciliārais aparāts, kas saistīts ar uztura funkciju, ir sarežģīts. Mēs jau redzējām, ka gandrīz mutes dobums (peristome), kura apakšā atrodas mutes atvere, kas ved uz rīkli, atrodas dzīvnieka priekšējā pusē kreisajā pusē. Gar kreiso malu, sākot no paša ķermeņa priekšējā gala, ir stipri attīstīta periorālo (adorālo) membranoļu zona. Ar savu sitienu tie virza ūdens plūsmu uz mutes atveri. Turklāt peristomālā padziļinājuma rajonā ir vēl trīs kontraktilās membrānas (membrānas), kas ar iekšējiem galiem iestiepjas rīklē, un vairākas īpašas periorālas cilijas (95. att.). Viss šis kompleksais aparāts kalpo ēdiena uztveršanai un virzīšanai mutes atverē.
Stilonychia ir viens no vienšūņiem ar ļoti plašu barības objektu klāstu. To var pamatoti saukt par visēdāju. Viņa var ēst baktērijas, piemēram, kurpes. Starp tās pārtikas objektiem ir flagellates, vienšūnu aļģes (bieži vien kramaļģes). Visbeidzot, Stilonychia var būt arī plēsējs, kas uzbrūk citām, mazākām skropstu sugām un aprij tās.
Stilonychia ir saraušanās vakuola. Tas sastāv no centrālā rezervuāra, kas atrodas peristomas kreisajā aizmugurējā galā, un viena adduktora kanāla, kas vērsts atpakaļ.
Kodolaparāts, kā vienmēr ciliātiem, sastāv no makrokodola un mikrokodola.
Makrokodolu veido divas puses, kas savienotas ar plānu sašaurinājumu; ir divi mikrokodoli, tie atrodas tieši pie abām Ma pusēm.
Stilonychia, daļēji bursaria, trompeteris - tie visi ir ciliāti ar plašu pārtikas produktu klāstu. Spēja uzņemt dažādus pārtikas produktus ir raksturīga lielākajai daļai ciliātu. Tomēr starp tām var atrast arī tādas sugas, kas ir stingri specializētas attiecībā uz pārtikas dabu.
INFŪZIJAS PLĒSĒJI
Starp skropstiņiem ir plēsēji, kas ir ļoti "izvēlīgi" pret savu upuri. Labs piemērs ir infuzorija. didīnija(Didinium nasutum). Didīnijs ir salīdzinoši mazs ciliāts, kura garums ir vidēji 0,1–0,15 mm. Priekšējais gals ir izstiepts proboscis formā, kura galā novietota mutes atvere. Ciliārais aparāts ir attēlots ar diviem ciliāru vainagiem (96. att.). Didinius ātri peld ūdenī, bieži mainot virzienu. Vēlamais didīniju ēdiens ir apavu ciliāti. Šajā gadījumā plēsējs ir mazāks par savu upuri. Didinius ar stumbru iekļūst laupī un pēc tam, pakāpeniski paplašinot muti arvien vairāk, norij apavu veselu! Proboscī ir īpašs, tā sauktais stienis, aparāts. Tas sastāv no vairākām elastīgām stiprām nūjām, kas atrodas citoplazmā gar proboscis perifēriju. Tiek uzskatīts, ka šis aparāts palielina spārnu sieniņu izturību, kas nelūzt, norijot tik milzīgu laupījumu, salīdzinot ar didīniju, kā kurpes. Didinius - piemērs pēdējais līdzeklis plēsoņa vienšūņu vidū. Ja salīdzinām didīniju, kas norij savu laupījumu - apavus - ar plēsoņām augstākiem dzīvniekiem, tad līdzīgus piemērus atrast ir grūti.

Didinius, norijot parameciju, protams, ļoti uzbriest. Gremošanas process ir ļoti ātrs, istabas temperatūrā tas aizņem tikai aptuveni divas stundas. Tad nesagremotās atliekas tiek izmestas ārā, un didīnijs sāk medīt citu upuri. Īpašos pētījumos noskaidrots, ka didīnijas ikdienas "diēta" ir 12 apavi - patiesi kolosāla apetīte! Jāpatur prātā, ka intervālos starp nākamajām "medībām" didīnijas dažreiz sadalās. Pietrūkstot ēdienam, didīnija ļoti viegli enzija un tikpat viegli atkal izkļūst no cistām.
zālēdāju ciliāti
Daudz retāk nekā plēsonis, tas ir sastopams starp skropstiņiem "tīrais veģetārisms" - ēd tikai augu barība. Viens no nedaudzajiem "veģetāro" ciliātu piemēriem var būt pārstāvji ģints passula(Nassula). Viņu barības objekts ir pavedienveida zilaļģes (97. att.).

Tie iekļūst endoplazmā caur muti, kas atrodas sānos, un pēc tam savērpj ciliātus stingrā spirālē, kas pakāpeniski tiek sagremota. Aļģu pigmenti daļēji iekļūst skropstu citoplazmā un nokrāso to spilgti tumši zaļā krāsā.
SUVOIKA (VORTICELLA NEBULIFERA)
Interesanta un sugu skaita ziņā diezgan liela skropstu grupa ir substrātam piestiprinātas sēdošās formas, veidojot apaļo acu atdalīšanās(Peritricha). Visizplatītākie šīs grupas pārstāvji ir suvoyki(Vorticella ģints sugas).
Suvoyki atgādina elegantu ziedu kā zvaniņu vai maijpuķīti, kas sēž uz gara kāta, kas ar galu piestiprināts pie substrāta. Lielāko dzīves daļu suvojka pavada stāvoklī, kas piestiprināts pie substrāta.

Apsveriet skropstu ķermeņa uzbūvi. Dažādām sugām to izmēri atšķiras diezgan plašā diapazonā (līdz aptuveni 150 mikroniem). Mutes disks (98. att.) atrodas uz paplašinātas ķermeņa priekšējās daļas, kurā pilnībā nav skropstu. Ciliārais aparāts atrodas tikai gar mutes (peristomālā) diska malu (98. att.) īpašā rievā, ārpus kuras veidojas veltnis (peristomālā lūpa). Gar veltņa malu iet trīs ciliētas membrānas, no kurām divas atrodas vertikāli, viena (ārējā) ir horizontāla. Tie veido nedaudz vairāk nekā vienu pilnu spirāles apgriezienu. Šīs membrānas atrodas pastāvīgā mirgojošā kustībā, virzot ūdens plūsmu uz mutes atveri. Mutes aparāts sākas diezgan dziļi ar piltuvi peristomālā lauka malā (98. att.), kuras dziļumā ir mutes atvere, kas ved uz īsu rīkli. Suvoys, tāpat kā kurpes, barojas ar baktērijām. Viņu mutes atvērums pastāvīgi ir atvērts, un mutē nepārtraukti plūst ūdens.
Viena saraušanās vakuole bez adduktora kanāliem atrodas netālu no mutes atveres. Makrokodolumam ir lentai vai desai līdzīga forma, cieši blakus tam ir viens mazs mikrokodols.
Suvoyka spēj strauji saīsināt kātu, kas sekundes daļā tiek savīti ar korķviļķi. Tajā pašā laikā ciliāta ķermenis arī saraujas: peristomālais disks un membrānas tiek ievilktas uz iekšu, un viss priekšējais gals aizveras.

Protams, rodas jautājums: tā kā suvoys ir piestiprinātas pie pamatnes, kādā veidā tiek veikta to nogulsnēšanās rezervuārā? Tas notiek, veidojoties brīvi peldošai stadijai, klaidonam. Skropstas ķermeņa aizmugurējā galā parādās skropstu vainags (99. att.). Tajā pašā laikā peristomālais disks ievelkas uz iekšu un ciliāts atdalās no kātiņa. Iegūtais klaidonis spēj peldēt vairākas stundas. Pēc tam notikumi tiek izspēlēti apgrieztā secībā: skropstas ar savu aizmugures galu piestiprinās pie substrāta, aug kātiņš, samazinās skropstu aizmugurējā vainags, priekšējā galā iztaisnojas peristomālais disks un sāk darboties adorālās membrānas. Klaidoņu veidošanās suvojkā bieži ir saistīta ar aseksuālās vairošanās procesu. Infuzorija uz kātiņa sadalās, un viens no meitas indivīdiem (un dažreiz arī abi) kļūst par tramīgu un aizpeld.
Daudzi suvoku veidi nelabvēlīgos apstākļos spēj izveidoties.
Starp sēdošajiem skropstiņiem, kas pieder apaļo skropstu grupai, tikai salīdzinoši nedaudzas sugas, piemēram, iepriekš aplūkotās suvoks, ir vientuļas dzīvas formas. Lielākā daļa šeit iekļauto sugu ir koloniāli organismi.
Parasti kolonialitāte rodas nepilnīgi pabeigtas aseksuālās vai veģetatīvās reprodukcijas rezultātā. Reprodukcijas rezultātā izveidotie indivīdi lielākā vai mazākā mērā saglabā saikni savā starpā un kopā veido augstākas kārtas organisku individualitāti, kas vieno lielos daudzumos atsevišķi indivīdi, kas iegūst kolonijas nosaukumu (mēs jau esam tikušies ar koloniālo organismu piemēriem flagellātu klase.

Apaļciliātu kolonijas veidojas tā rezultātā, ka atdalītie īpatņi nepārvēršas par klaidoņiem, bet ar kātu palīdzību uztur kontaktus savā starpā (100. att.). Šajā gadījumā kolonijas galveno stublāju, kā arī tās pirmos zarus nevar attiecināt uz kādu no indivīdiem, bet gan pieder visai kolonijai kopumā. Dažkārt kolonija sastāv tikai no neliela skaita īpatņu, savukārt citās ciliātu sugās atsevišķu kolonijas īpatņu skaits var sasniegt vairākus simtus. Tomēr jebkuras kolonijas pieaugums nav neierobežots. Sasniedzot šai sugai raksturīgos izmērus, kolonija pārstāj palielināties un dalīšanās rezultātā izveidotajiem īpatņiem veidojas skropstu vainags, tie kļūst par klaidoņiem un aizpeld, veidojot jaunas kolonijas.
Apaļveida skropstu kolonijas ir divu veidu. Dažās vietās kolonijas kāts ir nesamazināms: kairinājuma gadījumā saraujas tikai atsevišķi kolonijas īpatņi, kas ievelk pinnātu, bet visa kolonija kopumā nemainās (šajā kolonijā ietilpst, piemēram, ģintis Epistylis, Opercularia). Citos (piemēram, Carchesium ģintī) visas kolonijas kāts spēj sarauties, jo citoplazma iet cauri visiem zariem un tādējādi savieno visus kolonijas indivīdus savā starpā. Kad šādas kolonijas tiek kairinātas, tās pilnībā saraujas. Visa kolonija šajā gadījumā reaģē kā veselums, kā organiska individualitāte.
Starp visiem koloniālajiem apaļiem ciliātiem īpaši interesē, iespējams, zootamnic(Zoothamnium arbuscula). Šī ciliāta kolonijas izceļas ar īpašu struktūras regularitāti. Turklāt šeit kolonijā ir iezīmēta interesanta polimorfisma bioloģiska parādība.

Zootamnijas kolonija izskatās kā lietussargs. Uz viena, galvenā, kolonijas kātiņa ir sekundārie zari (101. att.). Pieaugušo koloniju izmērs ir 2-3 mm, tāpēc tie ir skaidri redzami ar neapbruņotu aci. Zootamnii dzīvo mazie dīķi no tīrs ūdens. Viņu kolonijas parasti atrodas uz zemūdens augiem, visbiežāk uz elodejas (ūdens mēris).
Zootamnijas kolonijas kāti ir kontraktili, jo kontraktilā citoplazma iet cauri visiem kolonijas zariem, izņemot galvenā kātiņa bazālo daļu. Ar samazinājumu, kas notiek ļoti ātri un pēkšņi, visa kolonija savāc vienreizēju.

Zootamnijai raksturīgs stingri regulārs zaru izvietojums. Viens galvenais kāts ir piestiprināts pie pamatnes. Deviņi galvenie kolonijas zari atkāpjas no tās augšdaļas plaknē, kas ir perpendikulāra kātiņam, stingri regulāri izvietoti viens pret otru (102., 6. att.). No šiem zariem stiepjas sekundārie zari, uz kuriem sēž atsevišķi kolonijas indivīdi. Katrā sekundārajā zarā var būt līdz 50 ciliātiem. Kopējais īpatņu skaits kolonijā sasniedz 2-3 tūkstošus īpatņu.
Lielākā daļa kolonijas īpatņu pēc savas struktūras atgādina mazus atsevišķus suvoikus, kuru izmērs ir 40-60 mikroni. Bet bez maziem īpatņiem, kurus sauc par mikrozoīdiem, pieaugušajās kolonijās, aptuveni galveno zaru vidū, attīstās pavisam cita veida un izmēra īpatņi (102., 5. att.). Tie ir lieli sfēriski indivīdi, kuru diametrs ir 200–250 mikroni, kas simts vai vairāk reižu pārsniedz mikrozoīda tilpumu. Lielus indivīdus sauc par makrozoīdiem.
Savā struktūrā tie būtiski atšķiras no mazajiem kolonijas indivīdiem. Viņu peristoma nav izteikta: tā ir ievilkta uz iekšu un nefunkcionē. No paša attīstības sākuma no mikrozoīda makrozoīds pārstāj uzņemt barību pats par sevi. Tam trūkst gremošanas vakuolu. Makrozoīda augšana acīmredzot notiek uz vielu rēķina, kas nonāk caur citoplazmas tiltiem, kas savieno visus kolonijas indivīdus savā starpā. Makrozoja ķermeņa daļā, ar kuru tas ir piestiprināts pie kātiņa, ir uzkrājušies īpaši graudi (granulas), kuriem, kā mēs redzēsim, ir liela nozīme tā tālākajā liktenī. Kas ir šie lielie sfēriskie makrozoīdi, kāda ir to bioloģiskā loma zootamnijas kolonijas dzīvē? Novērojumi liecina, ka makrozoīdi ir nākotnes klaidoņi, no kuriem veidojas jaunas kolonijas. Sasniedzot izmēra ierobežojums, makrozoīdā veidojas skropstu vainags, tas atdalās no kolonijas un aizpeld. Tajā pašā laikā tā forma nedaudz mainās, no sfēriskas tā kļūst koniska. Pēc kāda laika tramps vienmēr tiek piestiprināts pie pamatnes ar to pusi, kurā atrodas granularitāte. Nekavējoties sākas kātiņa veidošanās un augšana, un kātiņa konstrukcijai tiek iztērētas granulas, kas lokalizējas klaidoņa aizmugurējā galā. Kātam augot, graudainība pazūd. Pēc tam, kad kāts sasniedz zootamnijai raksturīgo galīgo garumu, sākas virkne strauji secīgu dalījumu, kas noved pie kolonijas veidošanās. Šie dalījumi tiek veikti stingri noteiktā secībā (102. att.).
Mēs nekavēsimies pie šī procesa detaļām. Pievērsīsim uzmanību tikai šādai interesantajai parādībai. Zootamnijas trampju pirmajās dalīšanās laikā, veidojoties kolonijai veidojošajos īpatņos, nefunkcionē spārns un mute. Barošana sākas vēlāk, kad jaunā kolonija jau sastāv no 12-16 īpatņiem. Tādējādi visi pirmie kolonijas attīstības posmi tiek veikti tikai uz to rezervju rēķina, kas veidojās makrozoīda ķermenī tā augšanas un attīstības laikā mātes kolonijā. Pastāv nenoliedzama līdzība starp zootamnia vagrant attīstību un olšūnas attīstību daudzšūnu dzīvniekiem. Šī līdzība izpaužas faktā, ka attīstība abos gadījumos tiek veikta uz iepriekš uzkrāto rezervju rēķina, neuztverot pārtiku no ārējās vides.
Pētot sēdošos apaļcilitārus, rodas jautājums: kā tiek veikta skropstiņiem raksturīgā dzimumprocesa forma - konjugācija? Izrādās, ka saistībā ar mazkustīgu dzīvesveidu tas piedzīvo dažas būtiskas izmaiņas. Līdz dzimumprocesa sākumam kolonijā veidojas īpaši, ļoti mazi klaidoņi. Aktīvi pārvietojoties ar skropstu vainagu, viņi kādu laiku rāpo pa koloniju un pēc tam nonāk konjugācijā ar lieliem normāliem kolonijas sēdošiem indivīdiem. Tādējādi šeit notiek konjugantu diferenciācija divās indivīdu grupās: maza, kustīga (mikrokonjuganti) un lielāka, nekustīga (makrokonjuganti). Šī konjugantu iedalīšana divās kategorijās, no kurām viena (mikrokonjuganti) ir mobila, bija nepieciešama pielāgošanās mazkustīgam dzīvesveidam. Bez tā acīmredzot nevarētu nodrošināt normālu dzimumprocesa (konjugācijas) norisi.
SŪKŠANAS CILITARIUS (SUCTORIA)
Ļoti savdabīga grupa ēšanas veida ziņā ir pārstāvēta nepieredzējis ciliāti(Suctoria). Šie organismi, tāpat kā suvoyka un citi skropstu ciliāti, ir sēdoši. Šai kārtai piederošo sugu skaitu mēra ar vairākiem desmitiem. Sūcošo skropstu ķermeņa forma ir ļoti dažāda. Dažas to raksturīgās sugas ir parādītas 103. attēlā. Dažas sēž uz substrāta uz vairāk vai mazāk gariem kātiem, citiem nav stublāju, dažiem ķermenis diezgan spēcīgi zarojas utt. raksturo šādas divas pazīmes:
1) pilnīga ciliārā aparāta neesamība (pieaugušā formā),
2) īpašu piedēkļu klātbūtne - taustekļi, kas kalpo medījuma izsūkšanai.

Dažāda veida sūcošajām skropstiņām taustekļu skaits nav vienāds. Bieži vien viņi tiek savākti grupās. Ar lielu mikroskopa palielinājumu var redzēt, ka tausteklis galā ir aprīkots ar nelielu nūjas formas sabiezējumu.
Kā darbojas taustekļi? Uz šo jautājumu nav grūti atbildēt, kādu laiku novērojot sūcošus skropstiņus. Ja kāds mazs vienšūnis (flagellate, infusoria) pieskaras suktorijas tausteklim, tad tas uzreiz pieķersies pie tā. Visi upura mēģinājumi atrauties parasti ir veltīgi. Turpinot vērot pie taustekļiem pielipušo laupījumu, var redzēt, ka tas pamazām sāk samazināties. Tās saturs caur taustekļiem tiek pakāpeniski “iesūknēts” sūcošā skropstu endoplazmā, līdz no upura paliek tikai viens pīlings, kas tiek izmests. Tādējādi sūcošo skropstu taustekļi ir pilnīgi unikāli, nekur citur dzīvnieku pasaulē nav orgānu slazdīšanai un vienlaikus barības sūkšanai (103. att.).
Piesūcošas skropstas ir nekustīgi plēsēji, kas medījumu nevis dzenā, bet uzreiz noķer, ja vien neuzmanīgs medījums tiem pieskaras pats.

Kāpēc mēs šos savdabīgos organismus attiecinām uz ciliātu klasi? No pirmā acu uzmetiena viņiem nav nekāda sakara ar tiem. Sekojošie fakti runā par suktoriju piederību ciliātiem. Pirmkārt, tiem ir ciliātiem raksturīgs kodolaparāts, kas sastāv no makrokodola un mikrokodola. Otrkārt, vairošanās laikā viņiem attīstās skropstas, kuru nav "pieaugušiem" indivīdiem. Bezdzimuma pavairošana un tajā pašā laikā zīdošo skropstu pārvietošana tiek veikta, veidojot klaidoņus, kas aprīkoti ar vairākām gredzenveida skropstu vainagiem. Klaidoņu veidošanās suktorijās var notikt dažādos veidos. Dažkārt tie veidojas ne visai vienveidīgas dalīšanās (budding) rezultātā, kurā katra niere, kas atdalās uz āru, saņem makrokodolu segmentu un vienu mikrokodolu (104. att., L). Vienam mātes indivīdam vienlaikus var veidoties vairāki meitas pumpuri (104., 5. att.). Citām sugām (104. att., D, E) novērojama ļoti savdabīga "iekšējās pumpurēšanās" metode. Tajā pašā laikā mātes zīdīšanas ķermeņa iekšpusē veidojas dobums, kurā veidojas trampā nieres. Tas iznāk caur īpašiem caurumiem, caur kuriem tas “izspiežas” ar zināmām grūtībām.
Šī embrija attīstība mātes ķermenī un pēc tam bērna piedzimšana ir interesanta visvienkāršākā līdzība ar to, kas notiek augstākos daudzšūnu organismos.
Iepriekšējās lappusēs tika aplūkoti vairāki tipiski brīvi dzīvojoši ciliātu klases pārstāvji, kas atšķirīgi pielāgoti dažādiem vides apstākļiem. Interesanti ir pieiet jautājumam par ciliātu pielāgošanu dzīves apstākļiem un, no otras puses, redzēt, kas ir raksturīgas kopīgas iezīmes ciliāti, kas dzīvo noteiktos, stingri noteiktos vides apstākļos.
Kā piemēru ņemsim divus ļoti krasi atšķirīgus biotopus: dzīvību planktona sastāvā un dzīvību dibenā smilšu biezumā.
PLANKTONISKĀ INFUSORIJA
Diezgan liels skaits skropstu sugu sastopamas gan jūras, gan saldūdens planktonā.
Pielāgošanās dzīvei ūdens kolonnā iezīmes ir īpaši izteiktas radiolāros. Galvenā pielāgošanās līnija planktoniskajam dzīvesveidam ir samazināta līdz tādu struktūras iezīmju attīstībai, kas veicina organisma planēšanu ūdens stabā.

Tipiska planktoniska, turklāt gandrīz tikai jūras skropstu dzimta tintinīni(Tintinnidae, 105., 5. att.). Kopējais līdz šim zināmo tintinīdu sugu skaits ir aptuveni 300. Tās ir nelielas formas, kurām raksturīgs tas, ka ciliāta protoplazmiskais ķermenis atrodas caurspīdīgā, vieglā un vienlaikus spēcīgā mājā, kas sastāv no organiskām vielām. No mājas izvirzās disks, kurā ir skropstu vainags, kas atrodas pastāvīgā mirgojošā kustībā. Ūdens kolonnā peldošās infuzorijas stāvoklī to galvenokārt atbalsta pastāvīgs aktīvs ciliārā aparāta darbs. Māja, acīmredzot, veic ciliāta ķermeņa apakšdaļas aizsardzības funkciju. Saldūdenī dzīvo tikai 2 tintinīdu sugas (neskaitot 7 sugas, kas raksturīgas tikai Baikāla ezeram).
Saldūdens ciliātiem ir daži citi pielāgojumi dzīvei planktonā. Daudzās no tām citoplazma ir ļoti stipri vakuolēta (Loxodes, Condylostoma, Trachelius), tāpēc tā atgādina putas. Tas noved pie ievērojama īpatnējā smaguma samazināšanās. Visiem uzskaitītajiem ciliātiem turklāt ir ciliāra apvalks, pateicoties kuram skropstu ķermenis īpatnējā svara ziņā ir tikai nedaudz lielāks par īpaša gravitāteūdens, viegli uzturēts stāvoklī "planējošs". Dažām sugām ķermeņa forma veicina īpatnējās virsmas palielināšanos un atvieglo planēšanu ūdenī. Piemēram, daži Baikāla ezera planktona ciliāti pēc formas atgādina lietussargu vai izpletni (Liliomorpha, 105. att., 2). Baikāla ezerā sastopams viens planktona sūcējs ciliāts (Mucophrya pelagica, 105., 4. att.), kas krasi atšķiras no saviem sēdošajiem radiniekiem. Šai sugai nav stumbra. Tās protoplazmas ķermeni ieskauj plašs gļotains apvalks, kas ir pielāgojums, kas noved pie svara samazināšanas. Izceļas gari plāni taustekļi, kas līdz ar savu tiešo funkciju, iespējams, veic arī citu - īpatnējās virsmas palielināšanos, kas veicina planēšanu ūdenī.
Visbeidzot, jāmin vēl viena, tā teikt, netieša skropstu pielāgošanās formai dzīvei planktonā. Tā ir mazu skropstu pieķeršanās citiem organismiem, kas vada planktonisku dzīvesveidu. Jā, starp apgraizīti ciliāti(Peritricha) ir diezgan daudz sugu, kas pieķeras planktona copepodiem. Tas ir normāls un normāls šāda veida ciliātu dzīvesveids.
Kopā ar circumciliary ciliates un starp nepieredzējis(Suctoria) ir sugas, kas apmetas uz planktona organismiem.
MILTĪS DZĪVOTĀJI
Smilšainas pludmales un sēkļi ir ārkārtīgi savdabīgs biotops. Gar jūru piekrasti tie aizņem plašas telpas, un tos raksturo savdabīga fauna.
Veikts par pēdējie gadi Neskaitāmi pētījumi dažādās valstīs ir parādījuši, ka daudzu jūras smilšu slānis ir ļoti bagāts ar dažādu mikroskopisku vai mikroskopisku faunu. Starp smilšu daļiņām ir daudz mazu un niecīgu vietu, kas piepildītas ar ūdeni. Izrādās, ka šīs telpas ir bagātīgi apdzīvotas ar organismiem, kas pieder pie visdažādākajām dzīvnieku pasaules grupām. Šeit dzīvo desmitiem vēžveidīgo sugu, annelīdi, apaļtārpi, īpaši daudz plakano tārpu, daži gliemji, koelenterāti. Lielā skaitā sastopami arī vienšūņi, galvenokārt skropstas. Saskaņā ar mūsdienu datiem jūras smilšu biezumā mītošo skropstu faunā ir aptuveni 250-300 sugu. Ja paturam prātā ne tikai skropstiņus, bet arī citas organismu grupas, kas apdzīvo smilšu biezumu, tad kopējais to sugu skaits būs ļoti liels. Visu smilšu biezumu apdzīvojošo dzīvnieku kopumu, kas mīt mazākajās spraugās starp smilšu graudiem, sauc par psammofilo faunu.
Psammofilās faunas bagātību un sugu sastāvu nosaka daudzi faktori. Starp tiem, it īpaši nozīmi ir smilšu daļiņu izmērs. Rupjgraudainajām smiltīm ir slikta fauna. Nabadzīga ir arī ļoti smalkgraudainu dūņu smilšu (ar daļiņu diametru mazāku par 0,1 mm) fauna, kur acīmredzot atstarpes starp daļiņām ir pārāk mazas, lai tajās varētu dzīvot dzīvnieki. Visbagātākās smiltis ir vidēji un smalki graudainas.
Otrs faktors, kam ir svarīga loma psammofilās faunas attīstībā, ir smilšu bagātība organiskajās atliekās un sadalošās organiskās vielas (tā sauktā saprobitātes pakāpe). Smiltis, kurās nav organisko vielu, ir sliktas. No otras puses, smiltis ir arī gandrīz nedzīvas un ir ļoti bagātas ar organiskajām vielām, jo organisko vielu sabrukšana izraisa skābekļa samazināšanos. Bieži vien tam pievieno anaerobo sērūdeņraža fermentāciju.
Brīvā sērūdeņraža klātbūtne ir ārkārtīgi negatīvs faktors, kas ietekmē faunas attīstību.
Smilšu virsmas slāņos dažkārt veidojas diezgan bagāta vienšūnu aļģu flora (kramaļģes, peridīnijas). Tas ir faktors, kas veicina psammofīlās faunas attīstību, jo daudzi mazi dzīvnieki (tostarp skropstu dzimtas dzīvnieki) barojas ar aļģēm.
Visbeidzot, faktors, kas ļoti negatīvi ietekmē psammofilo faunu, ir sērfošana. Tas ir diezgan saprotams, jo sērfošana, mazgājot augšējos smilšu slāņus, nogalina visu dzīvo šeit. Psammofīlā fauna ir visbagātākā ar aizsargātiem, labi apsildāmiem līčiem. Ebb un plūsma nekavē psammofilās faunas attīstību. Kad bēguma laikā ūdens īslaicīgi aiziet, atsedzot smiltis, tad smilšu biezumā, intervālos starp smilšu graudiņiem, tas paliek, un tas neliedz dzīvniekiem eksistēt.
Ciliātos, kas ir daļa no psammofilās faunas un pieder pie dažādām sistemātiskām grupām (kārtām, ģimenēm), evolūcijas procesā veidojas daudzas kopīgas pazīmes, kas ir pielāgošanās savdabīgajiem eksistences apstākļiem starp smilšu daļiņām.

106. attēlā parādītas dažas psammofilās faunas sugas, kas pieder pie dažādām kārtām un ģimenēm. Starp tiem ir daudz līdzību. Lielākajai daļai no tiem ķermenis ir vairāk vai mazāk stipri izstiepts garumā, tārps. Tas ļauj viegli "saspiesties" mazākajos caurumos starp smilšu graudiņiem. Ļoti daudzām sugām (106. att.) ķermeņa pagarinājums ir apvienots ar tā saplacināšanu. Ciliārais aparāts vienmēr ir labi attīstīts, kas ļauj aktīviem ar noteiktu spēku pārvietoties šaurās spraugās. Diezgan bieži skropstas attīstās vienā tārpveidīgā saplacinātā ķermeņa pusē, pretējā puse ir kaila. Šī īpašība, iespējams, ir saistīta ar spēju, kas ir izteikta lielākajai daļai psammofīlo sugu, ļoti cieši un ļoti stingri pielipt (pieķerties) substrātam caur ciliāro aparātu (parādība, ko sauc par thigmotaksi). Šis īpašums ļauj dzīvniekiem palikt uz vietas gadījumos, kad šaurajās spraugās, kur tie dzīvo, rodas ūdens straumes. Šajā gadījumā, iespējams, ir izdevīgāk, ja tā puse, kas ir pretēja tai pusei, uz kuras dzīvnieks piestiprinājās pie substrāta, ir gluda.
Ko ēd psammofīlie ciliāti? Ievērojama daudzu sugu "uztura" daļa ir aļģes, īpaši kramaļģes. Baktērijas mazākā mērā tām kalpo kā barība. Tas lielā mērā ir atkarīgs arī no tā, ka smiltīs, kas nav īpaši piesārņotas, ir maz baktēriju. Visbeidzot, it īpaši starp lielākajiem psammofilajiem skropstiņiem, ir ievērojams skaits plēsīgo formu, kas ēd citus mazākām sugām piederīgus skropstiņus. Psammophilic ciliāti ir izplatīti, acīmredzot, visur.
INFUSORIJAS APOSTOMĀTI

ciliāti spirofrija(Spirophrya subparasitica) ensistētā stāvoklī bieži var atrast sēžam uz neliela kātiņa uz maziem planktona jūras vēžveidīgajiem (īpaši bieži uz Idia ģints vēžveidīgajiem). Kamēr vēžveidīgais aktīvi peld jūras ūdenī, uz tā sēdošā spirofrija nemainās. Ciliātu tālākai attīstībai nepieciešams, lai vēžveidīgo apēstu jūras hidroīda polips, kas notiek diezgan bieži (107. att.). Tiklīdz spirofrijas cistas kopā ar vēžveidīgo nonāk gremošanas dobumā, no tām uzreiz iznāk mazi skropstiņi, kas sāk enerģiski baroties ar norītā vēžveidīgo sagremošanas rezultātā radušos barības vircu. Stundas laikā infuzorijas izmērs palielinās 3-4 reizes. Tomēr reprodukcija šajā posmā nenotiek. Pirms mums ir tipisks ciliātu augšanas posms, ko sauc par trofontu. Pēc kāda laika kopā ar nesagremotajām pārtikas atliekām trofontu polips izmet jūras ūdenī. Šeit, aktīvi peldot, tas nolaižas gar polipa ķermeni līdz zolei, kur tas piestiprina sevi, būdams cistas ieskauts. Šo encistētu, lielu ciliātu, kas sēž uz polipa, sauc par tomontu. Šis ir audzēšanas posms. Tomonts nebarojas, bet ātri sadalās vairākas reizes pēc kārtas (107., 7. att.). Rezultāts ir vesela grupa ļoti mazu ciliātu. To skaits ir atkarīgs no tomonta lieluma, ko savukārt nosaka trofonta lielums, kas tam piešķīris izcelsmi. Tomonta dalīšanās rezultātā izveidojušies mazie ciliāti (tos sauc par tomītiem vai klaidoņiem) pārstāv apmetnes stadiju.
Viņi atstāj cistu, ātri peld (neēdot vienlaikus, bet izmantojot rezerves, kas viņiem ir citoplazmā). Ja viņiem ir “paveicies” sastapt spārnu kāju, viņi nekavējoties pieķeras tai un pieķeras. Šis ir posms, no kura mēs sākām cikla apsvēršanu.
Apskatītajā dzīves cikls spirophria vērš uzmanību uz to posmu krasi norobežošanu, kuriem ir atšķirīga bioloģiskā nozīme. Trofons ir augšanas stadija. Tas tikai aug, uzkrāj lielu daudzumu citoplazmas un visa veida rezerves vielas, pateicoties spēcīgai un ātrai barošanai. Trofonts nav spējīgs vairoties. Tomontā tiek novērota pretēja parādība - nespēja baroties un enerģiska ātra vairošanās. Pēc katras dalīšanas augšana nenotiek, un tāpēc tomonta vairošanās tiek samazināta līdz straujai sabrukšanai daudzos klaidoņos. Visbeidzot, klaidoņi pilda savu īpašo un vienīgo raksturīgo funkciju: tie ir indivīdi – sugas iemītnieki un izplatītāji. Viņi nespēj ēst vai vairoties.
Ihtioftirija DZĪVES CIKLS

Līdz augšanas perioda beigām ichthyophthirius sasniedz ļoti lielu izmēru, salīdzinot ar klaidoņiem: 0,5-1 mm diametrā. Sasniedzot robežvērtību, skropstas aktīvi virzās ārā no zivju audiem ūdenī un kādu laiku lēnām peld ar ciliārā aparāta palīdzību, kas aptver visu ķermeni. Drīz vien lielie ihtioftiri apmetas uz kāda zemūdens objekta un izdala cistu. Tūlīt pēc enstēšanas sākas ciliātu secīga dalīšanās: vispirms uz pusēm, tad katrs meitas indivīds atkal tiek sadalīts divās un tā līdz 10-11 reizēm. Rezultātā cistas iekšpusē veidojas līdz 2000 mazu, gandrīz noapaļotu īpatņu, kas pārklāti ar skropstiņām. Cistas iekšpusē klaidoņi aktīvi pārvietojas. Viņi caurdur čaulu un iznāk. Aktīvi peldošie klaidoņi inficē jaunas zivis.
Ihtioftirija dalīšanās ātrums cistās, kā arī augšanas ātrums zivju audos lielā mērā ir atkarīgs no temperatūras. Pēc dažādu autoru pētījumiem ir doti šādi skaitļi: 26-27°C klaidoņu attīstības process cistā ilgst 10-12 stundas, 15-16°C temperatūrā 28-30 stundas, 4. -5°C tas ilgst 6 -7 dienas.
Cīņa pret ihtioftiriju rada ievērojamas grūtības. Šeit primāri svarīgi ir preventīvie pasākumi, kuru mērķis ir novērst brīvi peldošu klaidoņu iekļūšanu zivju audos. Lai to izdarītu, ir lietderīgi veikt biežu slimu zivju transplantāciju jaunos rezervuāros vai akvārijos, lai radītu plūsmas apstākļus, kas ir īpaši efektīvi cīņā pret ihtioftiriju.
ciliates trichodynes

Visa trihodīnu adaptācijas sistēma dzīvībai uz saimnieka virsmas ir vērsta uz to, lai neatrautos no saimnieka ķermeņa (kas gandrīz vienmēr ir līdzvērtīgs nāvei), vienlaikus saglabājot mobilitāti. Šīs ierīces ir ļoti ideālas. Lielākajai daļai trihodīnu ķermenis ir diezgan plakana diska formā, dažreiz vāciņš. Puse, kas vērsta pret saimnieka ķermeni, ir nedaudz ieliekta, veido pieķeršanās piesūcekni. Gar piesūcekņa ārējo malu atrodas labi attīstītu skropstu vainags, ar kura palīdzību skropstu kustība (rāpošana) galvenokārt notiek pa zivs ķermeņa virsmu. Šis vainags atbilst iepriekš aplūkotajiem sēdošajiem apaļcilitainajiem klaidoņiem sastopamajam vainagu. Tādējādi trihodīnu var salīdzināt ar trampu. Uz vēdera virsmas (uz piesūcekņa) trihodīniem ir ļoti sarežģīts atbalsta un piestiprināšanas aparāts, kas palīdz noturēt skropstiņus uz saimnieka. Neiedziļinoties tās struktūras detaļās, mēs atzīmējam, ka tā pamatā ir sarežģītas konfigurācijas gredzens, kas sastāv no atsevišķiem segmentiem ar ārējiem un iekšējiem zobiem (109. att., B). Šis gredzens veido elastīgu un vienlaikus spēcīgu vēdera virsmas pamatu, kas darbojas kā piesūceknis. Dažādi trihodīna veidi atšķiras viens no otra ar gredzenu veidojošo segmentu skaitu, kā arī ārējo un iekšējo āķu konfigurāciju.
Trichodina korpusa pusē, kas atrodas pretī diskam, ir pinnāts un mutes aparāts. Tās struktūra ir vairāk vai mazāk raksturīga apgraizīti ciliāti. Pulksteņa kustības virzienā savītas adorālās membrānas ved uz padziļinājumu, kura apakšā ir mute. Trihodīna kodolaparāts parasti ir sakārtots ciliātiem: viens lentei līdzīgs makrokodols un viens mikrokodols, kas atrodas blakus. Ir viena saraušanās vakuole.
Trihodīni ir plaši izplatīti visu veidu rezervuāros. Īpaši bieži tie sastopami dažādu zivju sugu mazuļiem. Ar masveida pavairošanu tiek pielietoti trihodīni liels kaitējums zivis, it īpaši, ja masas pārklāj žaunas. Tas traucē normālu zivju elpošanu.
Lai zivis attīrītu no trihodīna, ārstnieciskās vannas ieteicams pagatavot no 2% nātrija hlorīda šķīduma vai 0,01% kālija permanganāta šķīduma (cepumiem - 10-20 minūtes).
UNGATU ZARNU TRAKTA INFŪZIJAS
No rētas caur sietu ēdiens tiek atraugas mutes dobumā, kur tas tiek papildus sakošļāts (košļājamā gumija). Atkal pa speciālu barības vada krokām veidotu caurulīti norīta sakošļātā pārtikas masa vairs nenonāk rētā, bet gan grāmatā un no turienes uz vēdera dobumu, kur tā tiek pakļauta atgremotāju gremošanas sulām. Siera fermentā skābes reakcijas apstākļos un gremošanas enzīmu klātbūtnes apstākļos skropstiņi mirst. Nokļūstot ar košļājamo gumiju, tie tiek sagremoti.
Vienšūņu skaits spureklī (kā arī tīklā) var sasniegt kolosālas vērtības. Ja paņem pilienu rētas satura un apskati to mikroskopā (karsējot, jo istabas temperatūrā skropstas apstājas), tad skropstas burtiski spieto redzes laukā. Pat kultūrā ir grūti iegūt šādu ciliātu masu. Ciliātu skaits 1 cm3 rētas satura sasniedz miljonu, un bieži vien vairāk. Runājot par visu rētas apjomu, tas sniedz patiesi astronomiskus skaitļus! Spurekļa satura bagātība ar ciliātiem lielā mērā ir atkarīga no atgremotāju barības veida. Ja barība ir bagāta ar šķiedrvielām un nabadzīga ar ogļhidrātiem un olbaltumvielām (zāle, salmi), tad spureklī ir salīdzinoši maz skropstu. Ja diētai pievieno ogļhidrātus un olbaltumvielas (klijas), skropstu skaits krasi palielinās un sasniedz milzīgu skaitu. Jāpatur prātā, ka pastāv pastāvīga ciliātu aizplūšana. Nokļūstot kopā ar košļājamo gumiju vēdera dobumā, viņi mirst. Augsts līmenis ciliātu skaitu uztur to enerģiska vairošanās.
Arī pārnadžu (zirgs, ēzelis, zebra) gremošanas traktā ir liels skaits skropstu, taču to lokalizācija saimniekorganismā ir atšķirīga. Pāra nagaiņiem nav sarežģīta vēdera, kā dēļ nav iespējams attīstīt vienšūņus gremošanas trakta priekšējās daļās. Bet zirgu dzimtas dzīvniekiem ļoti labi attīstīti ir lielie un aklās zarnas, kuras parasti ir aizsērējušas ar barības masām un kurām ir būtiska loma gremošanā. Šajā zarnu daļā, tāpat kā atgremotāju spureklī un sietā, veidojas ļoti bagāta vienšūņu fauna, galvenokārt skropstas, no kurām lielākā daļa arī pieder endodiniomorfo kārtai. Taču sugu sastāva ziņā atgremotāju spurekļa fauna un zirgu dzimtas dzīvnieku resnās zarnas fauna nesakrīt.
ATGRĒJOŠU ZARNU TRAKTA CILĪTI
Vislielāko interesi rada ciliāti ofrioskolecīdu ģimene(Ophryoscolecidae), kas saistīti ar kārtas endodiniomorfs. Šīs atdalīšanas raksturīga iezīme ir nepārtraukta ciliāra apvalka trūkums. Sarežģīti ciliāru veidojumi - cirres - atrodas ciliātu ķermeņa priekšējā galā mutes atveres rajonā. Šiem ciliārā aparāta pamatelementiem var pievienot papildu cireju grupas, kas atrodas ķermeņa priekšējā vai aizmugurējā galā. Kopējais ofrioskolecīdu dzimtas ciliātu sugu skaits ir aptuveni 120.

110. attēlā parādīti daži no raksturīgākajiem ofrioskolecīdiem no atgremotāju spurekļa. Entodinium ģints ciliāti (Entodinium, 110. att., L) ir visvienkāršāk sakārtoti. Viņu ķermeņa priekšējā galā ir viena periorāla cirozes zona. Korpusa priekšējo galu, uz kura atrodas mutes atvere, var ievilkt uz iekšu. Ektoplazma un endoplazma ir krasi norobežotas. Anālā caurule ir skaidri redzama aizmugurējā galā, kas kalpo, lai noņemtu nesagremotas pārtikas atliekas. Nedaudz sarežģītāka struktūra anoplodīnija(Anoplodīnijs, 110. att., B). Viņiem ir divas ciliārā aparāta zonas - periorālā cirra un muguras cirra. Abi atrodas priekšpusē. Attēlā redzamās sugas ķermeņa aizmugurē ir gari asi izaugumi - tas ir diezgan raksturīgi daudzām ofrioskolecīdu sugām. Ir ierosināts, ka šie izaugumi veicina ciliātu "stumšanu" starp augu daļiņām, kas aizpilda rētu.
Veidi Eudiplodynia ģints(Eudiplodinium, 110. att., B) ir līdzīgi anoplodīnija, bet, atšķirībā no tiem, ir skeleta pamatplāksne, kas atrodas labajā malā gar rīkli. Šī skeleta plāksne sastāv no vielas, kas ir līdzīga ķīmiskā daba uz šķiedrvielām, tas ir, uz vielu, kas veido augu šūnu čaulas.
Plkst poliplastronu ģints(Polyplastron, 110. att., D, E) ir vēl viena skeleta komplikācija. Šo ciliātu struktūra ir tuva eidiplodinijai. Atšķirības galvenokārt ir saistītas ar faktu, ka vienas skeleta plāksnes vietā šiem ciliātiem ir pieci. Divas no tām, lielākās, atrodas ciliāta labajā pusē, un trīs, mazākas, atrodas ciliāta kreisajā pusē. Otrā poliplastrona iezīme ir kontraktilo vakuolu skaita palielināšanās. Entodinijai ir viena saraušanās vakuole, anoplodinijai un eudiplodinijai ir divas kontrakcijas vakuolas, un poliplastronam ir apmēram ducis no tiem.
Plkst epidīnijs(Epidinium, 110. att.), kuriem ir labi attīstīts ogļhidrātu skelets, kas atrodas ķermeņa labajā pusē, cirras muguras zona pāriet no priekšējā gala uz muguras pusi. Muguriņi bieži attīstās šīs ģints ciliātu aizmugurējā galā.
Sarežģītākā struktūra atklāj Ofrioscolex ģints(Ophryoscolex), pēc kura nosaukta visa ciliātu dzimta (110. att., E). Viņiem ir labi attīstīta cirras muguras zona, kas aptver apmēram 2/3 no ķermeņa un skeleta plākšņu apkārtmēra. Aizmugurējā galā ir izveidoti daudzi muguriņas, no kuriem viens parasti ir īpaši garš.
Iepazīšanās ar dažiem tipiskiem pārstāvjiem ofrioskolecīds parāda, ka šīs ģimenes ietvaros ir bijis ievērojams organizācijas sarežģījums (no entodīnijas līdz ofrioskoleksam).
Papildus ciliātiem ofrioskolecīdu ģimene, atgremotāju spureklī nelielos daudzumos sastopami mums jau zināmo pārstāvji. izocilāru ciliātu atdalīšanās. Tos pārstāv neliels skaits sugu. Viņu ķermenis vienmērīgi klāts ar gareniskām skropstu rindām, skeleta elementu nav. Kopējā spurekļa skropstu populācijas masā tiem nav būtiskas nozīmes, un tāpēc mēs šeit par tiem nekavēsimies.
Ko un kā ēd rioskolecīdu ciliāti? Šo jautājumu ir sīki pētījuši daudzi zinātnieki, īpaši detalizēti profesors V. A. Dogels.

Ofrioskolecīdu barība ir diezgan daudzveidīga, un dažādās sugās tiek novērota noteikta specializācija. Mazākās Entodynia ģints sugas barojas ar baktērijām, cietes graudiem, sēnēm un citām mazām daļiņām. Ļoti daudzi vidēji un lieli ofrioskolecīdi absorbē augu audu daļiņas, kas veido lielāko daļu spurekļa satura. Dažu sugu endoplazma ir burtiski aizsērējusi ar augu daļiņām. Var redzēt, kā skropstas uzbrūk augu audu lūžņos, burtiski saplēš tos gabalos un pēc tam norij, bieži savā ķermenī savērpjot spirālē (111., 4. att.). Dažkārt ir jāvēro tādi attēli (111., 2. att.), kad paša ciliāta ķermenis ir deformēts norīto lielo daļiņu dēļ.
Ofrioskolecīda gadījumā dažreiz tiek novērota plēsonība. Lielākas sugas aprij mazākās. Plēsonība (112. att.) ir apvienota ar vienas sugas spēju baroties ar augu daļiņām.

Kā ciliāti iekļūst atgremotāja spureklī? Kādi ir inficēšanās ceļi ar ofrioskolecīdiem? Izrādās, ka jaundzimušajiem atgremotājiem spureklī vēl nav ciliātu. To arī nav, kamēr dzīvnieks barojas ar pienu. Bet, tiklīdz atgremotājs pāriet uz augu barību, rētā un sietā uzreiz parādās skropstiņi, kuru skaits strauji pieaug. No kurienes viņi nāk? Ilgu laiku tika pieņemts, ka spurekļa skropstas veido sava veida atpūtas stadijas (visticamāk, cistas), kas dabā ir plaši izplatītas un, norijot, izraisa aktīvas ciliātu stadijas. Turpmākie pētījumi ir parādījuši, ka atgremotāju ciliātiem nav atpūtas stadiju. Bija iespējams pierādīt, ka inficēšanās notiek ar aktīviem kustīgiem ciliātiem, kas, atraugas, iekļūst mutes dobumā. Ja jūs izmeklējat zem mikroskopa, kas ņemts no mutes dobums košļājamā gumija, bet tajā vienmēr ir liels skaits aktīvi peldošu ciliātu. Šīs aktīvās formas var viegli iekļūt mutē un tālāk citu atgremotāju spureklī no kopīga dzeršanas trauka kopā ar zāli, sienu (kurā var būt siekalas ar ciliātiem) utt. Šis infekcijas ceļš ir pierādīts eksperimentāli.
Ja ofrioskolecīdos nav atpūtas stadiju, tad acīmredzami ir viegli iegūt "bez infūzijas" dzīvniekus, izolējot tos, kad tie vēl barojas ar pienu. Ja augošajiem mazuļiem un atgremotājiem nav pieļaujams tiešs kontakts ar skropstiņiem, tad jaunus dzīvniekus spureklī var atstāt bez skropstiņiem. Šādus eksperimentus veica vairāki zinātnieki dažādās valstīs. Rezultāts bija skaidrs. Ja jaundzivniekiem (barošanas laikā no mātes paņemts piens) un atgremotājiem nav kontakta ar skropstiņiem spureklī, dzīvnieki izaug sterili attiecībā pret skropstiņiem. Taču, lai sterilu dzīvnieku spureklī parādītos skropstiņi, pietiek ar pat īslaicīgu kontaktu ar dzīvniekiem ar ciliātiem (kopīga barotava, kopīgs spainis dzeršanai, kopīgas ganības).
Iepriekš bija eksperimentu rezultāti par atgremotāju saturu, kuriem spureklī un tīklā pilnībā nebija ciliātu. Tas tiek panākts, kā mēs redzējām, agrīni izolējot jauniešus. Eksperimenti tika veikti ar aitām un kazām.
Tādā veidā bija iespējams veikt "bez infūzijas" dzīvnieku novērojumus ievērojamu laika periodu (vairāk nekā gadu). Kā skropstu neesamība spureklī ietekmē saimnieka dzīvi? Vai skropstu neesamība ietekmē saimniekorganismu negatīvi vai pozitīvi? Lai atbildētu uz šo jautājumu, ar kazām tika veikti šādi eksperimenti. Dvīņu mazuļi (viena metiena un viena dzimuma) tika ņemti, lai iegūtu vairāk līdzīgu materiālu. Tad viens no šī pāra dvīņiem tika audzināts bez skropstiņiem spureklī (agrīna izolācija), bet otrs jau no paša sākuma, kad barojās ar augu pārtiku, bija bagātīgi inficēts ar daudzu veidu skropstiņiem. Abi saņēma tieši tādu pašu diētu un tika audzināti tādos pašos apstākļos. Vienīgā atšķirība starp tām bija ciliātu klātbūtne vai neesamība. Vairākiem kazlēnu pāriem, kas tika pētīti šādā veidā, netika konstatētas atšķirības katra pāra abu locekļu ("infuzors" un "neinfuzors") attīstības gaitā. Tādējādi var apgalvot, ka spureklī un tīklā mītošiem skropstiņiem nav nekādas krasas ietekmes uz saimniekdzīvnieka dzīvībai svarīgām funkcijām.
Iepriekš minētie eksperimentu rezultāti tomēr neļauj apgalvot, ka spurekļa ciliāti ir pilnīgi vienaldzīgi pret saimnieku. Šie eksperimenti tika veikti ar parastu saimnieka uzturu. Iespējams, citos apstākļos ar atšķirīgu uzturu (piemēram, ar nepietiekamu barošanu) izdosies atklāt spureklī mītošo vienšūņu faunas ietekmi uz saimnieku.
Literatūrā ir izteikti dažādi ieteikumi par spurekļa vienšūņu faunas iespējamo pozitīvo ietekmi uz saimnieka gremošanas procesiem. Tika norādīts, ka daudzi miljoni skropstu, aktīvi peldoties spureklī un drupinot augu audus, veicina gremošanas trakta priekšējās daļās esošo pārtikas masu fermentāciju un gremošanu. Ievērojams skaits ciliātu, kas kopā ar košļājamo gumiju nonāk abomasum, tiek sagremoti, un tiek absorbēts proteīns, kas veido ievērojamu skropstu ķermeņa daļu. Tāpēc infuzorijas var būt papildu olbaltumvielu avots saimniekam. Ir arī ierosināts, ka ciliāti veicina šķiedrvielu, kas veido lielāko daļu atgremotāju barības, sagremošanu un tās pārnešanu uz vieglāk sagremojamu stāvokli.
Visi šie pieņēmumi nav pierādīti, un pret dažiem no tiem ir izteikti iebildumi. Tika norādīts, piemēram, ka ciliāti veido sava ķermeņa protoplazmu no olbaltumvielām, kas kopā ar saimnieka barību nonāk spureklī. Absorbējot augu olbaltumvielas, tās pārvērš to ķermeņa dzīvnieku olbaltumvielās, kuras pēc tam tiek sagremotas abomasumā. Joprojām nav skaidrs, vai tas sniedz kādu labumu saimniekam. Visi šie jautājumi rada lielu praktisku interesi, jo mēs runājam par atgremotāju - galveno lopkopības objektu - gremošanu. Ir ļoti vēlami turpmāki pētījumi par spurekļa ciliātu lomu atgremotāju gremošanu.
Atgremotāju ofrioskolecīdiem, kā likums, ir plaša specifika. Sugu ziņā liellopu, aitu un kazu rētas un tīkla populācija ir ļoti tuvu viena otrai. Ja salīdzinām Āfrikas antilopu spurekļa sugu sastāvu ar liellopiem, tad arī šeit aptuveni 40% no kopējā sugu skaita būs izplatīti. Tomēr ir daudzas ofrioskolecīda sugas, kas sastopamas tikai antilopēs vai tikai briežos. Tādējādi uz ofrioskolecīdu vispārējās plašās specifikas fona var runāt par atsevišķiem, šaurāk specifiskiem to veidiem.
Zirgu dzimtas dzīvnieku zarnu infuzorijas
Tagad pievērsīsimies īsai iepazīšanai ar ciliātiem, kas apdzīvo zirgu dzimtas dzīvnieku lielos un aklās zarnas.
Arī šī fauna sugu ziņā, tāpat kā atgremotāju spurekļa fauna, ir ļoti daudzveidīga. Šobrīd ir aprakstītas aptuveni 100 ciliātu sugas, kas dzīvo zirgu dzimtas dzīvnieku resnajā zarnā. Šeit sastopamie skropstiņi pēc piederības dažādām sistemātiskām grupām ir daudzveidīgāki nekā atgremotāju spurekļa skropstas.

Zirgu zarnās ir diezgan daudz ciliātu sugu, kas pieder pie izocilāru kārtas, tas ir, ciliāru aparāts neveido membrānu vai cirozi netālu no mutes zonas (113. att., 1).
Kārtības entodīniomorfs(Entodiniomorpha) ir bagātīgi pārstāvēta arī zirga zarnās. Kamēr atgremotāju spureklī ir sastopama tikai viena endodiniomorfu dzimta (ofrioskolecīdu dzimta), zirga zarnās dzīvo trīs ģimeņu pārstāvji, taču pie kuru īpašībām šeit nepakavēsimies, aprobežojoties tikai ar dažiem tipiskas zirgu sugas (113. att.) .
Detalizēti A. Strelkova pētījumi parādīja, ka dažāda veida ciliāti nebūt nav vienmērīgi izkliedēti pa zirga resnajām zarnām. Ir divas dažādas sugu grupas, divas faunas, it kā. Viena no tām apdzīvo aklajā zarnā un resnās zarnas vēdera daļā (resnās zarnas sākotnējās sekcijās), bet otra apdzīvo lielās un mazās resnās zarnas muguras daļu. Šie divi sugu kompleksi ir diezgan krasi norobežoti. Šīm divām sekcijām kopīgu sugu ir maz — mazāk nekā duci.
, 
Interesanti atzīmēt, ka starp daudzajām skropstu sugām, kas apdzīvo zirgu dzimtas dzīvnieku resnās zarnas, ir vienas ģints pārstāvji, kas saistīti ar zīdītājiem. Kā mēs redzējām iepriekš, nepieredzējis ciliāti(Suctoria) ir tipiski brīvi dzīvojoši sēdoši organismi ar ļoti īpašu barošanās veidu ar taustekļu palīdzību (103. att.). Viens no dzemdību sūkšana pielāgota tādam šķietami neparastam biotopam kā zirga resnā zarna, piemēram, vairākas sugas alantoze(Alantosoma). Šiem ļoti savdabīgajiem dzīvniekiem (114. att.) nav kātiņa, nav skropstu, labi attīstīti galos sabiezināti nūjveida taustekļi.
Ar taustekļu palīdzību alantosomas pielīp pie dažāda veida skropstiņiem un izsūc tos. Bieži vien upuris ir daudzkārt lielāks par plēsēju.
Jautājums par attiecību raksturu starp zirgu dzimtas dzīvnieku resnās zarnas ciliātiem un to saimniekiem joprojām ir neskaidrs. Skropstu skaits var būt tikpat liels un dažreiz pat vairāk nekā atgremotāju spureklī. Ir dati, kas liecina, ka ciliātu daudzums zirga resnajās zarnās var sasniegt 3 miljonus 1 cm3. Dažu zinātnieku ieteiktā simbiotiskā nozīme ir vēl mazāk iespējama nekā spurekļa ciliātiem.
Visticamāk uzskata, ka tie rada zināmu kaitējumu saimniekam, uzņemot ievērojamu daudzumu pārtikas. Daļa ciliātu tiek izvadīta ar fekāliju masām, un tādējādi organiskās vielas (ieskaitot olbaltumvielas), kas veido to ķermeni, saimnieks paliek neizmantotas.
Jautājums par zirgu dzimtas dzīvnieku inficēšanās veidiem ar ciliātiem, kas apdzīvo resnajā zarnā, vēl nav atrisināts.

Balantidia uztver dažādas pārtikas daļiņas no resnās zarnas satura. Īpaši labprāt viņš barojas ar cietes graudiem. Ja balantidia dzīvo cilvēka resnās zarnas lūmenā, tad tā barojas ar zarnu saturu un tai nav kaitīgas ietekmes. Tas ir tipisks "kariete", kuru mēs satikām jau, domājot par dizentērijas amēbu. Tomēr balantidia ir mazāka iespēja nekā dizentērijas amēba palikt tik "nekaitīgs īrnieks".
Šobrīd speciālisti ir labi attīstījušies dažādas metodes, ļaujot kultivēt balantīdijas mākslīgā vidē – ārpus saimniekorganisma.

Kā redzams attēlā, troglodptella ir viens no kompleksajiem endodiniomorfiem. Viņai papildus periorālajai cirozes zonai (ķermeņa priekšējā galā) ir vēl trīs labi attīstītas cirozes zonas, kas gredzenveida pārklāj ciliāta ķermeni. Trogloditeliem ir labi attīstīts skeleta aparāts, kas sastāv no ogļhidrātiem, kas aptver gandrīz visu ķermeņa priekšējo galu. Šo savdabīgo ciliātu izmēri ir diezgan nozīmīgi. Garumā tie sasniedz 200-280 mikronus.
Bez mutes ciliāti ASTOMATS

Balstošie skeleta veidojumi veidojas galvenokārt ķermeņa priekšējā galā, kam ir jāpiedzīvo mehāniska slodze un jāpārvar šķēršļi, izspiežoties caur zarnu lūmenu starp pārtikas daļiņām. Sugas radiofrijas ģints(Radiophrya) priekšējā galā vienā ķermeņa pusē (ko nosacīti uzskata par vēdera pusi) ir ļoti spēcīgas elastīgas ribas (spiculas), kas atrodas ektoplazmas virsmas slānī (117. att., B, D, E) . Sugas Menilella ģints(Mesnilella) ir arī atbalsta stari (spiculas), kas lielāko daļu sava garuma atrodas citoplazmas dziļākajos slāņos (endoplazmā, 117. att., A). Līdzīgi izkārtoti atbalsta veidojumi veidojas arī dažu citu Astomat ģinšu sugās.
Dažiem ciliātiem astomat norit savdabīgā veidā. Lielākajai daļai ciliātu raksturīgā šķērseniskā dalījuma divos vietā daudziem astromātiem ir nevienmērīgs dalījums (budings). Tajā pašā laikā nieres, kas atdalās aizmugurējā galā, kādu laiku paliek saistītas ar mātes indivīdu (117. att., B). Rezultātā tiek iegūtas ķēdes, kas sastāv no priekšējiem lieliem un aizmugurējiem mazākiem indivīdiem (nierēm). Nākotnē nieres pakāpeniski atdalās no ķēdes un pāriet uz neatkarīgu eksistenci. Šī savdabīgā vairošanās forma ir plaši izplatīta, piemēram, mums jau zināmajā radiofrijā. Dažu astomu ķēdes, kas radušās pumpuru veidošanās rezultātā, ir līdzīgas izskats lenteņu ķēdes. Šeit mēs atkal sastopamies ar konverģences fenomenu.
Astomāta kodolaparātam ir ciliātiem raksturīga uzbūve: makrokodolu, visbiežāk lentveida formas (117. att.), un viens mikrokodolu. Kontrakcijas vakuoli parasti ir labi attīstīti. Lielākajai daļai sugu ir vairākas (dažreiz vairāk nekā desmit) kontrakcijas vakuolas, kas sakārtotas vienā gareniskā rindā.
Pētījums par Astomat sugu izplatību starp dažāda veida saimniekiem liecina, ka lielākā daļa Astomat sugu ir ierobežotas ar stingri noteiktām saimnieksugām. Lielākajai daļai astromātu ir raksturīga šaura specifika: par saimnieku tiem var kalpot tikai viena dzīvnieku suga.
Neraugoties uz lielo pētījumu skaitu, kas veltīts astomātu ciliātu izpētei, viens ļoti svarīgs to bioloģijas aspekts joprojām ir pilnīgi neskaidrs: kā notiek šo ciliātu pārnešana no viena saimnieka indivīda uz otru? Nekad nav bijis iespējams novērot cistu veidošanos šajos ciliātos.
Tāpēc tiek ierosināts, ka infekcija notiek aktīvi - mobilās stadijas.
JŪRAS EŽA ZARNU INFŪZIJAS
Jūras eži ir ļoti daudz mūsu ziemeļu (Barenca) un Tālo Austrumu jūras (Japānas jūrā, Kuriļu salu Klusā okeāna piekrastē) piekrastes zonā. Vairums jūras eži barojas ar augu barību, galvenokārt aļģēm, kuras tās nokasa no zemūdens priekšmetiem ar īpašiem asiem "zobiem", kas apņem mutes atveri. Šo zālēdāju ežu zarnās ir bagāta skropstu fauna. Nereti tās šeit attīstās masveidā, un jūras ežu zarnu saturs zem mikroskopa ir gandrīz tikpat "piesātināts" ar infuzorijām kā atgremotāju spurekļa saturs. Jāteic, ka līdzās dziļajām atšķirībām jūras ežu zarnu un atgremotāju spurekļa ciliātu dzīves apstākļos ir arī dažas līdzības. Tie slēpjas apstāklī, ka gan šur, gan tur ciliāti dzīvo augu atliekām ļoti bagātā vidē. Pašlaik ir zināms, ka jūras ežu zarnās dzīvo vairāk nekā 50 ciliātu sugas, kas sastopamas tikai piekrastes zonā, kur eži barojas ar aļģēm. Lielā dziļumā, kur aļģes vairs neaug, jūras ežiem nav ciliātu.

Saskaņā ar dzīvesveidu un uztura raksturu lielākā daļa jūras ežu zarnu skropstu ir zālēdāji. Viņi barojas ar aļģēm, kas lielos daudzumos piepilda saimnieka zarnas. Dažas sugas ir diezgan "izsmalcinātas" savā pārtikas izvēlē. Piemēram, strobilidium(Strobilidium, 118. att., A) barojas gandrīz tikai ar lieliem kramaļģiem. Ir arī plēsēji, kas ēd pārstāvjus citu, vairāk mazas sugas ciliāti.
Skropstiņos no jūras ežu zarnām, atšķirībā no astromātiem, nav stingras nosēšanās ar noteiktiem saimnieku veidiem. Viņi dzīvo visdažādākajos jūras ešos, kas ēd aļģes.
Jūras ežu inficēšanās veidi ar ciliātiem nav pētīti. Tomēr šeit ar lielu varbūtības pakāpi var pieņemt, ka tas notiek aktīvās brīvi peldošās formās. Fakts ir tāds, ka jūras ežu zarnu ciliāti jūras ūdenī var dzīvot ilgu laiku (daudzas stundas). Taču viņi jau tik ļoti pielāgojušies dzīvei ežu zarnās, ka ārpus ķermeņa, jūras ūdenī, agri vai vēlu iet bojā.
Pabeidzot iepazīšanos ar ciliātiem, vēlreiz jāuzsver, ka tie pārstāv sugām bagātu, plašu un pārtikušu dzīvnieku pasaules grupu (klasi). Paliekot šūnu organizācijas līmenī, infuzorijas, salīdzinot ar citām vienšūņu klasēm, ir sasniegušas vislielāko struktūras un funkciju sarežģītību.
Īpaši nozīmīga loma šajā progresīvajā attīstībā (evolūcijā) droši vien bija kodolaparāta transformācijai un kodolduālisma (kodolu kvalitatīvā neekvivalence) rašanās. Makrokodolu bagātība ar nukleīnvielām ir saistīta ar aktīviem vielmaiņas procesiem, ar enerģiskiem proteīnu sintēzes procesiem citoplazmā un kodolos.
Secinājums
Mēs esam nonākuši pie beigām mūsu apskatam par plaša veida dzīvnieku pasaules struktūru un dzīvesveidu - vienšūņi. Viņu raksturīgā iezīme, kā jau vairākkārt uzsvērts iepriekš, ir vienšūnas. Pēc savas struktūras vienšūņi ir šūnas. Taču tās nav salīdzināmas ar šūnām, kas veido daudzšūnu organismu ķermeni, jo tās pašas ir organismi. Tādējādi vienšūņi ir organismi šūnu organizācijas līmenī. Šķiet, ka daži augsti organizēti vienšūņi ar daudziem kodoliem jau pārsniedz šūnu struktūras morfoloģiskās robežas, kas dažiem zinātniekiem dod iemeslu šādus vienšūņus saukt par "supracelulāriem". Tas maz maina lietas būtību, jo vienšūnu organizācija joprojām ir raksturīga vienšūņiem.
Vienšūnu robežās vienšūņi ir nogājuši garu evolūcijas attīstības ceļu un ir devuši ļoti dažādas formas, kas pielāgotas visdažādākajiem dzīves apstākļiem. Vienšūņu ciltsraksta centrā ir divas klases: sarkodi un flagellāti. Jautājums par to, kura no šīm klasēm ir primitīvāka, joprojām tiek apspriests zinātnē. No vienas puses, Sarcodidae (ameba) apakšējiem pārstāvjiem ir visprimitīvākā struktūra. Bet flagellates uzrāda vislielāko vielmaiņas veida plastiskumu un it kā stāv uz robežas starp dzīvnieku un augu pasauli. Dažu sarkožu (piemēram, foraminifera) dzīves ciklā ir karogdziedzera stadijas (gametas), kas norāda uz to saistību ar flagellātiem. Ir acīmredzams, ka ne mūsdienu sarkodes, ne mūsdienu flagellates nevar būt sākotnējā dzīvnieku pasaules evolūcijas grupa, jo viņi paši ir gājuši garu ceļu vēsturiskajā attīstībā un ir izstrādājuši neskaitāmus pielāgojumus mūsdienu dzīves apstākļiem uz Zemes. Iespējams, abas šīs mūsdienu vienšūņu klases jāuzskata par diviem stumbriem evolūcijā, kas cēlušies no senām līdz mūsdienām neizdzīvotām formām, kas dzīvoja mūsu planētas dzīvības attīstības rītausmā.
Tālākajā vienšūņu evolūcijā notika dažāda rakstura izmaiņas. Dažas no tām izraisīja vispārēju organizācijas līmeņa paaugstināšanos, aktivitātes pieaugumu un dzīvības procesu intensitāti. Pie šādām filoģenētiskām (evolucionārām) pārvērtībām vajadzētu pieskaitīt, piemēram, kustību un barības uztveršanas organellu attīstību, kas ciliātu klasē sasniegusi augstu pilnības līmeni. Nav apstrīdams, ka skropstas ir organoīdi, kas atbilst (homologi) flagellām. Kamēr zīdaiņiem, ar dažiem izņēmumiem, ziepju skaits ir neliels, tad skropstiņu skaits sasniedzis daudzus tūkstošus. Ciliārā aparāta attīstība strauji palielināja vienšūņu aktivitāti, padarīja to attiecību formas ar vidi daudzveidīgākas un sarežģītākas, reakcijas formas uz ārējiem stimuliem. Diferencēta ciliāra aparāta klātbūtne neapšaubāmi bija viens no galvenajiem iemesliem progresīvai evolūcijai ciliātu klasē, kur radās ļoti dažādas formas, kas pielāgotas dažādiem biotopiem.
Šāda veida evolūcijas pārmaiņu piemērs ir skropstu ciliārā aparāta attīstība, ko nosauca Akad. Severtsova aromorfozes. Aromorfozēm raksturīgs vispārējs organizācijas pieaugums, plašas nozīmes adaptāciju attīstība. Ar organizācijas pieaugumu saprot izmaiņas, kas izraisa organisma vitālās aktivitātes pieaugumu; tie ir saistīti ar tā daļu funkcionālo diferenciāciju, kas noved pie daudzveidīgākām komunikācijas formām starp organismu un vidi. Ciliāru ciliārā aparāta attīstība attiecas tieši uz šāda veida strukturālām transformācijām evolūcijas procesā. Tā ir tipiska aromorfoze.
Vienšūņiem, kā uzsvēra V. A. Dogels, aromorfozes veida izmaiņas parasti ir saistītas ar organellu skaita palielināšanos. Notiek organellu polimerizācija. Ciliāru aparāta attīstība ciliātiem ir tipisks šādu izmaiņu piemērs. Otrs aromorfozes piemērs skropstu evolūcijā ir to kodolaparāts. Iepriekš mēs pārbaudījām skropstu kodola strukturālās iezīmes. Ciliātu kodolduālisms (mikrokodola un makrokodola klātbūtne) tika papildināts ar hromosomu skaita palielināšanos makrokodolā (poliploidijas fenomens). Tā kā hromosomas ir saistītas ar galvenajiem sintētiskajiem procesiem šūnā, galvenokārt ar proteīnu sintēzi, šis process izraisīja vispārēju galveno dzīvībai svarīgo funkciju intensitātes pieaugumu. Arī šeit notika polimerizācija, kas ietekmēja kodola hromosomu kompleksus.
ciliāti- viena no daudzskaitlākajām un progresīvākajām vienšūņu grupām, nāk no flagellātiem. Par to liecina to kustības organoīdu pilnīga morfoloģiskā līdzība. Šis evolūcijas posms bija saistīts ar divām lielām aromorfozēm: viena no tām skāra kustības organellus, otrā - kodolaparātu. Abi šie izmaiņu veidi ir savstarpēji saistīti, jo abas noved pie dzīvības aktivitātes palielināšanās un savstarpējo attiecību ar ārējo vidi formu sarežģījumiem.
Līdzās aromorfozēm pastāv vēl viens evolūcijas pārmaiņu veids, kas izpaužas adaptāciju (adaptācijas) attīstībā noteiktiem, asi definētiem eksistences apstākļiem. Šāda veida evolūcijas izmaiņas Severtsovs sauca par idioadaptāciju. Vienšūņu evolūcijā šāda veida izmaiņām bija ļoti liela nozīme. Iepriekš, apsverot dažādas klases vienšūņiem, ir sniegti daudzi idioadaptīvu izmaiņu piemēri. Pielāgošanās planktoniskajam dzīvesveidam dažādās vienšūņu grupās, pielāgošanās dzīvei smiltīs ciliātos, oocistu aizsargčaumalu veidošanās kokcīdijās un daudz kas cits - tās visas ir idioadaptācijas, kurām bija liela loma rašanās un attīstības procesā. atsevišķas grupas, bet nav saistītas ar vispārējām progresīvām izmaiņām organizācijā.
Adaptācijas dažādiem specifiskiem vienšūņu biotopiem ir ļoti dažādas. Tie nodrošināja šī tipa plašu izplatību visdažādākajos biotopos, kas tika detalizēti apspriests iepriekš, aprakstot atsevišķas klases.
Lielā medicīnas enciklopēdija
Infusoria Tetrahymena thermophila ... Wikipedia
- (Infusoria) visattīstītāko vienšūņu (Protozoa) klase. I. galvenās iezīmes: skropstu klātbūtne (kustībai un uzturam), divu veidu kodoli (poliploīds makrokodolis un diploīds mikrokodolis, kas atšķiras pēc struktūras un ... ... Lielā padomju enciklopēdija
Vai šķidrie dzīvnieki (Infusoria) vienšūņu klase, kas aprīkota ar mirdzošām bizēm vai skropstiņām, ko pieaugušā vecumā dažkārt aizstāj ar cilindriskiem izaugumiem, ar noteiktu ķermeņa formu, lielākoties ar ... ... enciklopēdiskā vārdnīca F. Brokhauss un I.A. Efrons
Ifuzoriju taksonomija klasifikācija un organismu sistematizēšana, kas pieder pie ciliātu tipa, saskaņā ar vispārējo vienšūņu taksonomijas koncepciju, jo īpaši, un dzīvniekiem kopumā Saturs 1 Sistemātika 1.1 Klasiskā ... Wikipedia
Vai šķidrie dzīvnieki (Infusoria) vienšūņu klase, kas aprīkota ar mirdzošām bizēm vai skropstiņām, kuras pieaugušā stāvoklī dažkārt aizstāj ar cilindriskiem sūcošiem izaugumiem, ar noteiktu ķermeņa formu, lielākoties ar ... ... Brokhausa un Efrona enciklopēdija
Ciliātu veidu parasti sauc arī par ciliātu - šo vienšūņu kustības orgāni ir skropstas. Ciliātu šūnai ir divi kodoli, tos sauc par maziem un lieliem. Pirmais regulē reprodukcijas procesu, bet otrais ir atbildīgs par uztura, kustību un elpošanas procesiem.
Šāda veida dzīves iezīmes jāņem vērā, piemēram, ciliātu apavu piemērā.
Kustības un elpošana
Infuzorija-kurpe, kuras garums ir aptuveni 0,5 mm, par savu dzīvotni izvēlas ūdenstilpes. Vienkāršākā ķermeņa formu ir viegli uzminēt pēc nosaukuma - tas atgādina apavu. Kustības ātrums ir aptuveni 2,5 mm sekundē.
Ārējā elastīgā apvalka klātbūtne nodrošina stabilu ķermeņa formu.
Citoplazmā, kas atrodas blakus čaumalam, ir atbalsta šķiedras, to attīstība ir ciliāta nemainīgās formas saglabāšanas garantija.
Uz skropstu virsmas ir 15 tūkstoši skropstu, to pamatnē atrodas bazālais ķermenis. Kustība notiek ar skropstu vibrāciju palīdzību: tās rada apmēram 30 sitienus sekundē, tādējādi stumjot skropstu apavu uz priekšu.
Tas elpo caur ķermeņa virsmu.
Uzturs
Ciliātu iezīme ir klātbūtne šūnu mute, kuras tuvumā ir īpaši garas un blīvas skropstas. Šūnu mute turpinās ar šūnu rīkli: skropstas iespiež tajā ūdeni un ciliāru barību - baktērijas.
Infuzorija jūt ķīmiskas vielas, kas atbrīvo baktēriju kopu. Tādējādi viņa meklē laupījumu.
Pēc tam pārtika nonāk gremošanas vakuolā, kur tā tiek sagremota. No šejienes tas jau nonāk citoplazmā.
Atlase
Atlase tiek veikta, izmantojot divas kontrakcijas vakuolas, viens atrodas priekšpusē, bet otrs atrodas aizmugurē. Vakuoli sastāv no rezervuāra un kanāliem.
Šķidrums piepilda kanālus, pēc tam seko centrālajam rezervuāram un pēc tam iziet no infuzorijas. Vakuolu kontrakcijas process ilgst 10-20 sekundes.
pavairošana
Infuzorija vairojas aseksuāli – sadalās divās daļās. Tās iezīme ir sadalījums visā ķermenī.
Skropstu kodoli ir sadalīti divās daļās: jaunizveidotajiem ciliātiem ir mazs un liels kodols. Meitu ciliātiem ir organoīdu daļas, un trūkstošie veidojas neatkarīgi. Reprodukcija notiek vairākas reizes dienā.
Ciliātiem-kurpēm iespējama arī dzimumvairošanās, taču šajā gadījumā īpatņu skaits nepalielinās. Uz laiku vienšūņi ir savienoti, veidojot savienojošo tiltu no citoplazmas.
Katrā indivīdā lielais kodols pazūd, un mazie tiek sadalīti divas reizes - parādās četri kodoli. No tiem paliek tikai viens kodols, kas arī sadalās. Indivīds satur divus kodolus, tad notiek kodolu apmaiņa – viens no kodoliem pāriet uz citu indivīdu.
Tur tas saplūst ar kodolu, kas paliek, un tādējādi katrā no indivīdiem veidojas mazs un lielāks kodols. Šis process, ko sauc konjugācija, ir nepieciešama ģenētiskā materiāla atjaunošanai starp indivīdiem.
Ciliātu veidi
Ciliāti ir sarežģīti organizēti vienšūņi, tiem ir aptuveni 7000 sugu.
1. Sarkodu daudzveidība.
2. Ziedu daudzveidība.
Otrā apakštipa pārstāvji pārvietojas ar flagellas palīdzību, kas atspoguļojas šīs grupas nosaukumā.
- Flagellates. Euglena green ir tipisks šīs grupas pārstāvis. Šim vienšūņiem ir fusiforma forma, pārklāta ar blīvu elastīgu membrānu. Ķermeņa priekšējā galā atrodas flagellum, netālu no tā pamatnes atrodas gaismas jutīga acs - stigma un kontrakta vakuola. Tuvāk aizmugurējam galam, citoplazmas biezumā, atrodas kodols, kas kontrolē visus organisma dzīvībai svarīgos procesus. Euglena zaļā krāsa ir saistīta ar daudziem hloroplastiem. Gaismā šis flagellāts veic fotosintēzi, bet, ilgstoši atrodoties neapgaismotās rezervuāra vietās, tas pāriet uz barību ar sarežģītu organisko vielu sabrukšanas produktiem, ekstrahējot tos no apkārtējās vides. Tādējādi zaļā euglena var apvienot gan autotrofisku metabolisma veidu, gan heterotrofisku. Organismu esamība ar šādu jauktu uztura veidu liecina par dzīvnieku un augu pasaules attiecībām. Lielākā daļa flagellas
3. Sporozoju un skropstu daudzveidība.
Ciliātu tipā ietilpst vienšūņi, kas izceļas ar vissarežģītāko organizāciju vienšūnu dzīvnieku vidū. Tipisks šīs grupas pārstāvis ir infuzorija-kurpe - parasta saldūdens iemītniece. Viņas ķermenis ir pārklāts ar blīvu apvalku - pīlingu, un tāpēc tam ir samērā nemainīga forma. Raksturīga struktūras iezīme ir skropstu klātbūtne, kas vienmērīgi pārklāj visu apavu korpusu. Skropstu kustības ir koordinētas kontraktilo šķiedru tīkla dēļ, kas atrodas citoplazmas virsmas slānī. Otra raksturīgā iezīme ir divu kodolu, lielo (makrokodolu) un mazo (mikrokodolu) klātbūtne. Kodoli atšķiras arī funkcionāli: lielais regulē vielmaiņu, bet mazais piedalās dzimumprocesā (konjugācijā). Infuzorijas apavi barojas ar baktērijām, vienšūnu aļģēm, kuras ar cilijām pielāgo šūnu mutei, kas atrodas pirmsmutes dobuma apakšā. Pārtikas daļiņas pēc izkļūšanas cauri šūnas rīklei, kas beidzas citoplazmā, tiek ievietotas gremošanas vakuolos, kur fermentu iedarbībā tās tiek sadalītas. Nesagremotās atliekas tiek izmestas caur pulveri. Apavu korpusā ir divas pārmaiņus pulsējošas sarežģītas kontrakcijas vakuolas. Kurpes vairojas aseksuāli, daloties uz pusēm. Daudzkārtēju aseksuālu vairošanos apavos aizstāj dzimumprocess – konjugācija, kuras laikā divi apavi saplūst kopā un apmainās ar ģenētisko materiālu. Pēc tam apavi atšķiras un drīz atkal sāk aseksuālu vairošanos. Konjugācijas bioloģiskā nozīme slēpjas divu indivīdu iedzimto īpašību apvienojumā vienā organismā. Tas palielina tā dzīvotspēju, kas izpaužas kā labāka pielāgošanās vides apstākļiem.
Vai neatradāt to, ko meklējāt? Izmantojiet meklēšanu
Šajā lapā materiāls par tēmām:
- īsumā par flagellātiem, pseidopodiem, sporozoāniem un skropstiņiem
- sarkodi un sporozoans
- flagellas dažādība
- infusoria slipper ziņojuma kopsavilkums
- kāda ir atšķirība starp ciliātu un sarkožu sugām
Viens no tipiskākajiem pazīstamākajiem ciliāru pārstāvjiem ir skropstu apavi. Tas, kā likums, dzīvo stāvošā virziena ūdenī, kā arī saldūdens tipa rezervuāros, kur straume izceļas ar pašpārliecinātības izņēmumu. Tās dzīvotnē obligāti jābūt trūdošām organiskām vielām. Būtu ieteicams detalizēti apsvērt visus šī faunas pārstāvja dzīves aspektus.
Skropstu pārstāvji

Jāpiebilst, ka ciliāti ir tips, kura nosaukums cēlies no vārda "tinktūra" (tulkojumā no latīņu valoda). Tas izskaidrojams ar to, ka pirmie vienšūņu pārstāvji tika atrasti tieši augu tinktūrās. Laika gaitā šāda veida attīstība sāka strauji uzņemt apgriezienus. Tādējādi jau šodien bioloģijā ir zināmi aptuveni 6-7 tūkstoši sugu, kas ietver arī ciliātu veidu. Ja paļaujamies uz 20. gadsimta 80. gadu datiem, tad var apgalvot, ka attiecīgais tips savā struktūrā satur divas klases: skropstiņveidīgie (ir trīs virskārtas) un Piesūcošie ciliāti. Saistībā ar šo informāciju varam secināt, ka dzīvo organismu daudzveidība ir ļoti plaša, kas rada patiesu interesi.
Infuzorijas veids: pārstāvji

Ievērojami šāda veida pārstāvji ir ciliates-balantidia un ciliates-shoe. Specifiskas īpatnībasŠie dzīvnieki ir apvalka pārklājums ar skropstiņām, kuras tiek izmantotas kustībai, skropstu aizsardzība caur speciāli izstrādātiem orgāniem, trihocistas (atrodas čaumalas ektoplazmā), kā arī divu kodolu (veģetatīvā) klātbūtne šūnā. un ģeneratīvā). Turklāt mutes dobums uz ciliāta ķermeņa veido mutes piltuvi, kurai ir tendence iekļūt šūnas mutē, kas ved uz rīkli. Tieši tur veidojas gremošanas vakuoli, kas kalpo tieši pārtikas gremošanai. Bet nesagremotās sastāvdaļas tiek izvadītas no ķermeņa caur pulveri. Ciliātu veida raksturojumsļoti daudzpusīgs, taču galvenie punkti ir apspriesti iepriekš. Vienīgais, kas jāpiebilst, ir tas, ka abi ciliāti atrodas pretējās ķermeņa daļās. Pateicoties to darbībai, no organisma tiek izvadīts liekais ūdens vai vielmaiņas produkti.
Infusoria apavi

Lai kvalitatīvi aplūkotu šādu interesantu vienšūnas uzbūves organismu uzbūvi un dzīvesveidu, būtu ieteicams atsaukties uz atbilstošo piemēru. Tam ir nepieciešami infuzorijas apavi, kas plaši izplatīti saldūdens rezervuāros. Tos var viegli audzēt parastos konteineros (piemēram, akvārijos), visvieglāk ir pļavas siena līcis saldūdens, jo šāda veida tinktūrās, kā likums, attīstās ļoti daudzas vienšūņu sugas, tai skaitā ciliāti-kurpes. Tātad, izmantojot mikroskopu, jūs varat praktiski izpētīt visu rakstā sniegto informāciju.
Ciliates-kurpju raksturojums
Kā minēts iepriekš, skropstas ir veids, kas ietver daudzus elementus, no kuriem interesantākais ir skropstu apavi. Tas ir pusmilimetra garums, kas apveltīts ar vārpstas formu. Jāpiebilst, ka vizuāli šis organisms atgādina kurpi, līdz ar to arī tik intriģējošs nosaukums. Infuzorijas kurpe pastāvīgi atrodas kustībā, un tā peld ar strupu galu uz priekšu. Interesanti, ka tā kustības ātrums nereti sasniedz 2,5 mm sekundē, kas šāda veida pārstāvim ir ļoti labi. Uz ciliātu-kurpju ķermeņa virsmas var novērot skropstas, kas kalpo kā motororganellas. Tāpat kā visiem ciliātiem, arī attiecīgā organisma struktūrā ir divi kodoli: lielais ir atbildīgs par uztura, elpošanas, motora un vielmaiņas procesiem, bet mazais piedalās seksuālajā aspektā.
Ciliates ķermenis-kurpes

Apavu ciliātu ķermeņa uzbūve ir ļoti sarežģīta. Šī pārstāvja ārējais pārklājums ir plāns elastīgs apvalks. Tas spēj saglabāt pareizu ķermeņa ķermeņa formu visu mūžu. Uzticīgi palīgi šajā jomā ir lieliski attīstītas atbalsta šķiedras, kas atrodas citoplazmas slānī, kas ir cieši blakus membrānai. Skropstu apavu korpusa virsma ir apveltīta ar milzīgu skaitu (apmēram 15 000) skropstu, kas svārstās neatkarīgi no ārējiem apstākļiem. Katra no tām pamatnē atrodas bazālais ķermenis. Cilijas pārvietojas apmēram 30 reizes sekundē, lai virzītu ķermeni uz priekšu. Svarīgi atzīmēt, ka šo instrumentu viļņveidīgās kustības ir ļoti koordinētas, kas ļauj infuzorijai kustības procesā lēni un skaisti griezties ap sava ķermeņa garenasi.
Ciliates - noteiktas intereses veids
Lai pilnībā izprastu visas ciliātu apavu iezīmes, ieteicams apsvērt galvenos tā dzīves procesus. Tātad runa ir par baktēriju un aļģu ēšanu. Organisma ķermenis ir apveltīts ar padziļinājumu, ko sauc par šūnu muti un kas nonāk rīklē, kura apakšā ēdiens nonāk tieši vakuolā. Tur tas tiek sagremots apmēram stundu, procesā veicot pāreju no skābas uz sārmainu vidi. Vakuoli pārvietojas ciliāta ķermenī caur citoplazmas plūsmu, un nesagremotās atliekas iziet ķermeņa aizmugurē caur pulveri.
Ciliātu apavu elpošana tiek veikta ar skābekļa palīdzību, kas caur ķermeņa apvalku nonāk citoplazmā. Un ekskrēcijas procesi notiek caur diviem kontrakcijas vakuoliem. Runājot par organismu uzbudināmību, ciliāti-kurpes mēdz apvienoties baktēriju kompleksos, reaģējot uz baktēriju izdalīto vielu iedarbību. Un tie peld prom no tāda kairinātāja kā galda sāls.
pavairošana

Apavu ciliāts var vairoties vienā no diviem veidiem. Plašāk ir kļuvusi bezdzimuma pavairošana, saskaņā ar kuru kodoli tiek sadalīti divās daļās. Šīs operācijas rezultātā katrs ciliāts satur 2 kodolus (lielus un mazus). Seksuālā pavairošana ir piemērota, ja ir kādi uztura trūkumi vai dzīvnieka ķermeņa temperatūras režīma izmaiņas. Jāatzīmē, ka pēc tam infuzorija var pārvērsties par cistu. Bet ar seksuālo reprodukcijas veidu indivīdu skaita pieaugums ir izslēgts. Tātad divi ciliāti uz noteiktu laiku tiek savienoti viens ar otru, kā rezultātā čaula izšķīst un starp dzīvniekiem veidojas savienojošais tilts. Svarīgi, lai katra lielais kodols pazūd bez pēdām, bet mazais sadalīšanās procesā izietu divas reizes. Tādējādi katrā ciliātā veidojas 4 meitas kodoli, pēc kuriem trīs no tiem tiek iznīcināti, bet ceturtais atkal tiek sadalīts. Šo seksuālo procesu sauc par konjugāciju. Un tā ilgums var būt līdz 12 stundām.
Infuzorija-kurpe dzīvo mazos stāvošos rezervuāros. Šim vienšūnas dzīvniekam, kura garums ir 0,5 mm, ir vārpstveida ķermenis, kas neskaidri atgādina kurpi. Ciliates pastāvīgi atrodas kustībā, peld ar strupu galu uz priekšu. Šī dzīvnieka kustības ātrums sasniedz 2,5 mm sekundē. Uz ķermeņa virsmas tiem ir kustības organelli - skropstas. Šūnā ir divi kodoli: liels kodols atbild par uzturu, elpošanu, kustībām, vielmaiņu; mazais kodols ir iesaistīts dzimumprocesā.
Apavu infuzorijas struktūra
Ciliātu organisms ir sarežģītāks. Plānais elastīgais apvalks, kas pārklāj ciliāta ārpusi, saglabā nemainīgu ķermeņa formu. To veicina arī labi attīstītās atbalsta fibrillas, kas atrodas citoplazmas slānī blakus apvalkam. Uz skropstas ķermeņa virsmas atrodas aptuveni 15 000 oscilējošu skropstu. Katras cilmes pamatnē atrodas bazālais ķermenis. Katras skropstas kustība sastāv no asa triepiena vienā virzienā un lēnākas, vienmērīgākas atgriešanās sākotnējā stāvoklī. Cilias vibrē apmēram 30 reizes sekundē un, tāpat kā airi, stumj infuzoriju uz priekšu. Skropstu viļņveidīgā kustība ir koordinēta. Kad ciliāta kurpe peld, tā lēnām griežas ap ķermeņa garenisko asi.
Dzīves procesi
Uzturs
Čības un daži citi brīvi dzīvojoši skropstiņi barojas ar baktērijām un aļģēm.
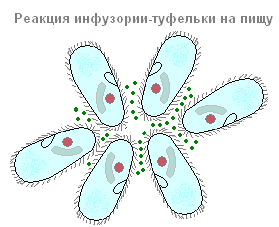
Ciliātu-kurpju reakcija uz pārtiku
Plāns elastīgs apvalks, ( šūnu membrānu) nosedzot skropstiņu no ārpuses, saglabā nemainīgu ķermeņa formu. Apmēram 15 tūkstoši skropstu atrodas uz ķermeņa virsmas. Uz ķermeņa ir padziļinājums - šūnu mute, kas nonāk šūnu rīklē. Rīkles apakšā pārtika nonāk gremošanas vakuolā. Gremošanas vakuolā pārtika tiek sagremota stundas laikā, vispirms ar skābu un pēc tam ar sārmainu reakciju. Gremošanas vakuoli pārvietojas skropstu ķermenī ar citoplazmas strāvu. Nesagremotās atliekas tiek izmestas ķermeņa aizmugurējā galā caur īpašu struktūru - pulveri, kas atrodas aiz mutes atveres.
Elpa
Elpošana notiek caur ķermeņa daļu. Skābeklis caur visu ķermeņa virsmu nonāk citoplazmā un oksidē sarežģītas organiskās vielas, kā rezultātā tās pārvēršas ūdenī, oglekļa dioksīdā un dažos citos savienojumos. Tajā pašā laikā tiek atbrīvota enerģija, kas nepieciešama dzīvnieka dzīvībai. Oglekļa dioksīds tiek izvadīts caur visu ķermeņa virsmu elpošanas laikā.
Atlase
Ciliātu-kurpju ķermenī ir divas kontrakcijas vakuolas, kas atrodas ķermeņa priekšējā un aizmugurējā galā. Tie savāc ūdeni ar izšķīdušām vielām, kas veidojas sarežģītu organisko vielu oksidēšanās laikā. Sasniedzot robežvērtību, saraušanās vakuoli tuvojas ķermeņa virsmai, un to saturs izplūst. Saldūdens vienšūnu dzīvniekiem liekā ūdens tiek izvadīts caur saraušanās vakuoliem, kas pastāvīgi nonāk viņu ķermenī no apkārtējās vides.
Aizkaitināmība
Reaģējot uz to izdalīto vielu iedarbību, ciliāti-kurpes pulcējas baktēriju kopās, bet aizpeld no tāda kairinātāja kā galda sāls.
Uzbudināmība ir visu dzīvo organismu īpašība reaģēt uz stimulu darbību – gaismu, siltumu, mitrumu, ķīmiskām vielām, mehāniskām ietekmēm. Aizkaitināmības dēļ vienšūnas dzīvnieki izvairās no nelabvēlīgiem apstākļiem, atrod barību, sava gada indivīdus.
pavairošana
aseksuāls
Ciliates parasti vairojas aseksuāli, daloties divās daļās. Kodoli ir sadalīti divās daļās, un katrā jaunajā ciliātā ir viens liels un viens mazs kodols. Katra no abām meitām saņem daļu organellu, bet pārējās veidojas no jauna.
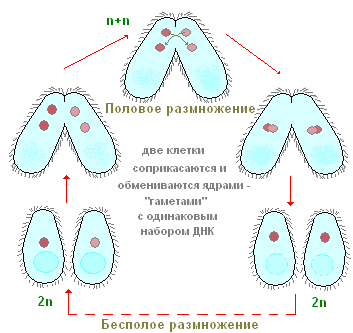
Ciliates-kurpju pavairošana
seksuāla
Ar pārtikas trūkumu vai temperatūras izmaiņām ciliāti pāriet uz seksuālo reprodukciju un pēc tam var pārvērsties par cistu.
Seksuālā procesa laikā indivīdu skaita pieaugums nenotiek. Divi ciliāti ir īslaicīgi saistīti viens ar otru. Saskares vietā čaula izšķīst, un starp dzīvniekiem veidojas savienojošais tilts. Katra ciliāta lielais kodols pazūd. Mazais kodols sadalās divas reizes. Katrā ciliātā veidojas četri meitas kodoli. Trīs no tiem tiek iznīcināti, bet ceturtais atkal tiek sadalīts. Rezultātā katrā paliek divi serdeņi. Kodolapmaiņa notiek gar citoplazmas tiltu, un tur tā saplūst ar atlikušo kodolu. Jaunizveidotie kodoli veido lielu un mazu kodolu, un ciliāti atšķiras. Šo seksuālo procesu sauc par konjugāciju. Tas ilgst apmēram 12 stundas. Seksuālais process izraisa atjaunošanos, apmaiņu starp indivīdiem un iedzimtā (ģenētiskā) materiāla pārdali, kas palielina organismu dzīvotspēju.
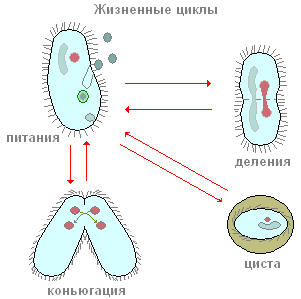
Skropstu apavu dzīves cikls
Mēs arī iesakām
 Komutācijas barošanas avots: remonts un uzlabošana
Komutācijas barošanas avots: remonts un uzlabošana
 Gaismas tālvadības pults
Gaismas tālvadības pults
 Peldēšanas nodarbības pirmsskolas vecuma bērniem
Peldēšanas nodarbības pirmsskolas vecuma bērniem
 Piezīmes meistaram - mājas sadzīves signalizācija
Piezīmes meistaram - mājas sadzīves signalizācija
 Pulksteņa propelleris uz Atmega8
Pulksteņa propelleris uz Atmega8
 Ierīču un releju pielietojuma piemēri, kā pareizi izvēlēties un pieslēgt releju Mikrokontrolleru un releju vienkāršas komutācijas shēmas
Ierīču un releju pielietojuma piemēri, kā pareizi izvēlēties un pieslēgt releju Mikrokontrolleru un releju vienkāršas komutācijas shēmas